
Feeding Preferences and Responses of Monochamus saltuarius to Volatile Components of Host Pine Trees


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Chemicals
2.3. Determination of Feeding Preference for Different Host Pine Trees
2.4. Host Pine Tree Volatile Collections and GC-MS Spectrometry
2.5. Four-Arm Olfactometer Test
2.6. Statistics
3. Results
3.1. Feeding Preference on Different Host Pine Trees
3.2. Analysis of Volatile Compounds in Different Host Pine Trees
3.3. Responses to Host Pine Tree Volatiles
3.3.1. Responses to Different Species of Host Pine Tree Volatiles
3.3.2. Responses to Different Concentrations of Host Pine Tree Volatiles
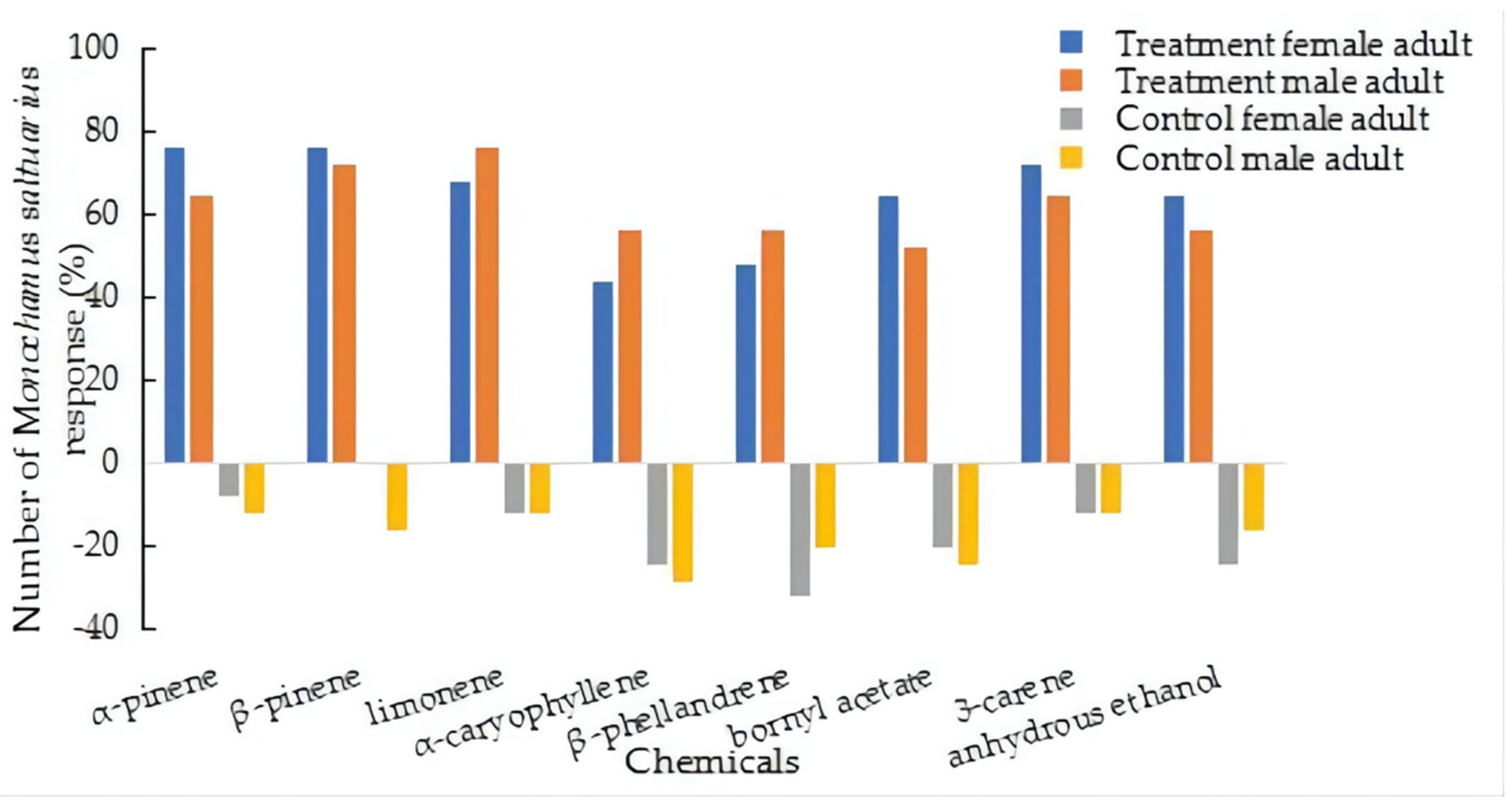
3.3.3. Responses to Different Components of Host Pine Tree Volatiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, B.J.; Pan, H.Y.; Tang, J. Bursaphelenchus xylophilus; China Forestry Publishing House: Beijing, China, 2003; pp. 1–9. [Google Scholar]
- Heisuke, S.; Takeshi, S.; Mitsunori, K. Transmission of Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle (Nematoda, Aphelenchoididae) by Monochamus saltuarius (Gebler) (Coleoptera, Cerambycidae). J. Jpn. For. Soc. 1987, 69, 492–496. [Google Scholar]
- Togashi, K.; Jikumaru, S. Horizontal transmission of Bursaphelenchus mucronatus (Nematoda: Aphelenchoididae) between insect vectors of Monochamus saltuarius (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1996, 31, 317–320. [Google Scholar] [CrossRef]
- Nakayama, Y.; Jikumaru, S.; Togashi, K. Reproductive traits and diel activity of adult Monochamus saltuarius (Coleoptera: Cerambycidae) at two different temperatures. J. For. Res. 1998, 3, 61–65. [Google Scholar] [CrossRef]
- Kwon, T.S.; Lim, J.H.; Sim, S.J.; Kwon, Y.D.; Son, S.K.; Lee, K.Y.; Kim, Y.T.; Park, J.W.; Shin, C.H.; Ryu, S.B. Distribution patterns of Monochamus alternatus and M. saltuarius (Coleoptera: Cerambycidae) in Korea. J. Korean Soc. For. Sci. 2006, 95, 543–550. [Google Scholar]
- Jung, C.S.; Koh, S.H.; Nam, Y.; Ahn, J.J.; Lee, C.Y.; Choi, W.I. A model for predicting spring emergence of Monochamus saltuarius (coleoptera: Cerambycidae) from Korean white pine Pinus koraiensis. J. Econ. Entomol. 2015, 108, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.C.; Shi, Y.; Jiang, S.W.; Zheng, Y.N. Study on the species of long-horned beetles carrying Bursaphelenchus xylophilus in Liaoning. For. Res. 2021, 34, 174–181. [Google Scholar]
- Hu, Q.; Jin, J.; Du, Y.B.; Fan, J.T. Variation of the composition of attractants in lures for Monochamus alternatus (Coleoptera: Cerambycidae) in the field and its influence on trapping efficacy. Acta Entomol. Sin. 2018, 61, 1310–1318. [Google Scholar] [CrossRef]
