
Antimicrobial Activity of Apidermin 2 from the Honeybee Apis mellifera
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honeybees
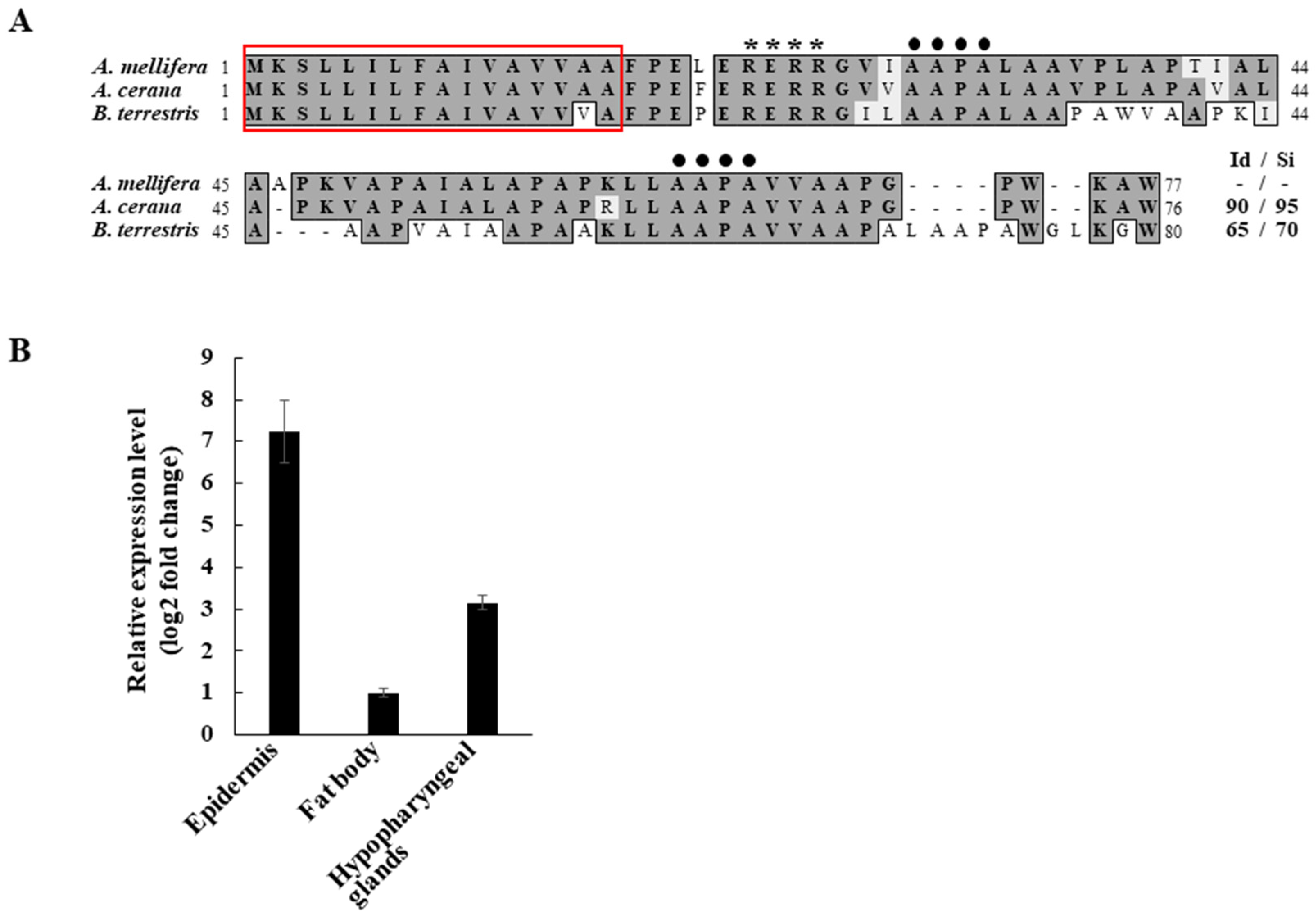
2.2. Peptide Sequence Analysis
2.3. RNA Extraction, cDNA Synthesis, and Quantitative Reverse Transcription-PCR (qRT-PCR)
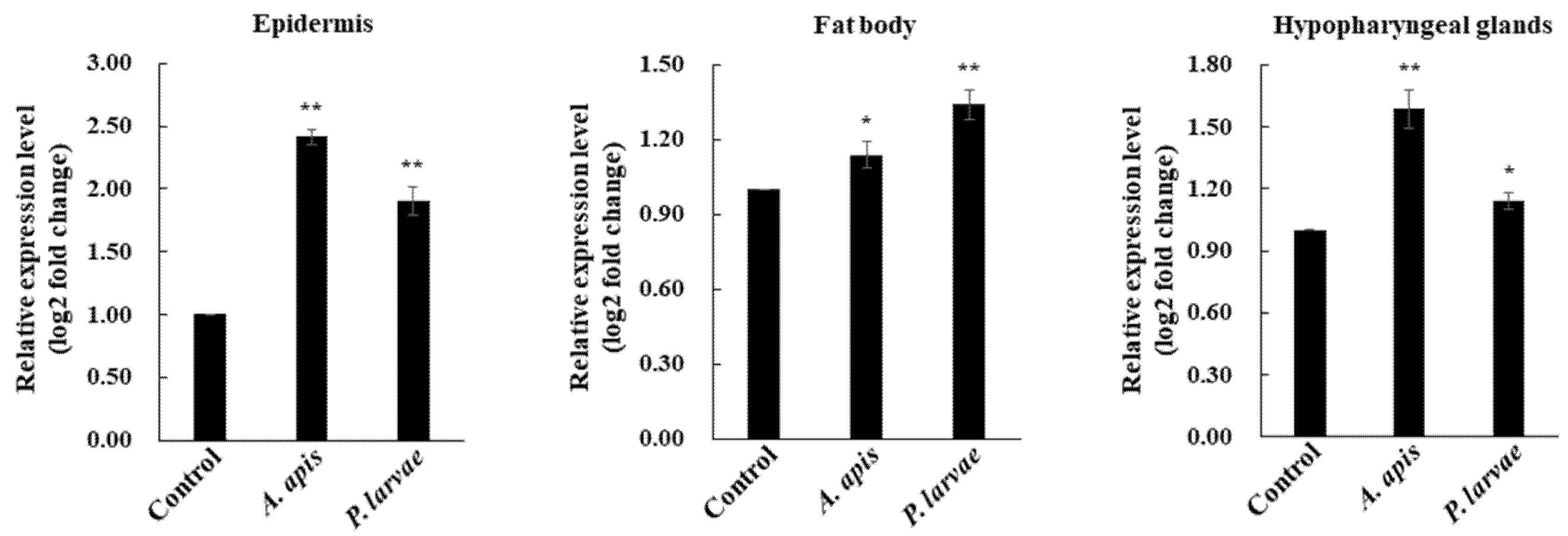
2.4. Microbial Feeding Experiment
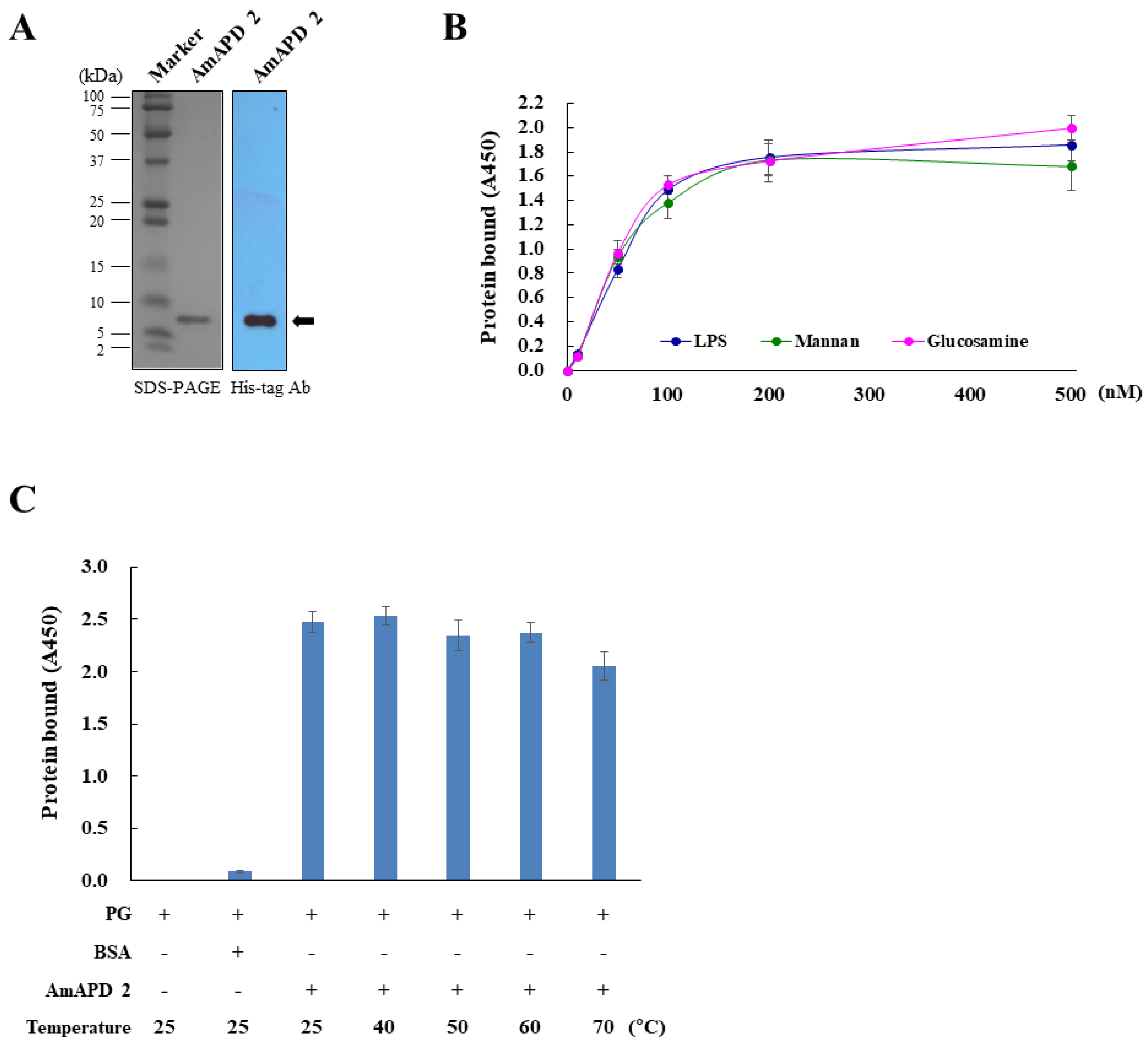
2.5. Production and Purification of Recombinant AmAPD 2
2.6. Sodium Dodecyl Sulfate-Polyacrylamide gel Electrophoresis (SDS-PAGE) and Western Blot Analysis
2.7. Binding of AmAPD 2 to Carbohydrates
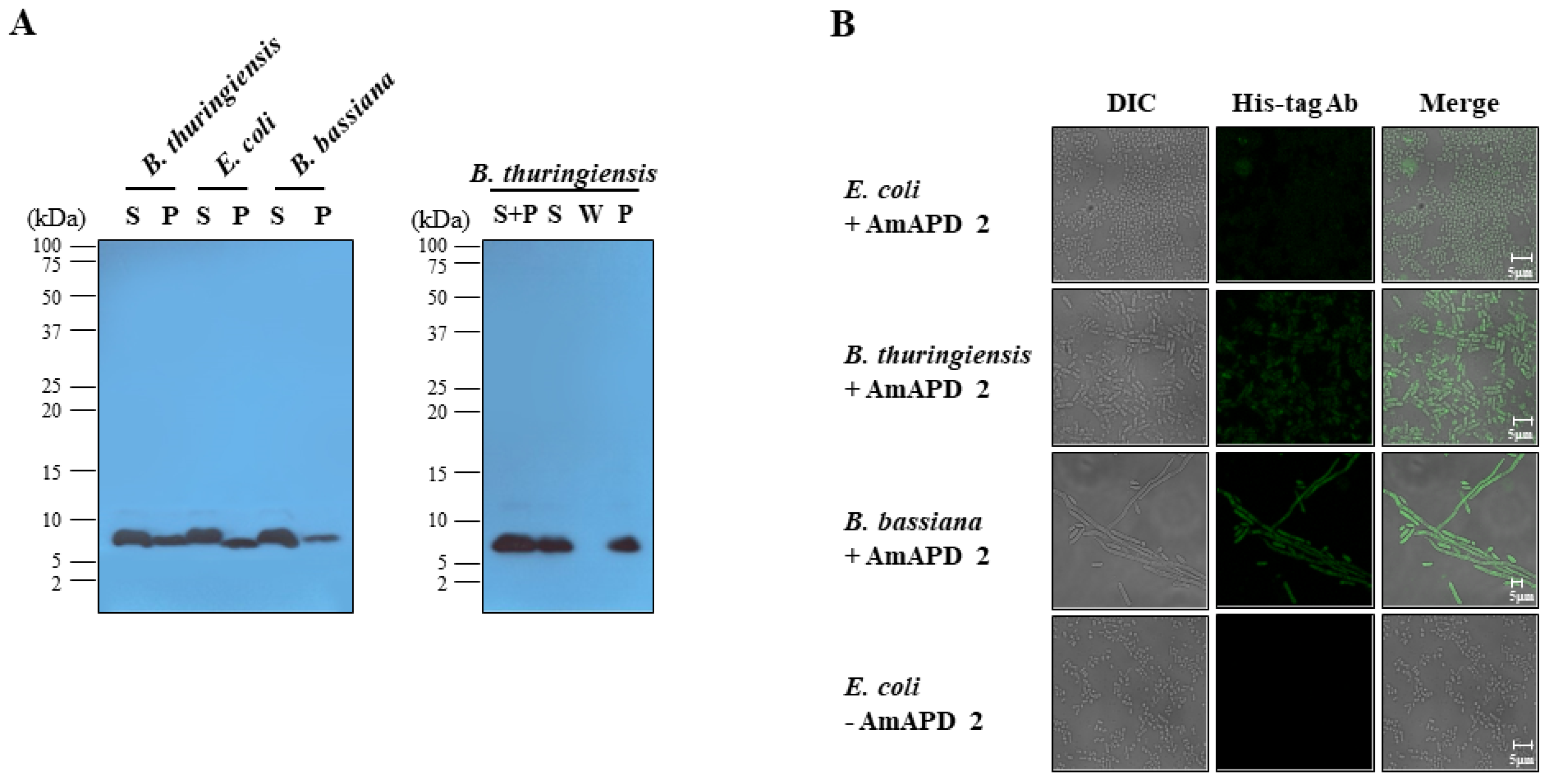
2.8. Microbial Binding Assay
2.9. Immunofluorescent Staining
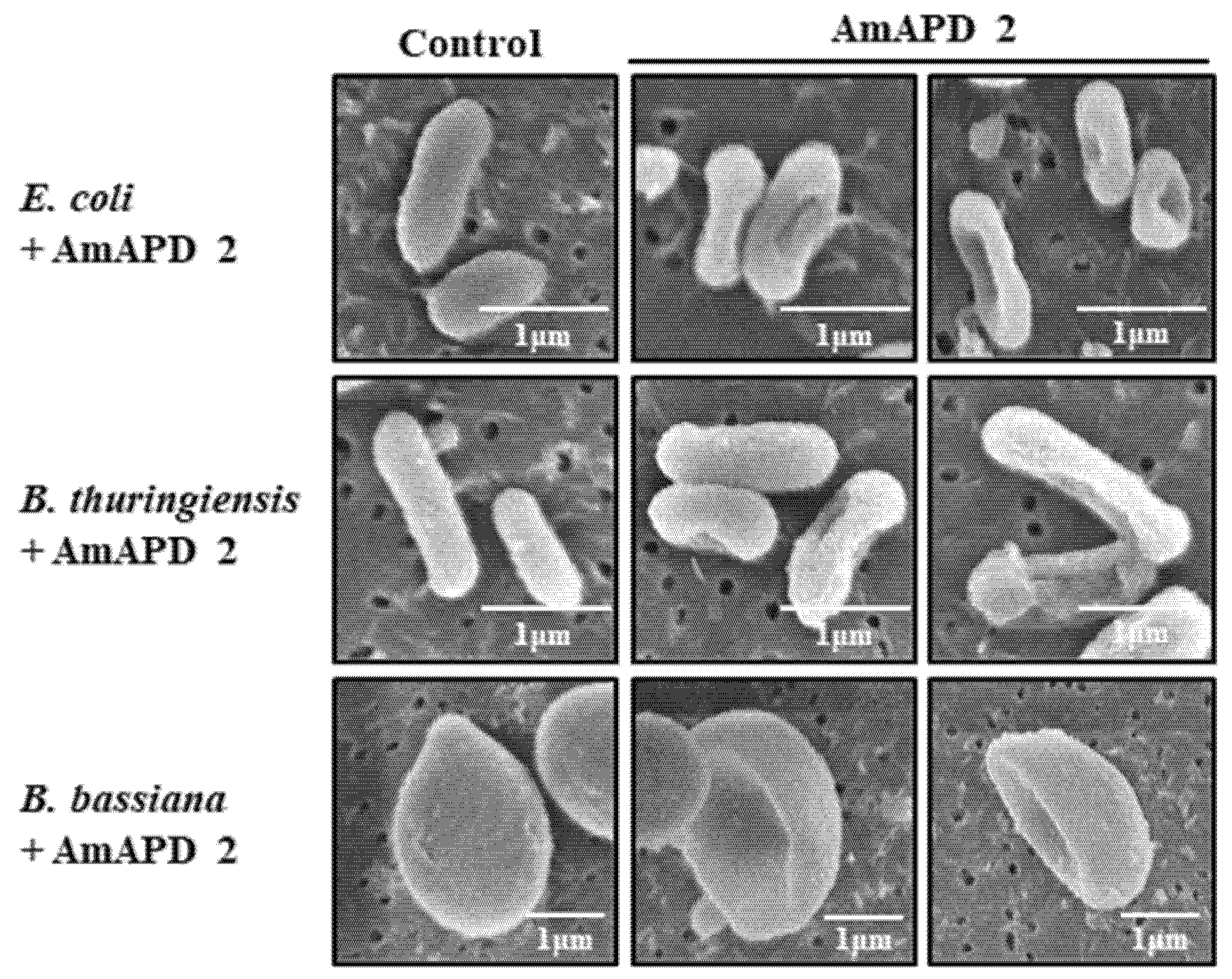
2.10. Scanning Electron Microscopy (SEM)
2.11. Antimicrobial Activity Assay
2.12. Statistical Analysis
3. Results
3.1. Expression Profile of AmAPD 2 in A. mellifera Worker Bees
3.2. Antimicrobial Activity of Recombinant AmAPD 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, S.O.; Hojrup, P.; Roepstorff, P. Insect cuticular proteins. Insect Biochem. Mol. Biol. 1995, 25, 153–176. [Google Scholar] [CrossRef]
