
Investigation of the Sandfly Fauna of Central Arid Areas and Northern Humid Regions of Tunisia, with Morphological and Molecular Identification of the Recently Established Population of Phlebotomus (Larroussius) perfiliewi
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
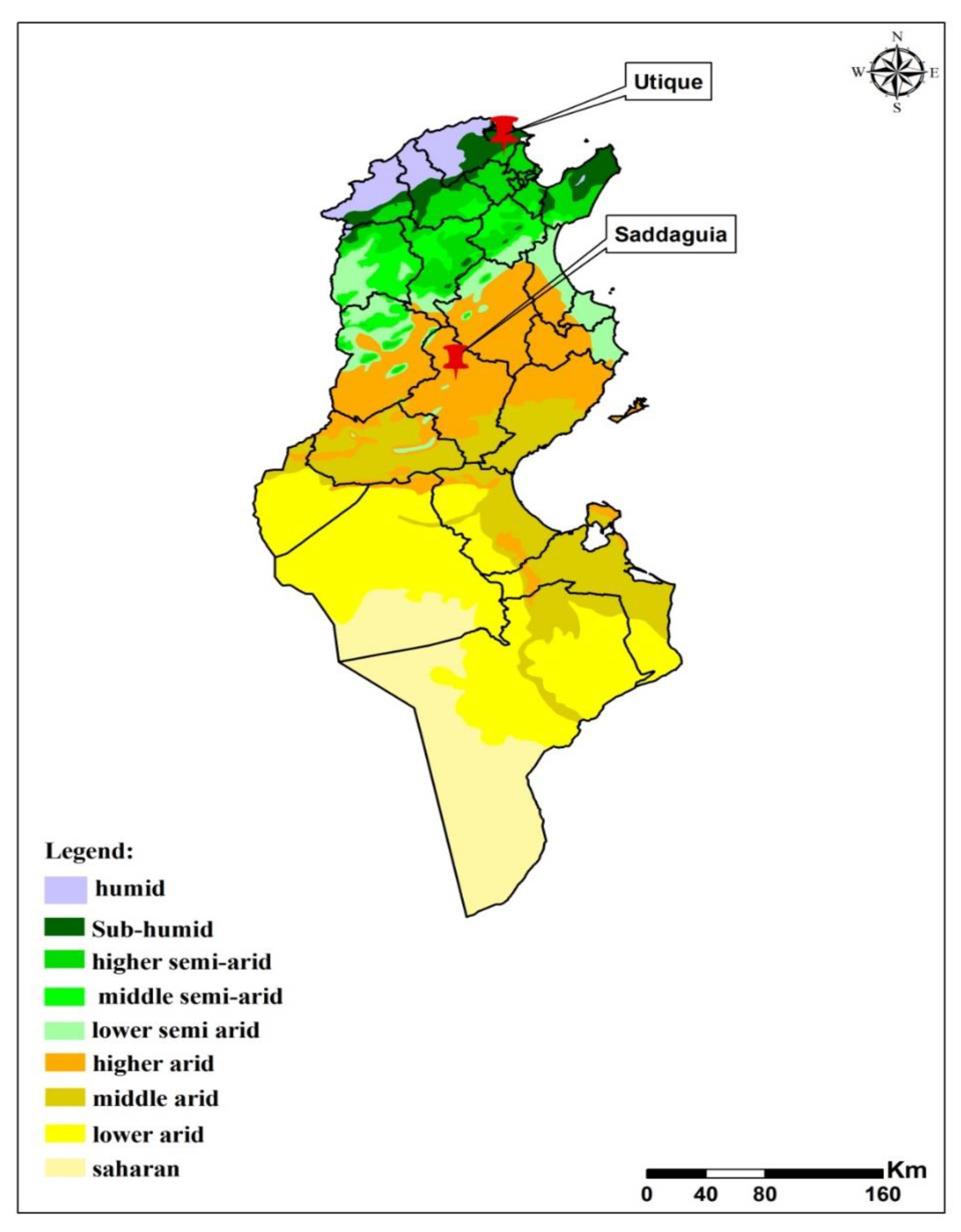
2.1. Study Sites
2.2. Collection of Sandflies
2.3. Population Structure and Diversity Indices
2.4. Molecular Analysis
3. Results
3.1. Sandfly Species, Relative Abundance, and Population Structure
3.2. Morphological Analysis
3.3. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izri, M.; Belazzoug, S. Phlebotomus (Larroussius) perfiliewi naturally infected with dermotropic Leishmania infantum at Tenes, Algeria. Trans. Roy. Soc. Trop. Med. Hyg. 1993, 87, 399. [Google Scholar] [CrossRef]
- Bennai, K.; Tahir, D.; Lafri, I.; Bendjaballah-Laliam, A.; Bitam, I.; Parola, P. Molecular detection of Leishmania infantum DNA and host blood meal identification in Phlebotomus in a hypoendemic focus of human lesihmaniasis in northern Algeria. PLoS Negl. Trop. Dis. 2018, 12, e0006513. [Google Scholar] [CrossRef]
- Maroli, M.; Gramiccia, M.; Gradoni, L. Natural infections of sandfy Phlebotomus perfiliewi with Leishmania infantum in a cutaneous leishmaniasis focus of the Abruzzi region, Italy. Trans. Roy. Soc. Trop. Med. Hyg. 1987, 81, 596–598. [Google Scholar] [CrossRef]
- Barhoumi, W.; Fares, W.; Cherni, S.; Derbali, M.; Dachraoui, K.; Chelbi, I.; Ramalho-Ortigao, M.; Beier, J.C.; Zhioua, E. Changes of sand fly populations and Leishmania infantum infection rates in an irrigated village located in arid Central Tunisia. Inter. J. Envir. Res. Public Health 2016, 13, 329. [Google Scholar] [CrossRef] [Green Version]
- Remadi, L.; Chargui, N.; Jiménez, M.; Molina, R.; Haouas, N.; González, E.; Chaabane-Banaouas, R.; Ben Salah, E.; Haddaji, M.; Chaabouni, Y.; et al. Molecular detection and identification of Leishmania DNA and blood meal analysis in Phlebotomus (Larroussius) species. PLoS Negl. Trop. Dis. 2020, 14, e0008077. [Google Scholar] [CrossRef]
- Rioux, J.A.; Croset, H.; Leger, N.; Rosin, G. Présence au Maroc de Phlebotomus perfiliewi. Ann. Parasitol. Hum. Comp. 1977, 52, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Parrot, L. Notes sur les phlébotomes. XXV. Présence de Phlebotomus perfiliewi dans la banlieue d’Alger. Arch. Inst. Pasteur Alger. 1941, 19, 360–361. [Google Scholar]
- Russo, J.; Rioux, J.A.; Lambert, P.; Rispal, A.; Belmonte, S.; Bechri, S. Chorologie des phlébotomes de l’Est Algérien (Diptera, Psychodidae). Ann. Parasitol. Hum. Comp. 1991, 66, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Parrot, L.; Clastrier, J. Notes sur les phlébotomes. XLV. Présence de Phlebotomus perfiliewi en Tunisie. Arch. Inst. Pasteur Alger. 1944, 22, 60–62. [Google Scholar]