- Fan, J.T.; Zhang, D.Y.; Zhang, Z.P.; Meng, J.G.; Wang, Y.P. Feeding behavior of Monochamus alternatus and its relationship with the host volatiles. J. Zhejiang Agric. For. Univ. 2014, 31, 78–82. [Google Scholar] [CrossRef]
- Su, X.; Li, H.; Yan, J.; Huang, D.; Wang, Z.; Zhang, S.; Bi, Y.; Yan, A.; Lv, F. Ecological geographical distribution and pollution-free control of Apriona germari hope and Anoplophora glabripennis (Motschulsky). For. Ecol. Sci. 2020, 35, 393–398. [Google Scholar] [CrossRef]
- Yu, H.; Wu, H. The new host plant and new vector insect of Bursaphelenchus xylophilus were found in Liaoning province. For. Pest Dis. 2018, 37, 61. [Google Scholar]
- Yu, H.; Wu, H.; Huang, R.; Wang, J.; Zhang, R.; Song, R. Separation and identification of Bursaphelenchus xylophilus from Pinus sylvestris var. mongolica in Fushun city. For. Pest Dis. 2020, 39, 6–10. [Google Scholar] [CrossRef]
- Yu, H.; Wu, H.; Zhang, X.; Wang, L.; Zhang, X.; Song, Y. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. [Google Scholar] [CrossRef]
- Lee, H.R.; Lee, S.C.; Lee, D.H.; Choi, W.S.; Jung, C.S.; Jeon, J.H.; Kim, J.E.; Park, I.K. Identification of the aggregation-sex pheromone produced by male Monochamus saltuarius, a major insect vector of the pine wood nematode. J. Chem. Ecol. 2017, 43, 670–678. [Google Scholar] [CrossRef]
- Lee, S.M.; Hong, D.K.; Jang, S.H.; Lee, K.Y.; Lee, C.W. Synergistic attraction of pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) to monochamol and α-pinene. Entomol. Res. 2017, 47, 125–128. [Google Scholar] [CrossRef]
- Zheng, Y.; Fan, L.; Shi, Y.; Chen, G.; Wang, J.; Jiang, S. A comparative study on trapping effect of different attractions on Monochamus saltuariu. J. Northeast For. Univ. 2021, 49, 102–104, 115. [Google Scholar] [CrossRef]
- Lu, W.; Hou, M.; Wen, J. Effects of plant volatiles on herbivorous insects. Plant Prot. 2017, 33, 7–11. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Blomquist, G.J.; Millar, J.G.; Hanks, L.M. The role of contact pheromones in mate location and recognition in Xylotrechus colonus. J. Chem. Ecol. 2005, 31, 1443–1444. [Google Scholar] [CrossRef]
- Chenier, J.V.R.; Philogene, B.J.R. Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. J. Chem. Ecol. 1989, 15, 1729–1745. [Google Scholar] [CrossRef] [PubMed]
- Wallin, K.F.; Raffa, K.F. Altered constitutive and inducible phloem monoterpenes following natural defoliation of jack pine: Implications to host mediated interguild interactions and plant defense theories. J. Chem. Ecol. 1999, 25, 861–880. [Google Scholar] [CrossRef]