- Willis, J.H. Structural cuticular proteins from arthropods: Annotation, nomenclature, and sequence characteristics in the genomics era. Insect Biochem. Mol. Biol. 2010, 40, 189–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, M.P.M.; Barchuk, A.R.; Simões, A.C.Q.; Cristino, A.S.; Freitas, F.C.P.; Canhos, L.L.; Bitondi, M.M.G. Genes involved in thoracic exoskeleton formation during the pupal-to-adult molt in a social insect model, Apis mellifera. BMC Genom. 2013, 15, 576. [Google Scholar] [CrossRef] [Green Version]
- Kucharski, R.; Maleszka, J.; Maleszka, R. Novel cuticular proteins revealed by the honey bee genome. Insect Biochem. Mol. Biol. 2007, 37, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Elias-Neto, M.; Soares, M.P.M.; Simões, Z.L.; Hartfelder, K.; Bitondi, M.M.G. Developmental characterization, function and regulation of a Laccase2 encoding gene in the honey bee, Apis mellifera (Hymenoptera, Apinae). Insect Biochem. Mol. Biol. 2010, 40, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Falcón, T.; Ferreira-Caliman, M.J.; Franco Nunes, F.M.; Tanaka, E.D.; do Nascimento, F.S.; Bitondi, M.M.G. Exoskeleton formation in Apis mellifera: Cuticular hydrocarbons profiles and expression of desaturase and elongase genes during pupal and adult development. Insect Biochem. Mol. Biol. 2014, 50, 68–81. [Google Scholar] [CrossRef]
