
Detection of Chitin Synthase Mutations in Lufenuron-Resistant Spodoptera frugiperda in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Populations
2.2. Chemicals
2.3. Bioassay
2.4. Bioinformatics Analysis
2.5. Sequencing Partial SfCHSA
2.6. Gene Expression of SfCHS in Different Tissues
2.7. Effects of Lufenuron on the Expression of SfCHS Gene
2.8. Statistical Analyses
3. Results
3.1. Bioassays
3.2. Homology Comparison and Molecular Phylogenetic Analysis of SfCHSA and SfCHSB Genes
3.3. Detection of Mutations of CHSA in the S. frugiperda Population
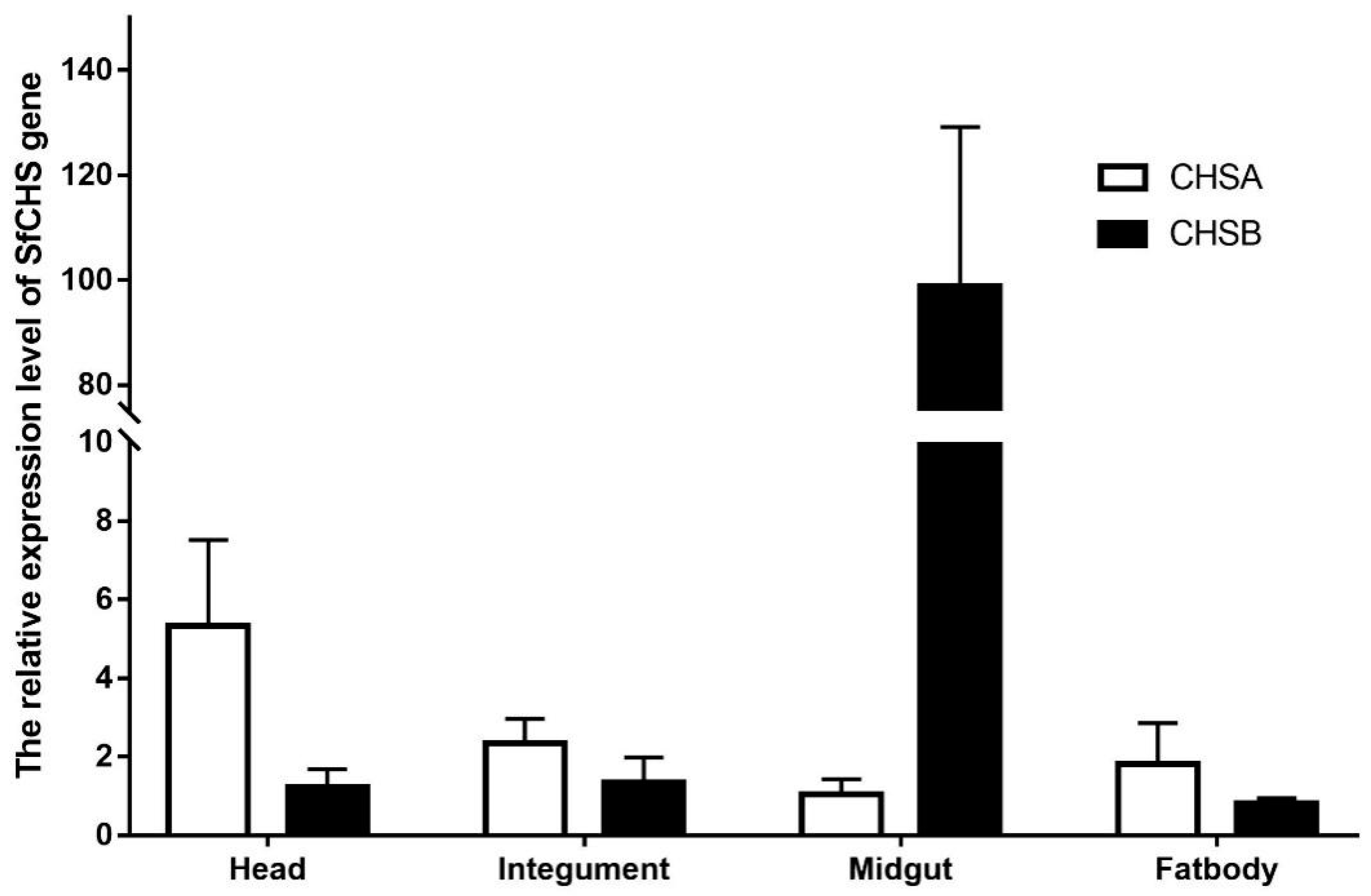
3.4. Expression of SfCHS Gene in Different Tissues
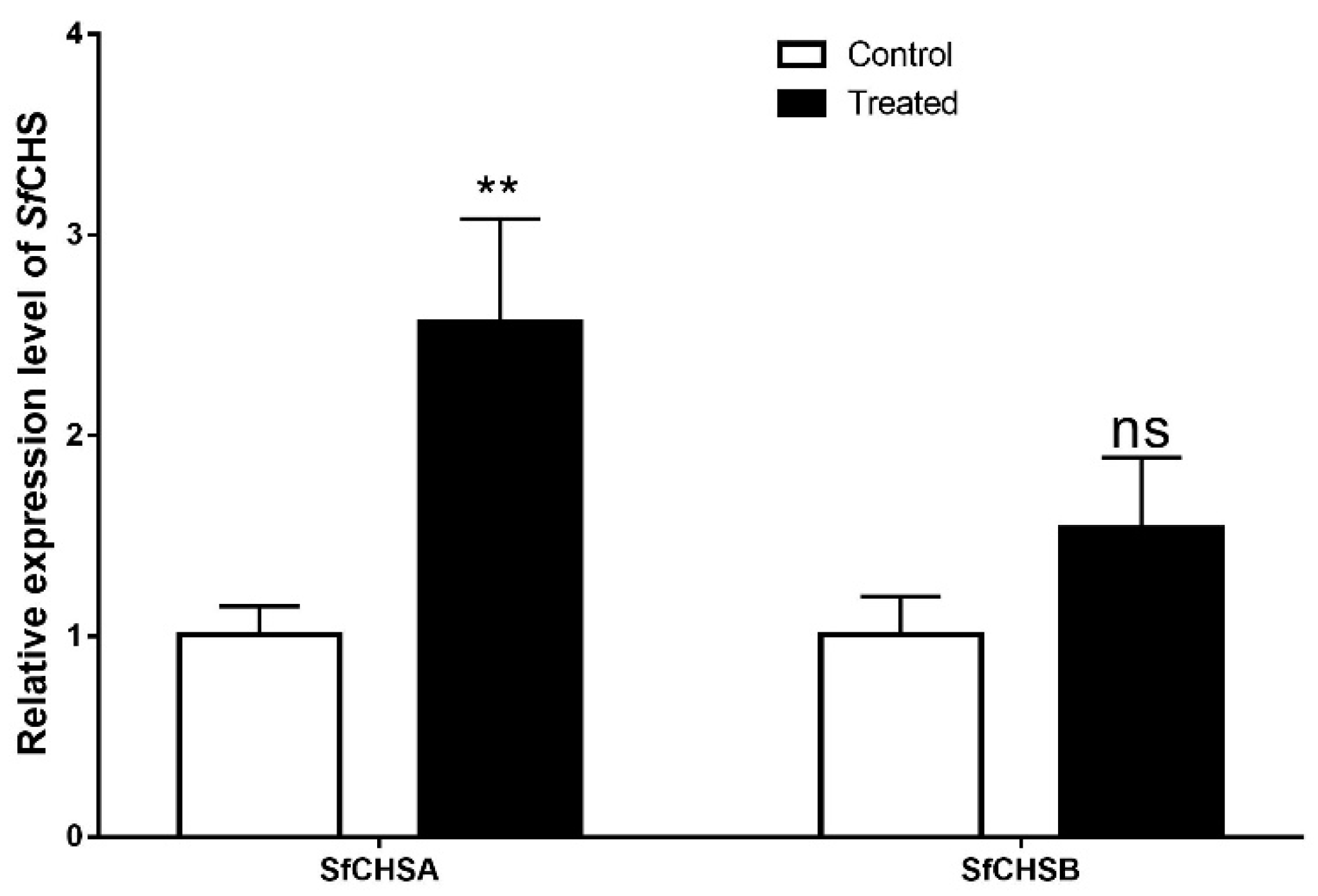
3.5. Effect of Lufenuron on the Expression of Chitin Synthase Genes (SfCHSA and SfCHSB) in S. frugiperda
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rose, A.H.; Silversides, R.H.; Lindquist, O.H. Migration flight by an aphid, Rhopalosiphum maidis (Hemiptera: Aphididae) and a noctuid, Spodoptera frugiperda (Lep. Noctuidae ). Can. Entomol. 1975, 107, 567–576. [Google Scholar] [CrossRef]
- Todd, E.L.; Poole, R.W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the western hemisphere. Ann. Entomol. Soc. Am. 1980, 73, 722–738. [Google Scholar] [CrossRef]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Liu, J.; Zhu, X.M. Occurrence dynamics and future trend of Spodoptera frugiperda invading China. China Plant Prot. 2019, 39, 33–35. [Google Scholar]
- Deshmukh, S.; Kalleshwaraswamy, C.M.; Asokan, R. First report of the fall armyworm, Spodoptera frugiperda (J.E, Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Yang, X.L.; Liu, Y.C.; Luo, M.Z.; Li, Y.; Wang, W.H.; Wan, F.; Jiang, H. The first discovery of Spodoptera frugiperda in Jiangcheng county, Yunnan Province. Yunnan Agric. 2019, 1, 72. [Google Scholar]
- Chen, H.; Yang, X.L.; Zhan, A.D.; Li, Y.C.; Wang, D.H.; Liu, J.; Hu, G. Immigration timing and origin of the first fall armyworms (Spodoptera frugiperda) detected in China. Chin. J. Appl. Entomol. 2020, 57, 1270–1278. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Integr. Agric. 2019, 18, 2–10. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M. Fall armyworm impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st. ed.; CIMMYT/USAID: EI Batan, Mexico, 2018; pp. 2–10. Available online: https://www.researchgate.net/publication/323612839 (accessed on 1 July 2022).