- Li, J. The VOCs Emittion of Two Tree Species of Platycladus orientalis and Pinus tabulaeformis in Urban Environment. Doctoral Dissertation, Chinses Academy of Forestry, Beijing, China, July 2009. [Google Scholar]
- Hao, D.J.; Yang, J.X.; Dai, H.G. Research progress and prospect on chemical ecology of Monochamus alternatus. Chin. J. Ecol. 2008, 27, 1227–1233. [Google Scholar] [CrossRef]
- Fan, J.T.; Sun, J.H. Influences of host volatiles on feeding behaviour of the Japanese pine sawyer, Monochamus alternatus. J. Appl. Entomol. 2006, 130, 238–244. [Google Scholar] [CrossRef]
- Hao, D.J.; Ma, F.L.; Wang, Y.; Dai, H.G.; Zhang, Y.H. Electroantennogram and behavioural responses of Monochamus alternatus to volatiles from Pinus massoniana. Chin. J. Appl. Entomol. 2007, 4, 541–544. [Google Scholar]
- Ning, T.; Fan, J.T.; Fang, Y.L.; Sun, J.H. Changes in contents of host volatile terpenes under different damaged states and electroantennogram response of Monochamus alternatus hope to these volatiles. Acta Entomol. Sin. 2006, 2, 179–188. [Google Scholar] [CrossRef]
- Fan, J.T.; Kang, L.; Sun, J.H. Role of host volatiles in mate location by the Japanese pine sawyer, Monochamus alternatus hope (Coleoptera: Cerambycidae). Environ. Entomo. 2007, 36, 58–63. [Google Scholar] [CrossRef]
- Li, S.Q.; Zhang, Z.N. EAG response of Monochamus alternatus hope (Coleoptera: Cerambycidae) to colatiles from larval frass and the repellency tests in fields. Acta Entomol. Sin. 2008, 3, 284–289. [Google Scholar] [CrossRef]
- Jiang, W.J.; Ji, B.Z.; Liu, S.W.; Song, J. Advances in the studies on semiochemicals and olfactory receptor mechanism in adults of cerambycid beetles. Acta Entomol. Sin. 2005, 3, 427–436. [Google Scholar] [CrossRef]
- Fan, H.; Jin, Y.; Li, J.; Chen, H. Advances on plant volatile semiochemicals attracting herbivorous insects. J. Beijing For. Univ. 2004, 26, 76–81. [Google Scholar] [CrossRef]
- Agelopoulos, N.G.; Chamberlain, K.; Pickett, J.A. Factors affecting volatile emissions of intact potato plants, Solanum tuberosum: Variability of quantities and stability of ratios. J. Chem. Ecol. 2000, 26, 497–511. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Pine koraiensis | Pine tabulaeformis | Pine sylvestris var. mongholica | Larix olgensis | ||||
|---|---|---|---|---|---|---|---|---|
| Chemicals | Relative Content (%) | Chemicals | Relative Content (%) | Chemicals | Relative Content (%) | Chemicals | Relative Content (%) | |
| 1 | α-pinene | 9.26 | α-pinene | 21.29 | α-pinene | 46.16 | α-pinene | 10.53 |
| 2 | 3-ethylbenzaldehyde | 5.74 | β-pinene | 2.58 | d-limonene | 5.35 | 5-hydroxy-4-octanone | 5.28 |