- Chan, Q.W.T.; Mutti, N.S.; Foster, L.J.; Kocher, S.D.; Amdam, G.V.; Wolschin, F. The worker honeybee fat body proteome is extensively remodeled preceding a major life-history transition. PLoS ONE 2011, 6, e24794. [Google Scholar] [CrossRef] [Green Version]
- Micas, A.F.D.; Ferreira, G.A.; Laure, H.J.; Rosa, J.C.; Bitondi, M.M.G. Proteins of the integumentary system of the honeybee, Apis mellifera. Arch. Insect Biochem. Physiol. 2016, 93, 3–24. [Google Scholar] [CrossRef]
- Ward, R.; Coffey, M.; Kavanagh, K. Proteomic analysis of summer and winter Apis mellifera workers shows reduced protein abundance in winter samples. J. Insect Physiol. 2022, 139, 104397. [Google Scholar] [CrossRef]
- Werren, J.H.; Richards, S.; Desjardins, C.A.; Niehuis, O.; Gadau, J.; Colbourne, J.K.; Nasonia Genome Working Group. Functional and evolutionary insights from the genomes of three parasitoid Nasonia species. Science 2010, 327, 343–348. [Google Scholar] [CrossRef]
- Wang, J.; Jin, H.; Yang, L.; Ye, X.; Xiao, S.; Song, Q.; Stanley, D.; Ye, G.; Fang, Q. Genome-wide identification and analysis of genes encoding cuticular proteins in the endoparasitoid wasp Pteromalus puparum (Hymenoptera: Pteromalidae). Arch. Insect. Biochem. Physiol. 2020, 103, e21628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.P.