- Dancesco, P.; Dedet, J.P.; Ben Osman, F.; Chadli, A. Les phlébotomes capturés dans des foyers de leishmaniose canine à Tunis. Rôle probable de Phlebotomus perniciosus et Phlebotomus perfiliewi dans la transmission. Arch. Inst. Pasteur Tunis. 1970, 52, 65–88. [Google Scholar]
- Izri, M.A.; Marty, P.; Fauran, P.; Le Fichoux, Y.; Rousset, J.J. Phlebotomus perfiliewi Parrot, 1930 (Dipteran, Psychodidae) dans le sud-est de la France. Parasite 1994, 1, 286. [Google Scholar] [CrossRef] [Green Version]
- Maroli, M.; Bigliocchi, F.; Khoury, C. Sandflies in Italy: Observation on their distribution and methods for control. Parasitologia 1994, 36, 251–264. [Google Scholar]
- Lewis, D. A taxonomic review of the genus Phlebotomus. Bull. Brit. Mus. Nat. His. (Ent.) 1982, 45, 121–209. [Google Scholar]
- Rioux, J.A.; Lanotte, G.; Petter, F.; Deureure, J.; Akalay, O.; Pratlong, F.; Velez, I.D.; Fikri, N.B.; Maazoun, R.; Denial, M.; et al. Les leishmanioses cutanées du bassin Méditerranéen occidental, de l’identification enzymatique à l’analyse éco-épidémiologique. L’exemple de trois «foyers» tunisien, marocain et français. In Leishmania. Taxonomie et Phylogénèse. Applications éco-épidémiologiques. International Colloquium CNRS/INSERM, (Juillet, 1984); IMEEE: Montpellier, France, 1986; pp. 471–478. [Google Scholar]
- Rioux, J.A.; Rispail, P.H.; Lanotte, G.; Lepart, J. Relations phlébotomes-bioclimats en écologie des leishmanioses. Corollaires épidémiologiques. L’exemple du Maroc. Bull. Soc. Bot. Franç. 1984, 131, 549–557. [Google Scholar] [CrossRef]
- Fares, W.; Charrel, R.N.; Dachraoui, K.; Bichaud, L.; Barhoumi, W.; Derbali, M.; Cherni, S.; Chelbi, I.; de Lamballerie, X.; Zhioua, E. Infection of sand flies collected from different bio-geographical areas with phleboviruses. Acta Trop. 2015, 141, 1–6. [Google Scholar] [CrossRef]
- Ayadi, A.; Ben Ismail, R.; Ben Rachid, M.S. Extension de l’aire de transmission du Kala Azar à Leishmania infantum (Nicolle 1908) vers le centre et le sud de la Tunisie. Arch. Inst. Pasteur Tunis. 1991, 68, 269–273. [Google Scholar]
- Besbes, A.; Pousse, H.; Ben Said, M.; Kharrat, H.; Chenimi, L. Leishmanioses viscérales infantiles du centre Tunisien (221 cas). Méd. Mal. Infect. 1994, 24, 628–634. [Google Scholar] [CrossRef]
- Ben Salah, A.; Ben Ismail, R.; Amri, F.; Chlif, S.; Ben Rzig, F.; Karrat, H.; Hadhri, H.; Hassouna, M.; Dellagi, K. Investigation of the spread of human visceral leishmaniasis in central Tunisia. Trans. Roy. Soc. Trop. Med. Hyg. 2000, 94, 382–386. [Google Scholar] [CrossRef]
- Zhioua, E.; Kaabi, B.; Chelbi, I. Entomological investigation following the spread of visceral leishmaniasis in Tunisia. J. Vect. Ecol. 2007, 32, 371–374. [Google Scholar] [CrossRef]
- Zoghlami, Z.; Chouihi, E.; Barhoumi, W.; Dachraoui, K.; Massoudi, N.; Ben Helel, K.; Habboul, Z.; Hadhri, M.H.; Limam, S.; Mhadhbi, M.; et al. Interaction between canine and human visceral leishmaniases in a holoendemic focus of Central Tunisia. Acta Trop. 2014, 139, 32–38. [Google Scholar] [CrossRef]
