
The Transcription of Flight Energy Metabolism Enzymes Declined with Aging While Enzyme Activity Increased in the Long-Distance Migratory Moth, Spodoptera frugiperda

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Sampling
2.2. cDNA Library Preparation and Sequencing
2.3. Quality Control and Assembly
2.4. Differential Expression Analysis and Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.5. Validation of RNAseq Sequencing Data by Using Real-Time Quantitative PCR
2.6. Effect of Age on Enzyme Activities
2.7. Statistical Analyses
3. Results
3.1. Summary of Sequencing Quality
3.2. Transcriptome-Based Discovery of Genes Encoding Flight Energy Metabolic Enzymes and Flight Muscle Structural Proteins
3.3. Overview of Aging-Associated Transcriptional Changes
3.4. Aging-Associated Expressional Changes in Flight Energy Metabolic Enzymes and Flight Muscle Structural Proteins

3.5. Aging-Associated Expressional Changes in Other Gene Groups
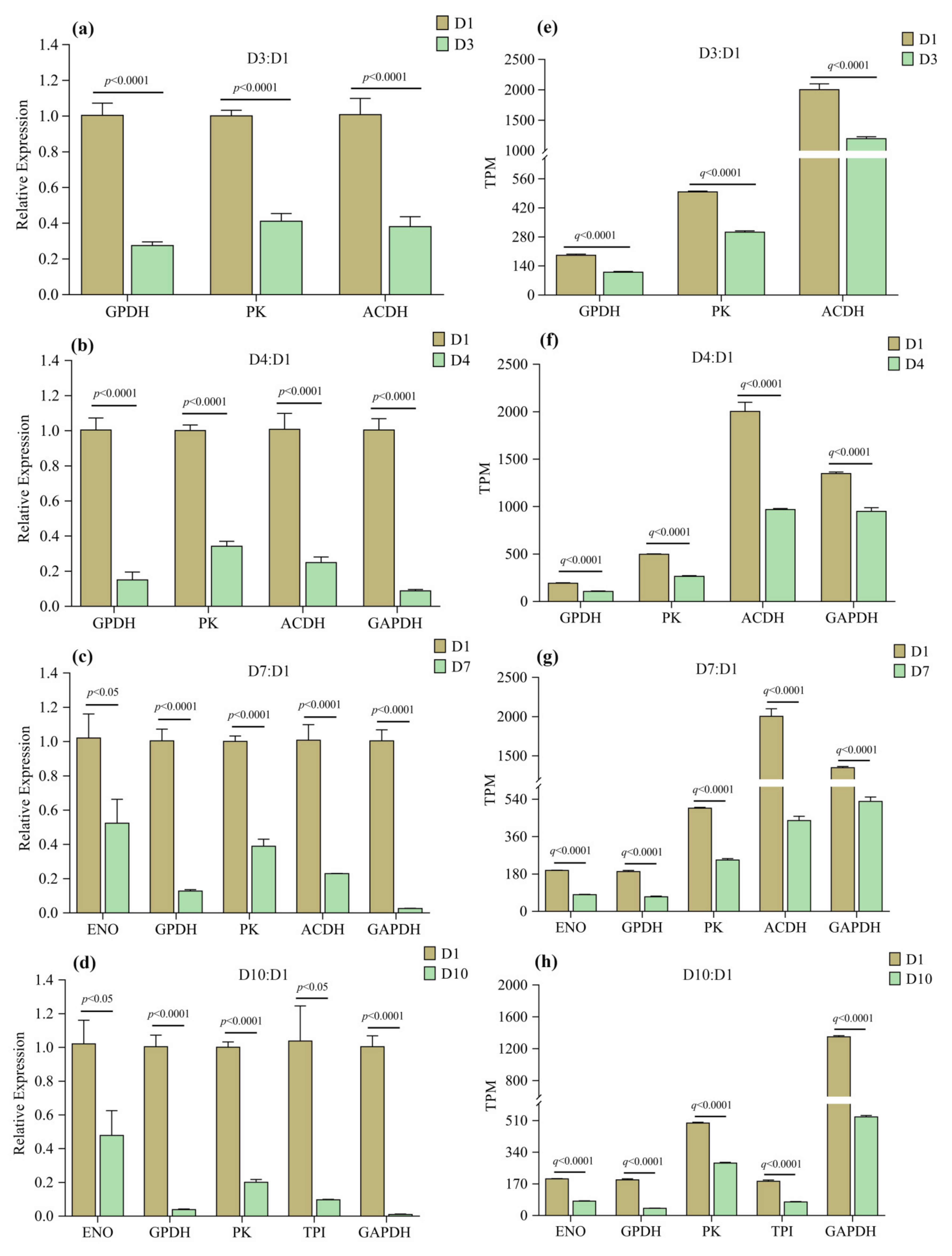
3.6. Validation of RNAseq Results by qPCR
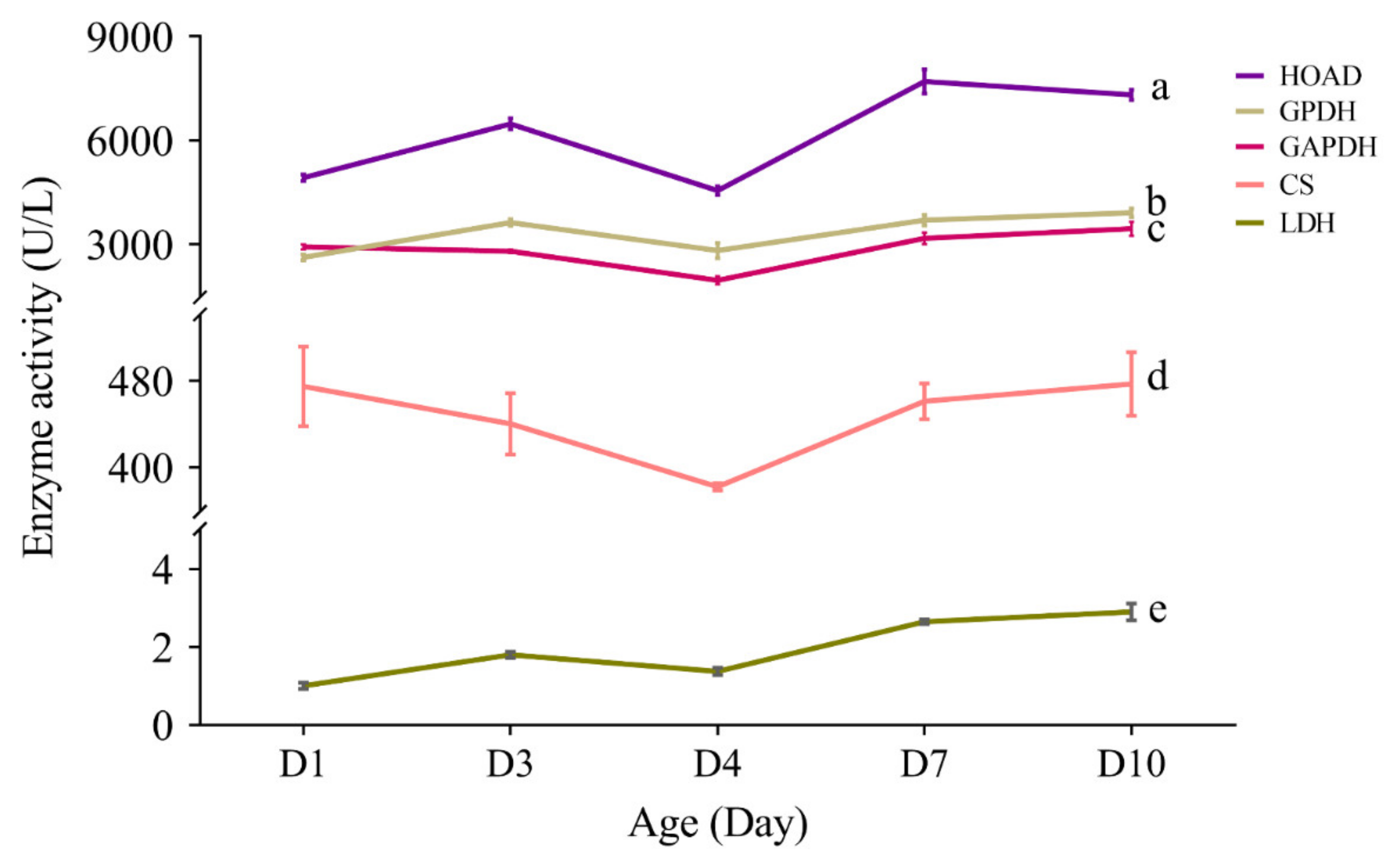
3.7. Effect of Aging on the Activity of Flight Energy Metabolic Enzymes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vigoreaux, J.O. Nature’s Versatile Engine: Insect Flight Muscle Inside and Out; Springer Science+Business Media, Inc.: Georgetown, Texas, USA, 2006. [Google Scholar]
- Glaeser, G.; Paulus, H.F.; Nachtigall, W. The Evolution of Flight; Springer International Publishing AG: Cham, Switzerland, 2017. [Google Scholar]
- Tiegs, O.W. The flight muscles of insects-their anatomy and histology; with some observations on the structure of striated muscle in general. Philos. T. Roy. Soc. B 1955, 2238, 221–347. [Google Scholar]
- Maughan, D.W.; Vigoreaux, I.O. An integrated view of insect flight muscle: Genes, motor molecules, and motion. News Physiol. Sci. 1999, 14, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ayer, G.; Vigoreaux, J.O. Flightin is a myosin rod binding protein. Cell Biochem. Biophys 2003, 38, 41–54. [Google Scholar] [CrossRef]