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barribeau, S.M.; Sadd, B.M.; du Plessis, L.; Schmid-Hempel, P. Gene expression differences underlying genotype-by-genotype specificity in a host-parasite system. Proc. Natl. Acad. Sci. USA 2014, 111, 3496–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.R.; Kim, J.M.; Choi, H.Y.; Choi, Y.S.; Jin, B.R.; Lee, K.S.; Choi, K.H. Analysis of the gut microbiome of susceptible and resistant honeybees (Apis cerana) against sacbrood virus disease. J. Appl. Entomol. 2022. Early View. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Kim, B.Y.; Kim, J.M.; Choi, Y.S.; Lee, M.Y.; Lee, K.S.; Jin, B.R. Differential expression of major royal jelly proteins in the hypopharyngeal glands of the honeybee Apis mellifera upon bacterial ingestion. Insects 2022, 13, 334. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Park, M.J.; Kim, B.Y.; Park, H.G.; Deng, Y.; Yoon, H.J.; Choi, Y.S.; Lee, K.S.; Jin, B.R. Major royal jelly protein 2 acts as an antimicrobial agent and antioxidant in royal jelly. J. Asia-Pac. Entomol. 2019, 22, 684–689. [Google Scholar] [CrossRef]
- Je, Y.H.; Chang, J.H.; Kim, M.H.; Roh, J.Y.; Jin, B.R.; O’Reilly, D.R.; Kang, S.K. A defective viral genome maintained in Escherichia coli for the generation of baculovirus expression vectors. Biotechnol. Lett. 2001, 23, 575–582. [Google Scholar] [CrossRef]