| 3 | β-pinene | 4.06 | 1-tetradecene | 2.06 | 2-tetradecene | 3.86 | 3-ethylbenzaldehyde | 4.76 |
| 4 | 3-carene | 3.66 | 2-tetradecene | 1.86 | 3-ethylbenzaldehyde | 3.64 | 4-ethylacetophenone | 4.41 |
| 5 | 4-ethylacetophenone | 3.21 | 4-Hydroxy-3-methylacetophenone | 1.73 | camphene | 3.13 | 2-tetradecene | 2.44 |
| 6 | myrcene | 2.51 | sabinene | 1.47 | β-pinene | 2.73 | β-pinene | 2.22 |
| 7 | 2-ethylbenzaldehyde | 2.31 | 1,4-diacetylbenzene | 1.38 | sabinene | 1.68 | 4-ethylbenzaldehyde | 1.70 |
| 8 | 4-isopropylbenzyl alcohol | 2.27 | 3-nitrostyrene | 1.23 | 4-ethylacetophenone | 1.66 | sabinene | 1.69 |
| 9 | limonene | 1.95 | 2-tert-butyl-4-methylphenol | 1.19 | 4-ethylbenzaldehyde | 1.41 | cis-3-hexen-1-ol | 1.44 |
| 10 | camphene | 1.70 | camphene | 0.78 | myrcene | 1.31 | 1,3-dimethylcyclopentanol | 1.28 |
| 11 | 1-dodecene | 1.37 | 1,4-diethylbenzene | 0.63 | 3-carene | 1.06 | camphene | 0.52 |
| 12 | di(2-ethylhexyl)phthalate | 1.36 | 1,5,6,7-tetrahydro-4 h-indol-4-one | 0.62 | 4-carene | 0.69 | 3-carene | 0.51 |
| 13 | 1,2-diethylbenzene | 0.49 | 5-nitro-m-xylene | 0.55 | butyl acetate | 0.58 | ||
| 14 | 2-hexanol | 0.40 | 4-nitrostyrene | 0.54 | L-bornyl acetate | 0.58 | ||
| 15 | 1,4-diethylbenzene | 0.29 | 3-ethylbenzaldehyde | 0.41 | butyl acrylate | 0.63 | ||
| 16 | 1-ethenyl-3-ethylbenzene | 0.27 | butyl propionate | 0.44 | ||||
| 17 | 3-hexanol | 0.24 | ||||||
| 18 | 4-ethylpropiophenone | 0.22 | ||||||
| 19 | 3-hexanone | 0.21 | ||||||
| 20 | 2-hexanone | 0.18 | ||||||
| Chemical Compound | Dilution Ratio | CK | No-Response | Reaction Rate | Selection Coefficient | ||
|---|---|---|---|---|---|---|---|
| 10 | 100 | 1000 | |||||
| Component I | 9 Female 5 Male | 6 Female 7 Male | 4 Female 3 Male | 2 Female 2 Male | 4 Female 8 Male | 76% | 78.95% |
| Component Ⅱ | 11 Female 9 Male | 5 Female 4 Male | 3 Female 3 Male | 1 Female 2 Male | 5 Female 7 Male | 76% | 84.21% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhang, S.; Zheng, Y. Feeding Preferences and Responses of Monochamus saltuarius to Volatile Components of Host Pine Trees. Insects 2022, 13, 888. https://doi.org/10.3390/insects13100888
Wang J, Zhang S, Zheng Y. Feeding Preferences and Responses of Monochamus saltuarius to Volatile Components of Host Pine Trees. Insects. 2022; 13(10):888. https://doi.org/10.3390/insects13100888
Chicago/Turabian StyleWang, Jue, Sufang Zhang, and Yanan Zheng. 2022. "Feeding Preferences and Responses of Monochamus saltuarius to Volatile Components of Host Pine Trees" Insects 13, no. 10: 888. https://doi.org/10.3390/insects13100888