- Clayton, J.D.; Cripps, R.M.; Sparrow, J.C.; Bullard, B. Interaction of troponin-H and glutathione S-transferase-2 in the indirect flight muscles of Drosophila melanogaster. J. Muscle. Res. Cell M. 1998, 19, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Beenakkers, A.M.T.; Vanderhorst, D.J.; Vanmarrewijk, W.J.A. Insect flight-muscle metabolism. Insect. Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Thompson, S.N.; Suarez, R.K. Metabolism. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: New York, NY, USA, 2009; pp. 623–627. [Google Scholar]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Gade, G.; Auerswald, L. Beetles’ choice—Proline for energy output: Control by AKHs. Comp. Biochem. Phys. B 2002, 132, 117–129. [Google Scholar] [CrossRef]
- Bartke, A.; Brannan, S.; Hascup, E.; Hascup, K.; Darcy, J. Energy metabolism and ging. World J. Mens. Health 2021, 39, 222–232. [Google Scholar] [CrossRef]
- Wone, B.W.M.; Kinchen, J.M.; Kaup, E.R.; Wone, B. A procession of metabolic alterations accompanying muscle senescence in Manduca sexta. Sci. Rep. 2018, 8, 1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindle, A.G.; Lawler, J.M.; Campbell, K.L.; Horning, M. Muscle aging and oxidative stress in wild-caught shrews. Comp. Biochem. Physiol. B 2010, 155, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Chen, P.; Ye, H.; Yuan, R.; Wang, X.; Xu, J. Flight capacity of Bactrocera dorsalis (Diptera: Tephritidae) adult females based on flight mill studies and flight muscle ultrastructure. J. Insect Sci. 2015, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.-L.; Wang, X.-W.; Yang, S.; Chen, P. Changes in the activities of enzymes related to energy metabolism in flight muscles of adult Bactrocera dorsalis (Diptera: Tephritidae) at different ages and during tethered flight. Acta Entomol. Sin. 2015, 58, 471–478. [Google Scholar]