- Kim, B.Y.; Kim, Y.H.; Park, M.J.; Yoon, H.J.; Lee, K.Y.; Kim, H.K.; Lee, K.S.; Jin, B.R. Dual function of a bumblebee (Bombus ignitus) serine protease inhibitor that acts as a microbicidal peptide and anti-fibrinolytic venom toxin. Dev. Comp. Immunol. 2022, 135, 104478. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Jin, B.R. Secapin, a bee venom peptide, exhibits anti-fibrinolytic, anti-elastolytic, and anti-microbial activities. Dev. Comp. Immunol. 2016, 63, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, K.S.; Kim, B.Y.; Choi, Y.S.; Yoon, H.J.; Jia, J.; Jin, B.R. Anti-fibrinolytic and anti-microbial activities of a serine protease inhibitor from honeybee (Apis cerana) venom. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2017, 201, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, K.S.; Ok, M.; Kim, B.Y.; Jin, B.R. Antimicrobial activity of major royal jelly protein 8 and 9 of honeybee (Apis mellifera) venom. J. Asia-Pac. Entomol. 2022, 25, 101964. [Google Scholar] [CrossRef]
- Chen, K.; Liu, C.; He, Y.; Jiang, H.; Lu, Z. A short-type peptidoglycan recognition protein from the silkworm: Expression, characterization and involvement in the prophenoloxidase activation pathway. Dev. Comp. Immunol. 2014, 45, 1–9. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, K.S.; Kim, S.R.; Gui, Z.Z.; Kim, Y.S.; Yoon, H.J.; Kim, I.; Kang, P.D.; Sohn, H.D.; Jin, B.R. A novel cellulase gene from the mulberry longicorn beetle, Apriona germari: Gene structure, expression, and enzymatic activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 140, 551–560. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria. Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Sjaarda, C. Honey glycoproteins containing antimicrobial peptides, jelleins of the major royal jelly protein 1, are responsible for the cell wall lytic and bactericidal activities of honey. PLoS ONE 2015, 10, e120238. [Google Scholar] [CrossRef] [Green Version]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Chalk, R.; Albuquerque, C.M.; Ham, P.J.; Townson, H. Full sequence and characterization of two insect defensins: Immune peptides from the mosquito Aedes aegypti. Proc. Biol. Sci. 1995, 261, 217–221. [Google Scholar]
- Axen, A.; Carlsson, A.; Engstrom, A.; Bennich, H. Gloverin, an antibacterial protein from the immune hemolymph of Hyalophora pupae. Eur. J. Biochem. 1997, 247, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Yun, E.Y.; Goo, T.W. Antimicrobial activity of an extract of Hermetia illucens larvae immunized with Lactobacillus casei against Salmonella species. Insects 2020, 11, 704. [Google Scholar] [CrossRef] [PubMed]