- Wu, K.M. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar]
- Gu, R.C.; Tang, Y.L.; Wu, Y.Y.; Niu, X.H.; Li, Q.Y.; Guo, Z.B.; Xiang, L.; Jiang, R.X.; Lei, Y.F.; Liu, X.; et al. Comparison of gut bacteria between Spodoptera frugiperda and Mythimna seperata feeding on sorghum in Chongqing area. J. Southwest Univ. Nat. Sci. Ed. 2019, 41, 6–13. [Google Scholar]
- Liu, J.; Jiang, Y.Y.; Li, H.; Li, Y.H.; Tai, H.K.; Wang, Z.Y. A preliminary report on sugarcane damage by Spodoptera frugiperda. China Plant Prot. 2019, 39, 35–36+66. [Google Scholar]
- Xu, L.N.; Hu, B.J.; Su, W.H.; Qi, R.D.; Qiu, K.; Zheng, Z.Y.; Zhang, Q.Y.; Zhou, Q.Y.; Zhou, Z.Y.; Qi, S.S.; et al. Fall armyworm damaging early sowing wheat in Anhui Province. Plant Prot. 2019, 45, 87–89. [Google Scholar]
- Zhao, M.; Men, X.Y.; Li, L.L.; Yang, J.G.; Wang, Z.Y.; Zhu, J.S.; Zhu, Y.Y.; Jiang, Y.Y.; Xu, Z.C.; Zhu, P.; et al. Spodoptera frugiperda were found damaging potato in Shandong Province. Plant Prot. 2019, 45, 84–86+97. [Google Scholar]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [Green Version]
- Merzendorfer, H. The cellular basis of chitin synthesis in fungi and insects: Common principles and differences. Eur. J. Cell Biol. 2011, 90, 759–769. [Google Scholar] [CrossRef]
- Tetreau, G.; Cao, X.L.; Chen, Y.R.; Muthukrishnan, S.; Jiang, H.B.; Blissard, G.W.; Kanost, M.R.; Wang, P. Overview of chitin metabolism enzymes in Manduca sexta: Identification, domain organization, phylogenetic analysis and gene expression. Insect Biochem. Mol. Biol. 2015, 62, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fan, H.W.; Huang, H.J.; Xue, J.; Wu, W.J.; Bao, Y.Y.; Xu, H.J.; Zhu, Z.R.; Cheng, J.A.; Zhang, C.X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-Sucking insects, Nilaparvata lugens and Laodelphax striatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. Biochem. Syst. Environ. Physiol. 2006, 176, 1–15. [Google Scholar] [CrossRef]
- Zimoch, L.; Merzendorfer, H. Immunolocalizaion of chitin synthase in the tobacco hornworm. Cell Tissue Res. 2002, 308, 287–297. [Google Scholar] [CrossRef]