- Depaquit, J.; Bounamous, A.; Akhoundi, M.; Augot, D.; Sauvage, F.; Dvorak, V.; Chaibullinova, A.; Pesson, B.; Volf, P.; Léger, N. A taxonomy study of Phlebotomus (Larroussius) perfiliewi s. l. Infect. Gen. Evol. 2013, 20, 500–508. [Google Scholar] [CrossRef]
- Gounot, M.; Le Houerou, H.N. Essai de synthèse sur la végétation et la phyto-écologie tunisiennes. In Élément de Botanique et de Phyto-Écologie Tunisiennes; Tome, I., Nabli, M.A., Eds.; Faculty of Sciences of Tunis and MAB: Tunis, Tunisia, 1989; p. 387. [Google Scholar]
- Croset, H.; Rioux, J.A.; Juminer, B.; Tour, S. Fluctuations annuelles des populations de Phlebotomus perniciosus Newstead, 1911, Phlebotomus perfiliewi Parrot, 1930, et Sergentomyia munita parroti (Adler et Theodor, 1927) (Diptera-Psychodidae) en Tunisie du Nord. Arch. Inst. Pasteur Tunis 1970, 47, 43–56. [Google Scholar]
- Chelbi, I.; Derbali, M.; AL-Ahmadi, Z.; Zaafouri, B.; El Fahem, A.; Zhioua, E. Phenology of Phlebotomus papatasi (Diptera: Psychodidae) relative to the seasonal prevalence of zoonotic cutaneous leishmaniasis in Central Tunisia. J. Med. Entomol. 2007, 44, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Dachraoui, K.; Chelbi, I.; Ben Said, M.; Ben Osman, R.; Cherni, S.; Charrel, R.; Zhioua, E. Transmission dynamic of Punique virus in Tunisia. Viruses 2022, 14, 904. [Google Scholar] [CrossRef]
- Abonnenc, E. Les phlébotomes de la région éthiopienne (Diptera, Psychodidae). Mém. ORSTOM 1972, 55, 289. [Google Scholar]
- Croset, H.; Rioux, J.A.; Master, M.; Bayar, N. Les phlébotomes de la Tunisie (Diptera, Phlebotominae). Mise au point systématique, chorologique et éthologique. Ann. Parasitol. Hum. Comp. 1978, 53, 711–749. [Google Scholar] [CrossRef] [Green Version]
- Léger, N.; Pesson, B.; Madulo-Leblond, G.; Abonnenc, E. Sur la différentiation des femelles du sous-genre Larroussius Nitzulescu, 1931 (Diptera: Phlebotominae) de la région méditerranéenne. Ann. Parasitol. Hum. Comp. 1983, 58, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Pesson, B.; Ready, J.S.; Benabdennbi, I.; Martín-Sánchez, J.; Esseghir, S.; Cadi-Soussi, M.; Morillas-Marquez, F.; Ready, P.D. Sandflies of the Phlebotomus perniciosus complex: Mitochondrial introgression and a new sibling species of Phlebotomus longicuspis in the Moroccan Rif. Med. Vet. Entomol. 2004, 18, 25–37. [Google Scholar] [CrossRef]
- Boussaa, S.; Boumezzough, A.; Remy, P.E.; Glasser, N.; Pesson, B. Morphological and isoenzymatic differentitation of Phlebotomus perniciosus and Phlebotomus longicuspis (Diptera: Psychodidae) in Southern Morocco. Acta Trop. 2008, 106, 184–189. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species diversity and the « Shannon-Wiener » Index. Glob. Ecol. Biogeogr. 