
European and Mediterranean Myzocallidini Aphid Species: DNA Barcoding and Remarks on Ecology with Taxonomic Modifications in An Integrated Framework
 , ,
, , {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aphids Collection and Specimen Identification
2.2. DNA Extraction, Amplification and Sequencing
2.3. Nucleotide Distance Analyses and Taxa Molecular Delimitation
3. Results
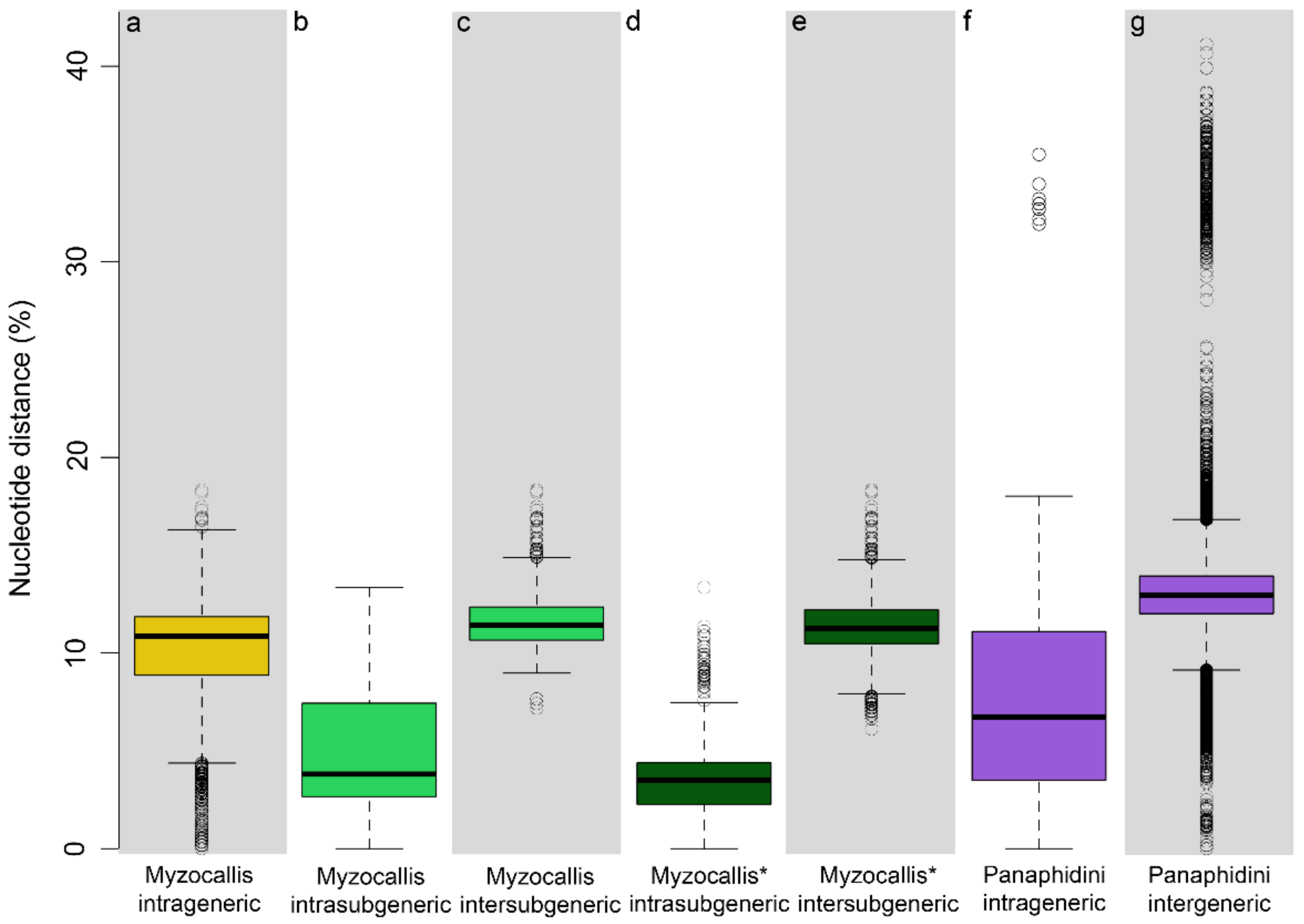
3.1. Nucleotide Distance Analyses
3.2. Species Delimitation of the Genus Myzocallis
4. Discussion
4.1. Myzocallis Subgenus Myzocallis
4.2. Myzocallis Subgenus Agrioaphis
4.3. Myzocallis Subgenus Pasekia
4.4. Nearctic Myzocallis Subgenera Lineomyzocallis and Neomyzocallis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Trees: An Identification and Information Guide; CAB International: Wallingford, UK, 1994; p. 987. Available online: www.aphidsonworldsplants.info (accessed on 23 September 2022).