- Arakane, Y.; Hogenkamp, D.G.; Zhu, Y.C.; Kramer, K.J.; Specht, C.A.; Beeman, R.W.; Kanost, M.R.; Muthukrishnan, S. Characterization of two chitin synthase genes of the red flour beetle, Tribolium castaneum, and alternate exon usage in one of the genes during development. Insect Biochem. Mol. Biol. 2004, 34, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Hogenkamp, D.G.; Arakane, Y.; Zimoch, L.; Merzendorfer, H.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Specht, C.A.; Muthukrishnan, S. Chitin synthase genes in Manduca sexta: Characterization of a gut-Specific transcript and differential tissue expression of alternately spliced mRNAs during development. Insect Biochem. Mol. Biol. 2005, 35, 529–540. [Google Scholar] [CrossRef]
- Boaventura, D.; Bolzan, A.; Padovez, F.E.; Okuma, D.M.; Omotoc, C.; Nauen, R. Detection of a ryanodine receptor target-Site mutation in diamide insecticide resistant fall armyworm, Spodoptera frugiperda. Pest Manag. Sci. 2020, 76, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Douris, V.; Steinbach, D.; Panteleri, R.; Livadarasa, I.; Pickette, J.A.; Leeuwen, T.V.; Nauen, R.; Vontasa, J. Resistance mutation conserved between insects and mites unravels the benzoylurea insecticide mode of action on chitin biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 14692–14697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishaaya, I.; Casida, J.E. Dietary TH 6040 alters composition and enzyme activity of housefly larval cuticle. Pest. Biochem. Physiol. 1974, 4, 484–490. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, K.Y. Characterization of a chitin synthase cDNA and its increased mRNA level associated with decreased chitin synthesis in Anopheles quadrimaculatus exposed to diflubenzuron. Insect Biochem. Mol. Biol. 2006, 36, 712–725. [Google Scholar] [CrossRef]
- Wu, J.W. On the application of insect growth regulators in control of agricultural insect pests. Pesticides 2002, 41, 6–8. [Google Scholar]
- Rui, C.H.; Liu, J.; Ren, L. Toxicological mechanisms and resistance of insect growth regulators. J. Biosaf. 2012, 21, 177–183. [Google Scholar]
- Zhou, Z.S.; Deng, G.R.; Luo, S.P. Research and application of insect growth regulator. Guangxi Agric. Sci. 2003, 1, 34–36. [Google Scholar]
- Luo, H.; Yang, Y.Q.; Chen, J. Progress on investigation of synthetic chitin inhibitors. Chem. Reag. 2009, 31, 697–700+712. [Google Scholar]