2003, 12, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Depaquit, J.; Ferté, H.; Léger, N.; Killick-Kendrick, R.; Rioux, J.A.; Killick-Kendrick, M.; Hanafi, H.A.; Gobert, S. Molecular systematics of the phlebotomine sandflies of the subgenus Paraphlebotomus (Diptera-Psychodidae, Phlebotomus) based on ITS2 rDNA sequences. Hypotheses of dispersion and speciation. Insect. Mol. Biol. 2000, 9, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Depaquit, J.; Léger, N.; Lefranc, F.; Alves-Pirez, C.; Hanafi, H.; Maroli, M.; Morillas-Marquez, F.; Rioux, J.A.; Svobodova, M.; Volf, P. ITS 2 sequences heterogeneity in Phlebotomus sergenti and Phlebotomus similis (Diptera, Psychodidae): Possible consequences in their ability to transmit Leishmania tropica. Inter. J. Parasitol. 2002, 32, 1123–1131. [Google Scholar] [CrossRef]
- Esseghir, S.; Ready, P.D.; Killick-Kendrick, R.; Ben-Ismail, R. Mitochondrial haplotypes and phylogeography of Phlebotomus vectors of Leishmania major. Insect. Mol. Biol. 1997, 6, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Depaquit, J. Molecular systematics applied to Phlebotomine sandflies: Review and perspectives. Infect. Genet. Evol. 2014, 28, 744–756. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chelbi, I.; Kaabi, B.; Bejaoui, M.; Derbali, M.; Zhioua, E. Spatial correlation between Phlebotomus papatasi Scopoli (Diptera: Psychodidae) and incidence of zoonotic cutaneous leishmaniasis in Tunisia. J. Med. Entomol. 2009, 46, 400–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghrab, J.; Rhim, A.; Bhach-Hamba, D.; Chahed, M.K.; Aoun, K.; Nouira, S.; Bouratbine, A. Phlebotominae (Diptera: Psychodidae) of human leishmaniasis sites in Tunisia. Parasite. 2006, 13, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Barhoumi, W.; Chelbi, I.; Zhioua, E. Effects of the development of irrigation systems in the arid areas on the establishment of Phlebotomus (Larroussius) perfiliewi Parrot, 1930. Bull. Soc. Pathol. Exot. 2012, 105, 403–405. [Google Scholar] [CrossRef]
- Barhoumi, W.; Qualls, W.A.; Archer, R.S.; Fuller, D.O.; Chelbi, I.; Cherni, S.; Derbali, M.; Arheart, K.L.; Zhioua, E.; Beier, J.C. Irrigation in the arid regions of Tunisia impacts the abundance and apparent density of sandfly vectors of Leishmania infantum. Acta Trop. 2014, 141, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, W.; Dachraoui, K.; Barhoumi, W.; Cherni, S.; Chelbi, I.; Zhioua, E. Co-circulation of Toscan virus and Leishmania infantum in a focus of zoonotic visceral leishmaniasis from Central Tunisia. Acta Trop. 2020, 204, 105342. [Google Scholar] [CrossRef]
- Abbas, M.A.S.; Lachheb, J.; Chelbi, I.; Louati, D.; Dachraoui, K.; Ben Miled, S.; Zhioua, E. Independant circulation of Leishmania major and Leishmania tropica in their respective sandfly vectors for transmission of zoonotic and chronic cutaneous leishmaniasis co-existing in a mixed focus of Central Tunisia. Pathogens 2022, 11, 855. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ph. perf. 113 | Ph. pern. 92 | Ph. long. 20 | Ph. pap. 8 | |