- Favret, C. Aphid Speciesfile. Version 5.0/5.0. 2022. Available online: https://Aphid.SpeciesFile.org (accessed on 23 September 2022).
- Quednau, F.W.; Remaudière, G. Le genre Myzocallis Passerini, 1860: Classification mondiale des sous-genres et nouvelle espèces paléarctiques (Homoptera: Aphididae). Can. Entomol. 1994, 126, 303–326. [Google Scholar] [CrossRef]
- Quednau, W. Atlas of the Drepanosiphine aphids of the world. Part I: Panaphidini—Myzocallidina. Contrib. Am. Entomol. Inst. 1999, 31, 1–281. [Google Scholar]
- Denk, T.; Grimm, G.W.; Manos, P.S.; Deng, M.; Hipp, A.L. An updates infrageneric classification of the oaks: Review of previous taxonomic schemes and synthesis of evolutionary patterns. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus, L.; Gil-Pelegrín, E., Peguero-Pina, J., Sancho-Knapik, D., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. II. The family Drepanosiphidae; Scandinavian Science Press: Klampenborg, Denamrk, 1982; p. 176. [Google Scholar]
- Stroyan, H.L.G. Homoptera Aphidoidea, Chaitophoridae & Callaphidinae. In Handbook Identification British Insects vol. II; Royal Entomological Society: London, UK, 1977; p. 130. [Google Scholar]
- Nieto Nafría, J.M.; Mier Durante, M.P. Hemiptera, Aphididae I., Fauna Iberica; CSIC: Madrid, Spain, 1998. [Google Scholar]
- Osiadacz, B.; Wieczorek, K.M. Myzocallis (Lineomyzocallis) walshii Monell, 1879 (Hemiptera, Aphidoidea), an aphid species new for Poland. Pol. J. Entom. 2006, 75, 233–338. [Google Scholar]
- Lee, Y.; Lee, W.; Kanturski, M.; Foottit, R.G.; Akimoto, S.I.; Lee, S. Cryptic diversity of the subfamily Calaphidinae (Hemiptera: Aphididae) revealed by comprehensive DNA barcoding. PLoS ONE 2016, 12, e0176582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbagallo, S.; Massimino Cocuzza, G.E. Description of a new Myzocallis (Hemiptera Aphididae) living on Valonia oak in Southern Italy with DNA barcoding accounts on allied species-group. Zootaxa 2022, 5183, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Remaudière, G. Découverte en France de l’espèce américaine Myzocallis (Lineomyzocallis) walshii (Monell) (Hom. Aphididae). Ann. Société Entomol. De Fr. 1989, 25, 117. [Google Scholar]
- Patti, I.; Lozzia, G.C. Presenza in Italia dell’afide neartico della quercia rossa, Myzocallis (Lineomyzocallis) walshii (Mon.). Boll. Zool. Agric. E Bachic. 1994, 26, 141–145. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identification through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Schindel, D.E.; Miller, S.E. DNA barcoding a useful tool for taxonomists. Nature 2005, 435, 17. [Google Scholar] [CrossRef] [Green Version]
- Virgilio, M.; Backeljan, I.; Nevado, B.; De Meyer, M. Comparative performances of DNA barcoding across insect orders. BMC Bioinform. 2010, 11, 206. [Google Scholar] [CrossRef] [Green Version]
- Jimbo, U.; Kato, T.; Ito, M. Current progress in DNA barcoding and future implications for entomology. Entomol. Sci. 2011, 14, 107–124. [Google Scholar] [CrossRef]
- Magoga, G.; Sassi, D.; Daccordi, M.; Leonardi, C.; Mirzaei, M.; Regalin, R.; Lozzia, G.; Montagna, M. Barcoding Chrysomelidae: A resource for taxonomy and biodiversity conservation in the Mediterranean Region. Zookeys 2016, 597, 27–38. [Google Scholar] [CrossRef]
- Magoga, G.; Sahin, D.C.; Fontaneto, D.; Montagna, M. Barcoding of Chrysomelidae of Euro-Mediterranean area: Efficiency and problematic species. Sci. Rep. 2018, 8, 13398. [Google Scholar] [CrossRef]