- Evenden, M.L.; Whitehouse, C.M.; Sykes, J. Factors influencing flight capacity of the mountain pine beetle (Coleoptera: Curculionidae: Scolytinae). Environ. Entomol. 2014, 43, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.; St-Pierre, J. PGC1 alpha and mitochondrial metabolism—Emerging concepts and relevance in ageing and neurodegenerative disorders. J. Cell Sci. 2012, 125, 4963–4971. [Google Scholar] [CrossRef] [Green Version]
- Wohlrab, H. Age-related changes in the flight muscle mitochondria from the blowfly Sarcophaga bullata. J. Gerontol. 1976, 31, 257–263. [Google Scholar] [CrossRef]
- Perez-Gomez, A.; Buxbaum, J.N.; Petrascheck, M. The aging transcriptome: Read between the lines. Curr. Opin. Neurobiol. 2020, 63, 170–175. [Google Scholar] [CrossRef]
- Eckley, D.M.; Coletta, C.E.; Orlov, N.V.; Wilson, M.A.; Iser, W.; Bastian, P.; Lehrmann, E.; Zhang, Y.Q.; Becker, K.G.; Goldberg, I.G. Transcriptome states reflect imaging of aging states. J. Gerontol. A biol. 2018, 73, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Tumasian, R.A.; Harish, A.; Kundu, G.; Yang, J.H.; Ubaida-Mohien, C.; Gonzalez-Freire, M.; Kaileh, M.; Zukley, L.M.; Chia, C.W.; Lyashkov, A.; et al. Skeletal muscle transcriptome in healthy aging. Nat. Commun. 2021, 12, 2014. [Google Scholar] [CrossRef]
- Arora, R.; Siddaraju, N.K.; Manjunatha, S.S.; Sudarshan, S.; Fairoze, M.N.; Kumar, A.; Chhabra, P.; Kaur, M.; Sreesujatha, R.M.; Ahlawat, S.; et al. Muscle transcriptome provides the first insight into the dynamics of gene expression with progression of age in sheep. Sci. Rep. 2021, 11, 22360. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium: Fall Armyworm Portal. Available online: https://www.cabi.org/isc/fallarmyworm (accessed on 26 November 2020).