- Park, H.G.; Kyung, S.S.; Lee, K.S.; Kim, B.Y.; Choi, Y.S.; Yoon, H.J.; Kwon, H.W.; Je, Y.H.; Jin, B.R. Dual function of a bee (Apis cerana) inhibitor cysteine knot peptide that acts as an antifungal peptide and insecticidal toxin. Dev. Comp. Immunol. 2014, 47, 247–253. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | MIC50 (µM) | |
|---|---|---|
| Gram-positive bacterium | B. thuringiensis | 9.59 ± 0.24 |
| Gram-negative bacterium | E. coli | 7.90 ± 0.12 |
| IC50 (µM) | ||
| Entomopathogenic fungus | B. bassiana | 23.8 ± 0.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.-Y.; Kim, Y.-H.; Choi, Y.-S.; Lee, M.-Y.; Lee, K.-S.; Jin, B.-R. Antimicrobial Activity of Apidermin 2 from the Honeybee Apis mellifera. Insects 2022, 13, 958. https://doi.org/10.3390/insects13100958
Kim B-Y, Kim Y-H, Choi Y-S, Lee M-Y, Lee K-S, Jin B-R. Antimicrobial Activity of Apidermin 2 from the Honeybee Apis mellifera. Insects. 2022; 13(10):958. https://doi.org/10.3390/insects13100958
Chicago/Turabian StyleKim, Bo-Yeon, Yun-Hui Kim, Yong-Soo Choi, Man-Young Lee, Kwang-Sik Lee, and Byung-Rae Jin. 2022. "Antimicrobial Activity of Apidermin 2 from the Honeybee Apis mellifera" Insects 13, no. 10: 958. https://doi.org/10.3390/insects13100958