- Massimino Cocuzza, G.E.; Cavalieri, V. Identification of aphids of Aphis frangulae-group living in Lamiaceae species through DNA barcode. Mol. Ecol. Resour. 2014, 14, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Kanturski, M.; Lee, Y.; Choi, J.; Lee, S. DNA barcoding and a precise morphological comparison revealed a cryptic species in the Nippolachnus piri complex (Hemiptera: Aphididae: Lachninae). Sci. Rep. 2018, 8, 8998. [Google Scholar] [CrossRef] [PubMed]
- Xi-Cha, Z.; Jing, C.; Rui, C.; Li-Yun, J.; Ge-Xia, Q. DNA barcoding and species delimitation of Chaitophorinae (Hemiptera, Aphididae). Zootaxa 2017, 656, 25–50. [Google Scholar]
- Foottit, R.G.; Maw, H.E.L.; Pike, K.S. DNA barcodes to explore diversity in aphids (Hemiptera Aphididae and Adelgidae). Redia 2009, 92, 87–91. [Google Scholar]
- Kim, H.; Lee, S.; Jang, Y. Macroevolutionary Patterns in the Aphidini Aphids (Hemiptera: Aphididae): Diversification, Host Association, and Biogeographic Origins. PLoS ONE 2011, 6, e24749. [Google Scholar] [CrossRef]
- Massimino Cocuzza, G.E.; Di Silvestro, S.; Giordano, R.; Rapisarda, C. Congruence between cytochrome oxidase I (COI) and morphological data in Anuraphis spp. (Hemiptera, Aphididae) with a comparison between the utility of the 5′ barcode and 3′ COI regions. Zookeys 2015, 529, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Kinyanjui, G.; Khamis, F.M.; Mohamed, S.; Ombura, L.O.; Warigia, M.; Ekesi, S. Identification of aphid (Hemiptera: Aphididae) species of economic importance in Kenya using DNA barcodes and PCR-RFLP based approach. Bull. Entomol. Res. 2016, 106, 63–72. [Google Scholar] [CrossRef]
- Rebijith, K.B.; Asokan, R.; Krishna Kumar, N.K.; Krishna, V.; Chaitanya, B.N.; Ramamurthy, V.V. DNA barcoding and elucidation of cryptic aphid species (Hemiptera: Aphididae) in India. Bull. Entomol. Res. 2013, 103, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Montagna, M.; Sassi, D.; Giorgi, A. Pachybrachis holerorum (Coleoptera: Chrysomelidae: Cryptocephalinae), a new species from the Apennines, Italy, identified by integration of morphological and molecular data. Zootaxa 2013, 3741, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Magoga, G.; Fontaneto, D.; Montagna, M. Factors affecting the efficiency of molecular species delimitation in a species-rich insect family. Mol. Ecol. Res. 2021, 21, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europea; Cambridge University Press: Cambridge, UK, 2001; Volume V, p. 2392. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagrcole-New Business Media: Bologna, Italy, 2017; Volume I, p. 1064. [Google Scholar]
- Quednau, F.W. Atlas of the drepanosiphine aphids of the world. Part II: Panaphidini Oestlund, 1923—Panaphidina Oestlund, 1923 (Hemiptera: Aphididae: Callaphidinae). Mem. Am. Entomol. Inst. 2003, 72, 301. [Google Scholar]
- Quednau, F.W. Atlas of the Drepanosiphine aphids of the world. Part III: Mindarinae Tullgren, 1909; Neophyllaphidinae Takahashi, 1921; Lizeriinae, E.E. Blanchard, 1923; Pterastheniinae Remaudière & Quednau, 1988; Macropodaphidinae Zachvatkin & Aizenberg, 1960; Taiwanaphidinae Quednau & Remaudière, 1994; Spicaphidinae Essig, 1953; Phyllaphidinae Herrich-Schaeffer in Koch, 1957; Israelaphidinae Ilharco, 1961; Saltusaphidinae Baker, 1920 (Hemiptera: Sternorrhycha, Aphididae). Mem. Am. Entomol. Inst. 2010, 83, 361. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Griekspoor, A.; Groothuis, T. 4Peaks V 1.7. 2019. Available online: https://nucleobytes.com (accessed on 20 September 2022).