- Congdon, B.S.; Webster, C.G.; Severtson, D.; Spafford, H. In-field capable loop-mediated isothermal amplification detection of Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae using a rapid and simple crude extraction technique. J. Econ. Entomol. 2021, 114, 2610–2614. [Google Scholar] [CrossRef]
- Guo, Z.B.; Jiang, R.X.; Tang, Y.L.; Gu, R.C.; Li, Q.Y.; Xing, T.; Xiang, L.; Wu, Y.Y.; Hu, Y.; Liu, X.; et al. Identification of new islolates of gut bacteria of Spodoptera frugiperda feeding on sorghum in Chongqing area. J. Southwest Univ. 2019, 41, 9–16. [Google Scholar]
- Nagoshi, R.N.; Nagoshi, B.Y.; Canarte, E.; Navarrete, B.; Solorzano, R.; Garces-Carrera, S. Genetic characterization of fall armyworm (Spodoptera frugiperda) in Ecuador and comparisons with regional populations identify likely migratory relationships. PLoS ONE 2019, 14, e0222332. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Yu, H.; Xu, J.; Li, Y.-H.; Ye, H. Flight muscle structure and flight capacity of females of the long-distance migratory armyworm, Spodoptera frugiperda: Effect of aging and reproduction and trade-offs between flight and fecundity. Entomol. Exp. Appl. 2022; in press. [Google Scholar]
- Li, W.; Zou, W.J.; Wang, L.H. The bionomics and control of Prodenia litura in Kunming. Southwest China J. Agric. Sci. 2006, 19, 85–89. [Google Scholar]
- Dong, Q.-J.; Zhou, J.-C.; Zhu, K.-H.; Z-T, Z.; Dong, H. A simple method for identifiying sexuality of Spodoptera frugiperd (J. E. Smith) pupae and adults. Plant Prot. 2019, 45, 96–98. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR gene indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [Green Version]
- Storey, J.D. The positive false discovery rate: A Bayesian interpretation and the q-value. Ann. Stat. 2003, 31, 2013–2035. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tevatiya, S.; Kumari, S.; Sharma, P.; Rani, J.; Chauhan, C.; Das De, T.; Pandey, K.C.; Pande, V.; Dixit, R. Molecular and functional characterization of trehalase in the mosquito Anopheles stephensi. Front. Physiol. 2020, 11, 575718. [Google Scholar] [CrossRef]
- Shakeel, M.; Du, J.; Li, S.W.; Zhou, Y.J.; Sarwar, N.; Bukhari, S.A.H. Characterization, knockdown and parental effect of hexokinase gene of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae) revealed by RNA interference. Genes 2020, 11, 1258. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C. Insect Physiology; China Agricultural Press: Beijing, China, 2004. [Google Scholar]
- Rose, I.A. Mechanism of the aldose-ketose isomerase reactions. Adv. Enzymol. Relat. Areas Mol. Biol. 1975, 43, 491–517. [Google Scholar] [PubMed]
- Hassan, A.; Huang, Q.; Xu, H.; Wu, J.; Mehmood, N. Silencing of the phosphofructokinase gene impairs glycolysis and causes abnormal locomotion in the subterranean termite Reticulitermes chinensis Snyder. Insect Mol. Biol. 2021, 30, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.A.; Hy, M.; Honegger, A.; Rose, K.; Brenner-Holzach, O. Fructose-1,6-bisphosphate aldolase from Drosophila melanogaster: Primary structure analysis, secondary structure prediction, and comparison with vertebrate aldolases. Arch. Biochem. Biophys. 1988, 266, 10–31. [Google Scholar] [CrossRef]
- Gavilane, F.G.; Gavilane, J.G.; Martin-Dudoigeon, R. Triosephosphate isomerase from the insect ceratitis-capitata molecular and enzymic properties. Comp. Biochem. Physiol. B Comp. Biochem. 1981, 70, 257–262. [Google Scholar] [CrossRef]
- Pelley, J.W. Glycolysis and pyruvate oxidation. In Elsevier’s Integrated Biochemistry; Pelley, J.W., Ed.; Mosby: Philadelphia, Pennsylvania, 2007; pp. 47–53. [Google Scholar]
- Wang, F.; Li, R.-H.; Ying, G.R. Research progress in phosphoglycerate mutase. Chin. J. Schisto. Control. 2012, 24, 353–357. [Google Scholar]