- Sheng, D.R.; Li, J.J.; Ha, S.L.D.; He, C.; Zhang, R. Effects of benzoylphenylurea insecticides on growth, development and reproduction of Scatopse sp. Henan Agric. Sci. 2020, 49, 85–90. [Google Scholar]
- Sheng, D.R.; He, C.; Lu, Q.; Chen, X.X.; Tian, X.J.; Zhang, H.R. Bioactivity and sublethal effect of benzoylurea insecticide against to Bradysia difformis. North. Hortic. 2018, 18, 42–47. [Google Scholar]
- Sun, L.L.; Li, L.; Jiang, C.X.; Zhu, Y. Advances in synthetic methods of lufenuron. Zhejiang Chem. Ind. 2011, 42, 5–7. [Google Scholar]
- Guo, Z.G.; An, J.J.; Gao, Z.L.; Dang, Z.H.; Pan, W.L.; Yuan, W.L.; Li, Y.F. Determination of the biological activity of benzoylurea insecticides against Apolygus lucorum. Xinjiang Agric. Sci. 2019, 56, 61–66. [Google Scholar]
- Lv, S.L.; Shi, Y.; Zhang, J.C.; Liang, P.; Zhang, L.; Gao, X.W. Detection of ryanodine receptor target-site mutations in diamide insecticide-resistant Spodoptera frugiperda in China. Insect Sci. 2021, 28, 639–648. [Google Scholar] [CrossRef]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to chlorantraniliprole and cross-Resistance to other diamide insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, M.; Hilliou, F.; Fricaux, T.; Audant, P.; Feyereisen, R.; Goff, G.L. Cytochrome P450s from the fall armyworm (Spodoptera frugiperda): Responses to plant allelochemicals and pesticides. Insect Mol. Blol. 2015, 24, 115–128. [Google Scholar] [CrossRef]
- Ma, L.; Dai, W.; Zhang, C.N. Toxicity of hexaflumuron and its effects on chitin and chitinase of cotton bollworm, helicoverpa armigera. J. Northwest AF Univ. Nat. Sci. Ed. 2014, 42, 141–147. [Google Scholar]
- Leeuwen, T.V.; Demaeght, P.; Osborne, E.J.; Dermauw, W.; Gohlke, S.; Nauen, R.; Grbic, M.; Tirry, L.; Merzendorfer, H.; Clark, R.M. Population bulk segregant mapping uncovers resistance mutations and the mode of action of a chitin synthesis inhibitor in arthropods. Proc. Natl. Acad. Sci. USA 2012, 109, 4407–4412. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, A.R.B.; Farias, J.R.; Bernardi, D.; Horikoshi, R.J.; Omoto, C. Genetic basis of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to the chitin synthesis inhibitor lufenuron. Pest Manag. Sci. 2016, 72, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Shiotsuki, T.; Jouraku, A.; Miura, K.; Minakuchi, C. Benzoylurea resistance in western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae): The presence of a point mutation in chitin synthase 1. J. Pestic. Sci. 2017, 42, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.G.; Cryan, J.R. Lufenuro, a chitin-Synthesis inhibitor, interrupts development of Drosophila melanogaster. J. Exp. Zool. 1997, 278, 37–44. [Google Scholar] [CrossRef]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Mol. Biol. 2005, 14, 453–463. [Google Scholar] [CrossRef]
- Zimoch, L.; Hogenkamp, D.G.; Kramer, K.J.; Muthukrishnan, S.; Merzendorfer, H. Regulation of chitin synthesis in the larval midgut of Manduca sexta. Insect Biochem. Mol. Biol. 2005, 35, 515–527. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Liu, X.J.; Zhang, J.Q.; Sun, Y.; Guo, Y.P.; Ma, E.B.; Zhu, K.Y. Silencing of two alternative splicing-derived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locust, Locusta migratoria manilensis (Meyen). Insect Biochem. Mol. Biol. 2010, 40, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Zhao, G.Y.; Yang, H.; Zhang, Y.N.; Fan, D. Effect of ecdysone and validamycin on chitin synthase B gene expression in Mythimna separata. J. Appl. Insect 2019, 5, 1026–1036. [Google Scholar]
- Bolognesi, R.; Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Terra, W.R.; Ferreira, C. Sequences of cDNAs and expression of genes encoding chitin synthase and chitinase in the midgut of Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2005, 35, 1249–1259. [Google Scholar] [CrossRef]
- Kumar, S.; Dahiya, B.; Chauhan, R. Ovicidal action of diflubenzuron against Helicoverpa armigera (Lepidoptera: Noctuidae). Agric. Sci. Dig. 1994, 14, 1–4. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence | |
|---|---|---|
| I1040M-F | 5′-ATCTCCTTCGGCTATATTCTTGAT-3′ | PCR |
| I1040M-R | 5′-CTTCGTCTTCTTAACTGCCACTTC-3′ | |
| L18-F | 5′-CGTATCAACCGACCTCCACT-3′ | RT-qPCR |
| L18-R | 5′-AGGCACCTTGTAGAGCCTCA-3′ | |
| CHSA-F | 5′-TCCTTATGTTGGTGGGTGCC-3 | |
| CHSA-R | 5′-GTACCGACGATCACAGCCAT-3′ | |
| CHSB-F | 5′-ATCCAGTTCACCGCCATGTT-3′ | |
| CHSB-R | 5′-CCAAGTCGTCGGTGTTCAGA-3′ |
| Insecticide | Strain | Coordinate | Year | Slope ± SE | LD50 (95% CL) (μg a.i. cm−2) | Df (χ2) | RR |