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Taura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Popescu, A.A.; Huber, K.T.; Paradis, E. Ape 3.0: New tools for distance based phylogenetics and evolutionary analysis in R. Bioinformatics 2012, 28, 1536–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.D.J.; Collins, R.A.; Boyer, S.; Lefort, M.-C.; Malumbres-Olarte, J.; Vink, C.J.; Cruickshank, R.H. SPIDER: An R package for the analysis of species identity and evolution, with particular reference to DNA barcoding. Mol. Ecol. Res. 2012, 12, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Lanfaer, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1974, 6, 461–464. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate likelihood ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Coeur D’acier, A.; Cruaud, A.; Artige, E.; Genson, G.; Clamens, A.L.; Pierce, E.; Hudaverdian, S.; Simon, J.-C.; Jousselin, E.; Rasplus, J.Y. DNA barcoding and the associated PhylAphidB@se Web site for the identification of European aphids (Insecta: Hemiptera: Aphididae). PLoS ONE 2014, 9, e97620. [Google Scholar] [CrossRef] [PubMed]
- Piffaretti, J.; Vanlenberghe-Masutti, F.; Tayeh, A.; Clamens, A.L.; Coeur D’acier, A.; Jousselin, E. Molecular phylogeny reveals the existence of two sibling species in the aphid pest Brachycaudus helichrysi (Hemiptera: Aphididae). Zool. Scr. 2012, 41, 266–280. [Google Scholar] [CrossRef]
- Popkin, M.; Piffaretti, J.; Clamens, A.L.; Qiao, G.X.; Chen, J.; Vitalis, K.; Vanlerberghe-Masutti, F.; Gupta, R.K.; Mamaari, M.; Langella, O.; et al. Large-scale phylogeographic study on the cosmopolitan aphid pest Brachycaudus helichrysi reveals host plant associated lineages that evolved in allopatry. Biol. J. Linn. Soc. 2017, 120, 102–114. [Google Scholar]
- Richards, W.R. A synopsis of the world fauna of Myzocallis. Mem. Entom. Soc. Can. 1968, 57, 1–76. [Google Scholar]
- Hille Ris Lambers, D.; Stroyan, H.L.G. Two new species and a new subspecies of aphid from Italy. Mem. Soc. Entomol. Ital. 1959, 38, 84–94. [Google Scholar]
- Hille Ris Lambers, D. On Palestine aphids, with description of new subgenera and new species (Homoptera, Aphididae). Trans. R. Entom. Soc. Lond. 1948, 99, 269–289. [Google Scholar] [CrossRef]
- Remaudière, G.; Nieto Nafría, J.M. Un Myzocallis nouveau du Sud-Ouest de l’Europe (Hom. Aphididae). Ann. Soc. Entom. Fr. 1974, 10, 751–756. [Google Scholar]
- Tsitsipis, J.A.; Katis, N.I.; Margaritopoulos, J.T.; Lykourresis, D.P.; Avgelis, A.D.; Gargalianou, I.; Zarpas, K.D.; Perdikis, D.C.; Papapanayotoua, A. A contribution to the aphid fauna of Greece. Bull. Insectology 2007, 60, 31–38. [Google Scholar]
- Vučetic, A.; Jovičić, I.; Petrović-Obradović, O. Several new and one nvasive aphid species (Aphididae, Hemiptera) caught by yellow water trap in Serbia. Phytoparasitica 2014, 42, 247–257. [Google Scholar] [CrossRef]
- Holman, J. Host Plant Catalogue of Aphids: Palaearctic Region; Springer: Berlin/Heidelberg, Germany, 2009; p. 1216. [Google Scholar]
- Patti, I.; Barbagallo, S. Annotazioni integrative sugli afidi della Sicilia. Redia 1999, 82, 49–82. [Google Scholar]
- Barbagallo, S.; Stroyan, H.L.G. Osservazioni biologiche, ecologiche e tassinomiche dell’afidofauna della Sicilia. Frustula Entomol. 1982, 3, 1–182. [Google Scholar]
- Barbagallo, S.; Binazzi, A. Gli Afidi delle Querce in Italia. In Atti Convegno “Problematiche fitopatologiche del genere Quercus in Italia”; Stamperia Granducale: Firenze, Italia, 1990; pp. 142–160. [Google Scholar]
- Barbagallo, S.; Stroyan, H.L.G. Rilievi preliminari sulla composizione dell’afidofauna siciliana. In Proceedings of the Atti XI Congresso Nazionale Italiano di Entomologia, Portici-Sorrento, Italy, 10–15 May 1976; pp. 241–251. [Google Scholar]