- Zhang, Z.; Qi, J.; Shang, R.; Chen, H.; Zhang, Y.; Shen, Z. Cloning and expression characteristics of pyruvate kinase gene of Nosema bombycis. Sciof. Sericult. 2019, 45, 212–217. [Google Scholar]
- Wang, D.-H.; QIin, Z.; Han, G.-B.; Wang, S.-G.; Tang, B.; ZHang, F. Cloning and sequence analysis of lactic dehydrogenase gene from Harmonia axyridis (Coleoptera: Coccinellinae). China. J. Biol. Contr. 2011, 27, 458–463. [Google Scholar]
- Tortoriello, G.; Beiersdorf, J.; Romani, S.; Williams, G.; Cameron, G.A.; Mackie, K.; Williams, M.J.; Di Marzo, V.; Keimpema, E.; Doherty, P.; et al. Genetic manipulation of sn-1-diacylglycerol lipase and CB1 cannabinoid receptor gain-of-function uncover neuronal 2-linoleoyl glycerol signaling in Drosophila melanogaster. Cannab. Cannabin. Res. 2021, 6, 119–136. [Google Scholar] [CrossRef]
- Tan, X.-L. Characterization of an Arabidopsis Long Chain Fatty Acyl-Coenzyme a Synthetase, Which Is Required for Seedling Establishment without Exogenous Sugar. Ph.D. Thesis, Northwest Agriculture and Forestry University, Xianyang, China, 2003. [Google Scholar]
- Hiltunen, J.K.; Qin, Y. β-oxidation—Strategies for the metabolism of a wide variety of acyl-CoA esters. Bioch. Biophys. Acta 2000, 1484, 117–128. [Google Scholar] [CrossRef]
- Stec, N.; Saleem, A.; Darveau, C.A. Proline as a sparker metabolite of oxidative metabolism during the flight of the bumblebee, Bombs impatiens. Metabolites 2021, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-H. Comparison and Development of Measuring Methods for Pyruvate Dehydrogenase Complex. Master’s Thesis, Central China Normal University, Wuhan, China, 2007. [Google Scholar]
- Ye, S.-C.; Yao, X.-H.; Wang, K.-L.; Lin, P.; Gong, H.-E.; Zhuo, R.-Y. Cloning and expression analysis of citrate synthase (CS) gene in Camellia oleifera. Bul. Bot. Res. 2016, 36, 9. [Google Scholar]
- Zhang, W.-L.; Shen, W.-B.; Ye, M.-B.; Xu, L.-L. Aconitase and its physiological roles in plants. J. Plant Physiol. 2003, 39, 391–398. [Google Scholar]
- Wang, H.-Y.; Zhao, Y.; Liu, X.-K. Research progress of isocitrate dehydrogenase inhibitors. Chin. J. New Drug. 2017, 26, 1272–1278. [Google Scholar]
- Wang, X. Catalytic functions and mechanisms of key enzymes in the pathway of α-ketoglutaric acid to succinic acid in cyanobacteria. Master Thesis, Jiangxi Agricultural University, Nanchang, China, 2019. [Google Scholar]
- Phillips, D.; Aponte, A.M.; French, S.A.; Chess, D.J.; Balaban, R.S. Succinyl-CoA synthetase is a phosphate target for the activation of mitochondrial metabolism. Biochemistry 2009, 48, 7140–7149. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jian, K.-L.; Liu, X.-G.; Li, X.-N. Effects of exercise on creatine kinase, lactate dehydrogenase and succinate dehydrocgenase in blood. J. PLA Inst. Phys. Edu. 2000, 25–30. [Google Scholar]
- Wang, B.; Hou, S.-T.; Chen, L.; Wang, Z. Research progress on the structure and function of fumarases. Anhui Agricul. Sci. Bul. 2009, 15, 55–57. [Google Scholar]
- Wang, X.-Y.; Wang, B.; Hou, S.-T.; Zhu, G.-P. Structure and function of malate dehydrogenase. J. Biol. 2009, 26, 69–72. [Google Scholar]
- Roosterman, D.; Cottrell, G.S. Connecting pyruvate carboxylase and citrate synthase to the flow of energy and material. Mol. Sci. 2021, 22, 6–17. [Google Scholar]
- Shelud’ko, N.S.; Matusovskaya, G.G.; Permyakova, T.V.; Matusovsky, O.S. Twitchin, a thick-filament protein from molluscan catch muscle, interacts with F-actin in a phosphorylation-dependent way. Arch. Biochem. Biophys. 2004, 432, 269–277. [Google Scholar] [CrossRef]
- Pu, Y.; Hai-Zhong, Y.; Jia-An, C.; Zeng-Rong, Z. The proteins in flight muscles of insects. Inst. Appl. Entom. 2005, 42, 726–731. [Google Scholar]