|---|---|---|---|---|---|---|---|
| Lufenuron | SUS | 114.8° E 30.85° N | 2019 | 1.226 ± 0.171 | 0.010 (0.007–0.014) | 18 (14.48) | 1.0 |
| Hainan | 110.28° E 20.02° N | 2019 | 1.376 ± 0.229 | 0.012 (0.007–0.016) | 14 (12.90) | 1.2 | |
| 2020 | 2.216 ± 0.311 | 0.036 (0.030–0.045) | 18 (5.922) | 3.6 | |||
| Guangdong | 113.22° E 23.4° N | 2019 | 2.028 ± 0.298 | 0.023 (0.017–0.029) | 14 (7.528) | 2.3 | |
| 2020 | 1.017 ± 0.170 | 0.022 (0.017–0.030) | 21 (2.965) | 2.2 | |||
| Guangxi | 103.14° E 24.40° N | 2019 | 2.327 ± 0.321 | 0.021 (0.017–0.027) | 14 (5.599) | 2.1 | |
| 2020 | 2.579 ± 0.309 | 0.020 (0.017–0.024) | 18 (16.78) | 2.0 | |||
| Hubei | 114.8° E 30.85° N | 2019 | 1.648 ± 0.218 | 0.023 (0.017–0.031) | 18 (11.29) | 2.3 | |
| 2020 | 1.400 ± 0.229 | 0.024 (0.016–0.032) | 18 (6.196) | 2.4 | |||
| Hunan | 111.73° E 27.25° N | 2019 | 1.803 ± 0.266 | 0.031 (0.022–0.039) | 14 (10.25) | 3.1 | |
| 2020 | 1.599 ± 0.240 | 0.021 (0.014–0.027) | 18 (8.065) | 2.1 | |||
| Jiangxi | 115.8° E 29.02° N | 2019 | 2.263 ± 0.299 | 0.024 (0.019–0.029) | 18 (9.056) | 2.4 | |
| 2020 | 1.047 ± 0.173 | 0.034 (0.023–0.050) | 22 (8.807) | 3.4 | |||
| Sichuan | 105.06° E 31.1° N | 2019 | 2.883 ± 0.348 | 0.018 (0.015–0.022) | 18 (14.06) | 1.8 | |
| 2020 | 1.092 ± 0.216 | 0.040 (0.026–0.058) | 18 (12.47) | 4.0 | |||
| Yunnan | 103.15° E 24.76° N | 2019 | 2.983 ± 0.417 | 0.028 (0.022–0.033) | 14 (8.952) | 2.8 | |
| 2020 | 1.347 ± 0.223 | 0.027 (0.018–0.037) | 18 (9.651) | 2.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, S.-L.; Xu, Z.-Y.; Li, M.-J.; Mbuji, A.L.; Gu, M.; Zhang, L.; Gao, X.-W. Detection of Chitin Synthase Mutations in Lufenuron-Resistant Spodoptera frugiperda in China. Insects 2022, 13, 963. https://doi.org/10.3390/insects13100963
Lv S-L, Xu Z-Y, Li M-J, Mbuji AL, Gu M, Zhang L, Gao X-W. Detection of Chitin Synthase Mutations in Lufenuron-Resistant Spodoptera frugiperda in China. Insects. 2022; 13(10):963. https://doi.org/10.3390/insects13100963
Chicago/Turabian StyleLv, Sheng-Lan, Zheng-Yi Xu, Ming-Jian Li, Amosi Leonard Mbuji, Meng Gu, Lei Zhang, and Xi-Wu Gao. 2022. "Detection of Chitin Synthase Mutations in Lufenuron-Resistant Spodoptera frugiperda in China" Insects 13, no. 10: 963. https://doi.org/10.3390/insects13100963