- Quednau, F.W.; Barbagallo, S. A new genus and three new species of aphids from Quercus calliprinos Webb in the mediterranean countries (Homoptera: Aphididae). Can. Ent. 1991, 123, 581–593. [Google Scholar] [CrossRef]
- Giardina, G.; Raimondo, F.M.; Spadaro, V. A catalogue of plants growing in Sicily. Bocconea 2007, 20, 5–582. [Google Scholar]
- La Greca, M. L’uso delle categorie sistematiche sottogenere e sottospecie in tassonomia, alla luce della ricerca biogeografica. Boll. Dell'istituto Di Entomol. Dell'universita Degli Studi Di Bologna 1987, 41, 159–171. [Google Scholar]
- Gebiola, M.; Bernardo, U.; Ribes, A.; Gibson, G.A.P. An integrative study of Necremnus Thomson (Hymenoptera: Eulophidae) associated with invasive pests in Europe and North America: Taxonomic and ecological implications. Zool J. Linn. Soc. 2015, 173, 352–423. [Google Scholar] [CrossRef] [Green Version]
- Montagna, M.; Kubisz, D.; Mazur, M.A.; Magoga, G.; Ścibior, R.; Kajtoch, Ł. Exploring species-level taxonomy in the Cryptocephalus flavipes species complex (Coleoptera: Chrysomelidae). Zool. J. Linn. Soc. 2017, 179, 92–109. [Google Scholar] [CrossRef] [Green Version]
- Montagna, M.; Gonella, E.; Mereghetti, V.; Magoga, G.; Ferrari, E.; Pontini, M.; Ferracini, C.; Alma, A. Molecular species delimitation of the Asian chestnut gall wasp biocontrol agent released in Italy. Insect Syst. Evol. 2019, 50, 327–345. [Google Scholar] [CrossRef]
- Masonick, P.; Weirauch, C. Integrative species delimitation in Nearctic ambush bugs (Heteroptera: Reduviidae: Phymatinae): Insights from molecules, geometric morphometrics and ecological associations. Syst. Entomol. 2020, 45, 205–223. [Google Scholar] [CrossRef]
- Gadawski, P.; Montagna, M.; Rossaro, B.; Gilka, W.; Pesic, V.; Grabowski, M.; Magoga, G. DNA barcoding of Chironomidae from the Lake Skadar region: Reference library and a comparative analysis of the European fauna. Divers. Distrib. 2022, 28, 1–20. [Google Scholar] [CrossRef]
- Salvi, D.; Berrilli, E.; D’Alessandro, P.; Biondi, M. Sharpening the DNA barcoding tool through a posteriori taxonomic validation: The case of Longitarsus flea beetles (Coleoptera: Chrysomelidae). PLoS ONE 2020, 15, e0233573. [Google Scholar] [CrossRef]
- Marconi, M.; Modesti, A.; Alvarez, L.P.; Ogoña, P.V.; Mendoza, A.C.; Vecco-Giove, C.D.; Luna, J.O.; Di Giulio, A.; Mancini, E. DNA Barcoding of Stingless Bees (Hymenoptera: Meliponini) in Northern Peruvian Forests: A Plea for Integrative Taxonomy. Diversity 2022, 14, 632. [Google Scholar] [CrossRef]
- Mayr, E. Principles of Systematic Zoology; Mc Grew-Hill: New York, NY, USA, 1969; p. 428. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massimino Cocuzza, G.E.; Magoga, G.; Montagna, M.; Nieto Nafría, J.M.; Barbagallo, S. European and Mediterranean Myzocallidini Aphid Species: DNA Barcoding and Remarks on Ecology with Taxonomic Modifications in An Integrated Framework. Insects 2022, 13, 1006. https://doi.org/10.3390/insects13111006
Massimino Cocuzza GE, Magoga G, Montagna M, Nieto Nafría JM, Barbagallo S. European and Mediterranean Myzocallidini Aphid Species: DNA Barcoding and Remarks on Ecology with Taxonomic Modifications in An Integrated Framework. Insects. 2022; 13(11):1006. https://doi.org/10.3390/insects13111006
Chicago/Turabian StyleMassimino Cocuzza, Giuseppe Eros, Giulia Magoga, Matteo Montagna, Juan Manuel Nieto Nafría, and Sebastiano Barbagallo. 2022. "European and Mediterranean Myzocallidini Aphid Species: DNA Barcoding and Remarks on Ecology with Taxonomic Modifications in An Integrated Framework" Insects 13, no. 11: 1006. https://doi.org/10.3390/insects13111006