- Vigoreaux, J.O.; Saide, J.D.; Valgeirsdottir, K.; Pardue, M.L. Flightin, a novel myofibrillar protein of Drosophila stretch-activated muscles. J. Cell Bio. 1993, 121, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.-F.; Li, X.-d. Locusta migratoria flight muscle troponin partially activates thin filament in a calcium-dependent manner. Insect Mol. Biol. 2022, 31, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Ojima, K. Myosin: Formation and maintenance of thick filaments. Anim. Sci. J. 2019, 90, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Rynkiewicz, M.J.; Prum, T.; Hollenberg, S.; Kiani, F.A.; Fagnant, P.M.; Marston, S.B.; Trybus, K.M.; Fischer, S.; Moore, J.R.; Lehman, W. Tropomyosin must interact weakly with actin to effectively regulate thin filament function. Biophys. J. 2017, 113, 2444–2451. [Google Scholar] [CrossRef] [Green Version]
- Kabsch, W.; Vandekerckhove, J. Structure and function of actin. Annu. Rev. Bioph. Biom. 1992, 21, 49–76. [Google Scholar] [CrossRef]
- Soto-Adames, F.N.; Alvarez-Ortiz, P.; Vigoreaux, J.O. An evolutionary analysis of flightin reveals a conserved motif unique and widespread in Pancrustacea. J. Mol. Evol. 2014, 78, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, B.; Newsholme, E.A. Comparative aspects of fuel utilization and metabolism by muscle. In Insect Muscle; Usherwood, P.N.R., Ed.; Academic Press: London, UK, 1975. [Google Scholar]
- Sullivan, D.T.; MacIntyre, R.; Fuda, N.; Fiori, J.; Barrilla, J.; Ramizel, L. Analysis of glycolytic enzyme co-localization in Drosophila flight muscle. J. Exp. Biol. 2003, 206, 2031–2038. [Google Scholar] [CrossRef] [Green Version]
- Beenakkers, A.M. Carbohydrate and fat as a fuel for insect flight: A comparative study. J. Insect Physiol. 1969, 15, 353–361. [Google Scholar] [CrossRef]
- Kim, K.-N.; Sin, U.-C.; Yun, C.-N.; Song, H.-S.; Huang, Z.-J.; Huang, Q.-Y.; Lei, C.-L. Influence of green light illumination on several enzymes involved in energy metabolism in the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). J. Asia-Pacific Entomol. 2019, 22, 487–492. [Google Scholar] [CrossRef]
- Li, K.; Luo, L. Activities of enzymes in the flight muscle of pupal and adult oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 1999, 42, 37–43. [Google Scholar]
- Gingras, A.C.; Raught, B.; Sonenberg, N. eIF4 initiation factors: Effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem. 1999, 68, 913–963. [Google Scholar] [CrossRef] [PubMed]
- Syntichaki, P.; Troulinaki, K.; Tavernarakis, N. eIF4E function in somatic cells modulates ageing in Caenorhabditis elegans. Nature 2007, 445, 922–926. [Google Scholar] [CrossRef]
- Ham, S.; Lee, S.-J.V. Advances in transcriptome analysis of human brain aging. Exp. Mol. Med. 2020, 52, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Ioannidou, A.; Goulielmaki, E.; Garinis, G.A. DNA damage: From chronic inflammation to age-related deterioration. Front. Genet. 2016, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Gorbunova, V.; Seluanov, A.; Mao, Z.; Hine, C. Changes in DNA repair during aging. Nucleic Acids Res. 2007, 35, 7466–7474. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID * | Gene Name | Abbreviation | Function | Reference |
|---|---|---|---|---|
| The glycolysis pathway | ||||
| LOC118280632 | Trehalose | TRE | Catalyzing the conversion of trehalose to glucose. | [34] |
| LOC118276330 | Hexokinase | HK | Catalyzes the conversion of glucose to glucose 6-phosphate (G6P). | [35] |
| LOC118265182 | Glycogen phosphorylase | GP | Phosphorylation of glycogen to glucose 1-phosphate (G1P). | [36] |
| —— | Glucose phosphate mutase | PGM | Catalytic conversion of G1P to G6P. | [9] |
| LOC118277306 | Glucose-6-phosphate isomerase | GPI | Rearrange G6P into 6-phosphate-fructose (F6P). | [37] |
| LOC118270306 | Phosphofructose kinase | PFK | Catalyzing F6P to produce fructose 1,6-diphosphate (FDP). | [38] |