|---|---|---|---|---|
| pi | 0.48497854 | 0;39484979 | 0.08583691 | 0.03433476 |
| Pi2 | 0.23520419 | 0.15590635 | 0.00736798 | 0.00117888 |
| ∑pi2 | 0.39965739 | |||
| Ar | 48.5 | 39.5 | 8.5 | 3.5 |
| Is | 2.50214315 | |||
| H | 1.51190652 | |||
| Equitability | 0.75336671 |
| Ph. perf. 295 | Ph. pern. 238 | Ph. long. 65 | Ph. pap. 87 | |
|---|---|---|---|---|
| pi | 0.43065693 | 0.34744526 | 0.09489051 | 0.1270073 |
| pi2 | 0.1854654 | 0.12071821 | 0.00900421 | 0.01613085 |
| ∑pi2 | 0.33131866 | |||
| Ar | 43.0656934 | 34.7445255 | 9.48905109 | 12.7007299 |
| Is | 3.01824228 | |||
| H | 2.46274993 | |||
| Equitability | 1.22716173 |
| Code of Specimens (GenBank Accession Number) | Nearest Sequence | Query Cover | Homology | Identified Species |
|---|---|---|---|---|
| 4F_uti (ON515731) | MN385555 | 100% | 99.05% | Ph. perfiliewi |
| 6M_uti (ON515732) | MN385555 | 100% | 97.79% | Ph. perfiliewi |
| 7M_uti (ON515733) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 8M_uti (ON515734) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 21F_sad (ON515735) | MN385544 | 100% | 100% | Ph. perfiliewi |
| 22F_sad (ON515736) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 23F_sad (ON515737) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 24F_sad (ON515738) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 26M_sad (ON515739) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 27M_sad (ON515740) | MN385555 | 100% | 99.68% | Ph. perfiliewi |
| 28M_sad (ON515741) | MN385555 | 100% | 100% | Ph. perfiliewi |
| 30F_sad (ON515742) | MN385555 | 100% | 100% | Ph. perfiliewi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chelbi, I.; Abdi, A.; Depaquit, J.; Fares, W.; Abbas, M.A.S.; Dachraoui, K.; Zhioua, E. Investigation of the Sandfly Fauna of Central Arid Areas and Northern Humid Regions of Tunisia, with Morphological and Molecular Identification of the Recently Established Population of Phlebotomus (Larroussius) perfiliewi. Insects 2022, 13, 1057. https://doi.org/10.3390/insects13111057
Chelbi I, Abdi A, Depaquit J, Fares W, Abbas MAS, Dachraoui K, Zhioua E. Investigation of the Sandfly Fauna of Central Arid Areas and Northern Humid Regions of Tunisia, with Morphological and Molecular Identification of the Recently Established Population of Phlebotomus (Larroussius) perfiliewi. Insects. 2022; 13(11):1057. https://doi.org/10.3390/insects13111057
Chicago/Turabian StyleChelbi, Ifhem, Amani Abdi, Jérôme Depaquit, Wasfi Fares, Mohammed Abdo Saghir Abbas, Khalil Dachraoui, and Elyes Zhioua. 2022. "Investigation of the Sandfly Fauna of Central Arid Areas and Northern Humid Regions of Tunisia, with Morphological and Molecular Identification of the Recently Established Population of Phlebotomus (Larroussius) perfiliewi" Insects 13, no. 11: 1057. https://doi.org/10.3390/insects13111057