| LOC118275099 | Fructose-bisphosphate aldolase | FBA | FDP was decomposed into glyceraldehyde 3-phosphate (GAP) and dihydroxyacetone phosphate (DHAP). | [39] |
| LOC118268149 | Glycerol-3-phosphate dehydrogenase | GPDH | Catalytic conversion between DHAP and glycerol 3-phosphate (G3P). | [15] |
| LOC118263360 | Triose phosphate isomerase | TPI | Catalytic conversion between GAP and DHAP. | [40] |
| LOC118271716 | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | Catalytic conversion of GAP to 1,3-diphosphoglyceride. | [15] |
| LOC118262502 | Phosphoglycerate kinase | PGK | Catalyzes the conversion of 1,3-diphosphoglyceride to 3-phosphoglycerate. | [41] |
| LOC118274617 | Phosphoglycerate mutase | PGAM | Catalytic conversion of 3-phosphoglyceric acid to 2-phosphoglyceric acid. | [42] |
| LOC118268804 | Enolase | ENO | Catalytic conversion of 2-phosphoglyceric acid to phosphoenolpyruvate. | [41] |
| LOC118281561 | Pyruvate kinase | PK | Catalyzes phosphoenolpyruvate to transfer high-energy phosphate groups to ADP to produce ATP and pyruvate (PYR). | [43] |
| LOC118272480 | Lactate dehydrogenas | LDH | Catalyze the reduction and oxidation reaction between PYR and lactic acid (LA). | [44] |
| The fat oxidation pathway | ||||
| LOC118266592 | Diacylglycerol lipase | DGL | Catalyze the hydrolysis of Diacylglycerol lipid (DG-LP) to release free fatty acids (FFA) and monoacylglycerol. | [45] |
| —— | Diglyceride lipase | DL | Catalyzing the conversion of diglycerides to FFA. | [36] |
| LOC118262085 | Acetyl CoA synthase | ACS | Catalytic synthesis of fatty acyl CoA (FA-CoA) from FFA. | [46] |
| —— | Carnitine palmitoyl transferase | CPT | Synthesis of lipoacyl carnitine catalyzed by lipoacyl coenzyme A and carnitine. | [36] |
| LOC118273677 | Carnitine O-acetyltransferase | CAT | Transfer of lipoacyl carnitine into mitochondrial matrix. | [36] |
| LOC118274443 | Acyl-CoA dehydrogenase | ACDH | Make (Cn) acyl coenzyme A in α and β One hydrogen is removed from each carbon atom to form β-enolyl-COA. | [47] |
| LOC118271967 | Enoyl-CoA hydratase | ECH | Catalyzing trans alkenylacyl coenzyme A to produce β-hydroxyacyl COA. | [47] |
| LOC118272083 | 3-hydroxyacyl-CoA Dehydrogenase | HOAD | Catalyzing 3-hydroxyacyl coenzyme A to produce β-ketoacyl-COA. | [36] |
| LOC118265763 | Thiolase | THI | Catalyzing the formation of (Cn-2) fatty acyl CoA from β-ketoacyl-COA. | [47] |
| The proline oxidation pathway | ||||
| LOC118274501 | Proline dehydrogenase | ProDH | The first step of catalyzing the catabolism of proline is to oxidize proline to pyrrole-5-carboxylic acid. | [48] |
| LOC118262586 | Pyrroline-5-carboxylate dehydrogenase | P5CDH | Glutamic acid oxide- γ- Glutamic acid from semialdehyde. | [36] |
| LOC118268408 | Alanine aminotransferase | ALT | Catalyzing the production of glutamate and alanine α-Ketoglutarate. | [36] |
| LOC118263189 | Aspartate aminotransferase | AST | Catalyzing the production of glutamic acid and oxaloacetic acid α-Ketoglutarate. | [48] |
| Tricarboxylic acid (TCA) cycle | ||||
| LOC118269912 | Pyruvate dehydrogenase | PDH | Catalytic oxidative dehydrogenation of PYR to acetyl-CoA. | [49] |
| LOC118262833 | Citrate synthase | CS | The key enzyme at the entrance of the tricarboxylic acid cycle catalyzes the synthesis of citrate from oxaloacetate and acetyl CoA. | [50] |
| LOC118264042 | Aconitase | ACO | Catalytic citrate acid to isocitrate. | [51] |
| LOC118267252 | Isocitrate dehydrogenase | IDH | Catalytic oxidative dehydrogenation of isocitrate α-Ketoglutarate, the main factor controlling circulation. | [52] |
| —— | α- Ketoglutarate dehydrogenase | α-KGDHC | Catalysis α-Ketoglutarate to succinyl CoA. | [53] |
| —— | Succinyl COA synthase | SCS | It is a TCA cycle enzyme that catalyzes succinyl CoA to produce succinate. | [54] |
| LOC118271964 | Succinate dehydrogenase | SDH | It is a key enzyme involved in the tricarboxylic acid cycle in mitochondria and catalyzes succinate to fumarate. | [55] |
| —— | Fumarase | Fum | Catalyze the reversible hydration reaction between fumarate acid and malate. | [56] |
| LOC118269324 | Malate dehydrogenase | MDH | Conversion between malate and oxaloacetate. | [57] |
| LOC118267676 | Pyruvate carboxylase | PC | Catalyzing the carboxylation of H2CO3 and PYR to form oxaloacetic acid. | [58] |
| Protein | Gene ID | Function | Reference |
|---|---|---|---|
| Twitchin | LOC118280582 (twitchin-like) | Interacts with F-actin. | [59] |
| Tropomodulin | LOC118281560 (tropomodulin-1-like) | Terminal capping protein of actin. | [60] |
| Flightin | LOC118267000 (flightin-like) | A myosin-binding protein that maintains sarcomere stability. | [61] |
| Troponin | LOC118263234 (troponin T); LOC118263900 (troponin I); LOC118266930 (troponin C) | A sensor that receives Ga2+ to regulate muscle contraction; consists of three subunits, TnT, TnI and TnC. | [62] |
| Myosin | A total of 26 transcripts were identified. Such as LOC118278200 (myosin-11-like); LOC118278607 (myosin-14-like); LOC118279007 (myosin-7B-like); LOC118263248 (myosin-M heavy chain-like); LOC118264677 (myosin heavy chain); LOC118268178 (myosin heavy chain 95F-like); LOC118271963 (myosin-10-like) | A superfamily of molecular motors that move along the track of actin filaments, accounting for 60% of the total protein of myofibrils. Plays a role in muscle contraction. | [63] |
| Paramyosin | LOC118267608 (paramyosin-like); LOC118269023 (paramyosin, long form-like) | Play roles in myoblast fusion, myofibril assembly, and muscle contraction. | [60] |
| Connectin, Titin | LOC118268606 (titin-like, transcript variant X4); LOC118268680 (titin-like, transcript variant X1); LOC118268988 (titin-like, transcript variant X2) LOC118274312 (titin-like); LOC118267569 (connectin-like); LOC118267725 (connectin-like, transcript variant X1) | Intertwined with thick and thin filaments, making muscles elastic. | [60] |
| Tropomyosin | LOC118273937 (tropomyosin-1, transcript variant X1); LOC118273938 (tropomyosin-2, transcript variant X1) | Regulates myofibrils function by interacting with actin. | [64] |
| Actin | A total of 41 transcripts were identified. Such as LOC118279073 (actin, muscl); LOC118279603 (actin-like); LOC118279605 (actin, alpha skeletal muscle-like); LOC118280131 (actin-like protein 9A); LOC118264039 (actin-like protein 15A); LOC118264040 (actin-like protein 16A); LOC118268744 (actin-like protein 24A) | Play roles in muscle movement, accounting for 20% of the total protein of muscles. Divided into α, β and γ-actin three categories. | [65] |
| Age (Day) | GAPDH:HOAD |
|---|---|
| D1 | 0.594 |
| D3 | 0.433 |
| D4 | 0.430 |
| D7 | 0.412 |
| D10 | 0.471 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Wu, T.; Yu, H.; Xu, J.; Zhang, J.-Z.; Fu, D.-Y.; Ye, H. The Transcription of Flight Energy Metabolism Enzymes Declined with Aging While Enzyme Activity Increased in the Long-Distance Migratory Moth, Spodoptera frugiperda. Insects 2022, 13, 936. https://doi.org/10.3390/insects13100936
Fu Y, Wu T, Yu H, Xu J, Zhang J-Z, Fu D-Y, Ye H. The Transcription of Flight Energy Metabolism Enzymes Declined with Aging While Enzyme Activity Increased in the Long-Distance Migratory Moth, Spodoptera frugiperda. Insects. 2022; 13(10):936. https://doi.org/10.3390/insects13100936
Chicago/Turabian StyleFu, Yan, Ting Wu, Hong Yu, Jin Xu, Jun-Zhong Zhang, Da-Ying Fu, and Hui Ye. 2022. "The Transcription of Flight Energy Metabolism Enzymes Declined with Aging While Enzyme Activity Increased in the Long-Distance Migratory Moth, Spodoptera frugiperda" Insects 13, no. 10: 936. https://doi.org/10.3390/insects13100936