


Effect of Entomopathogenic Fungi, Beauveria bassiana (Cordycipitaceae), on the Bark Beetle, Ips typographus (L.), under Field Conditions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
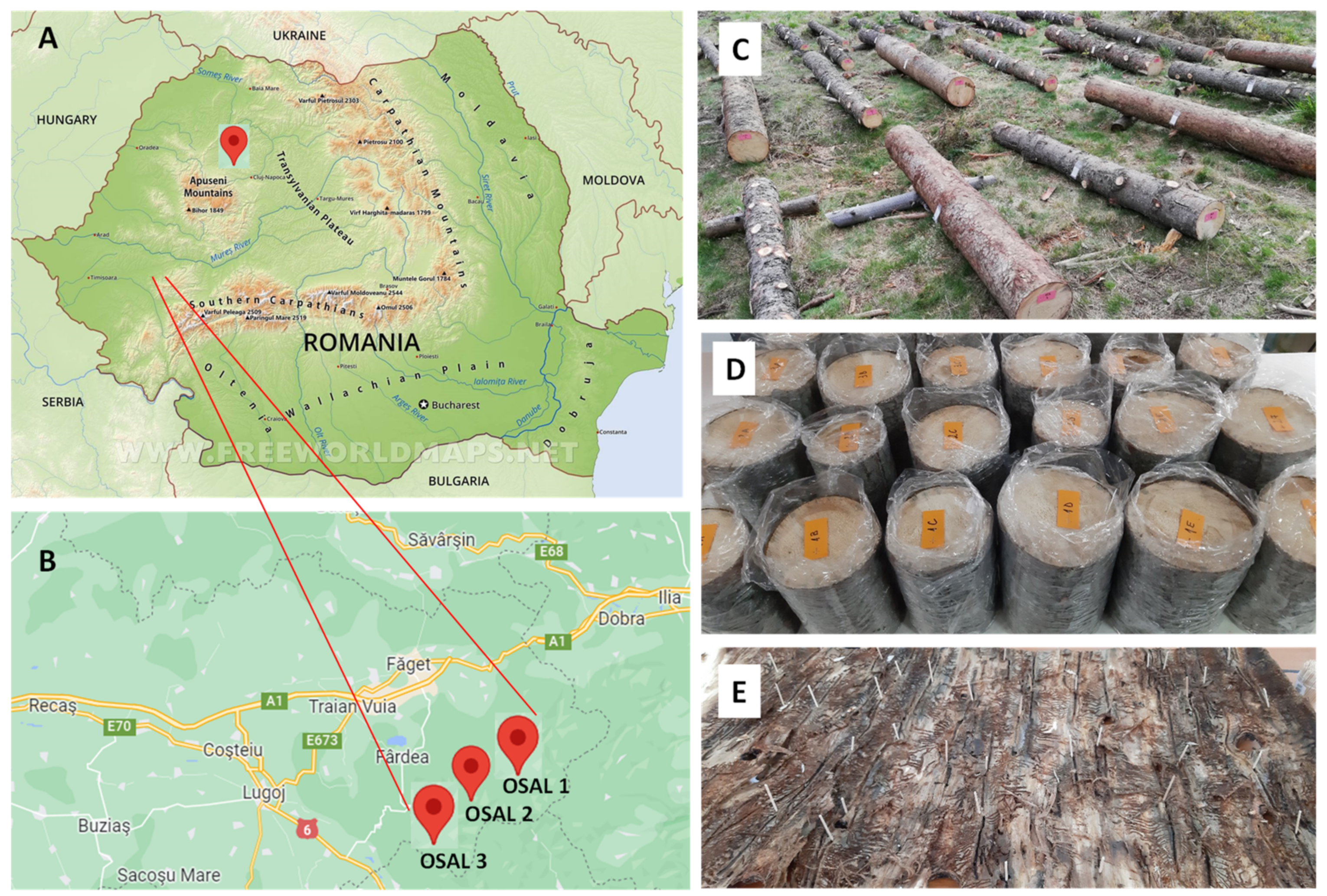
2. Material and Methods
2.1. Data Collection
2.2. Data Analyses
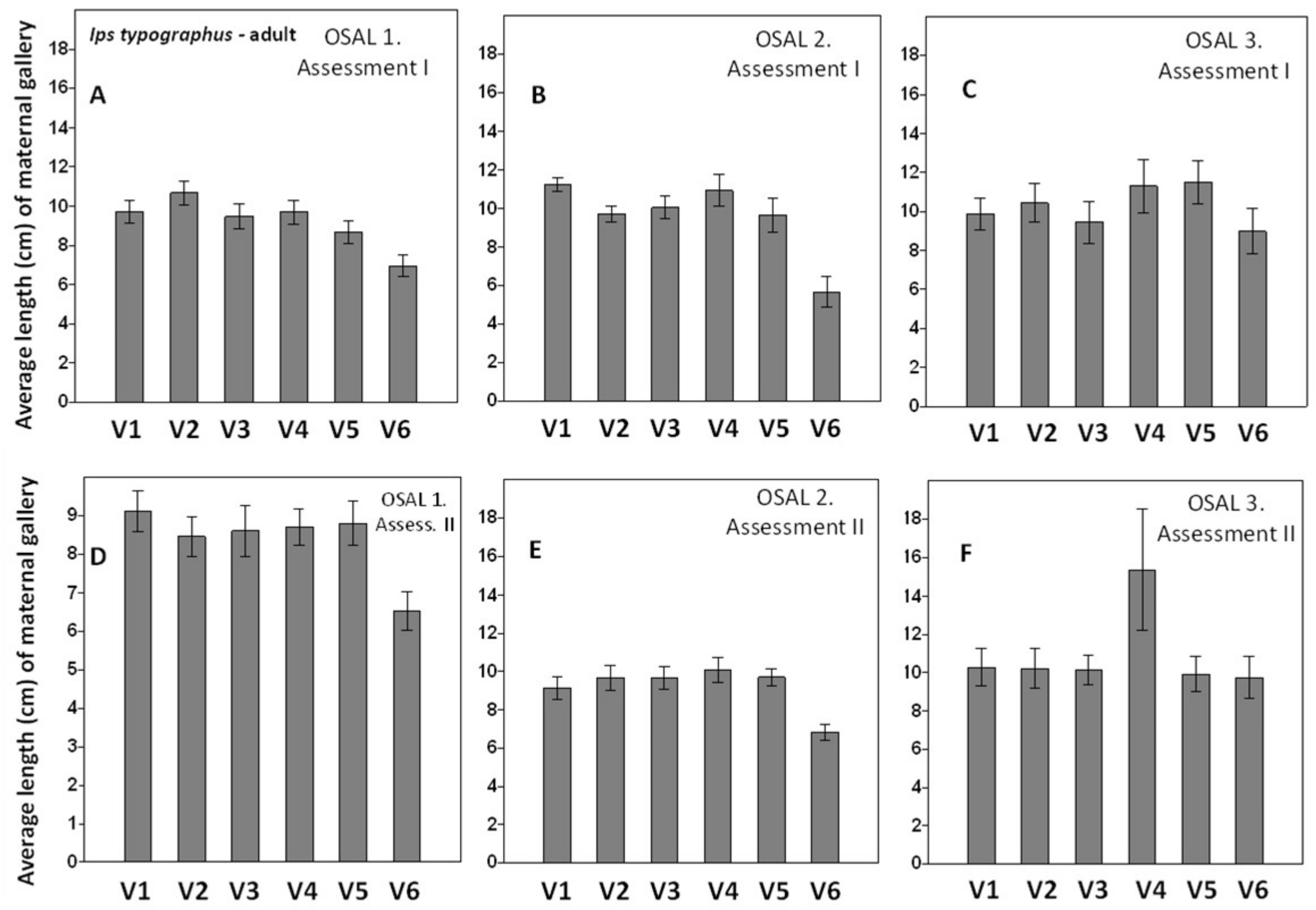
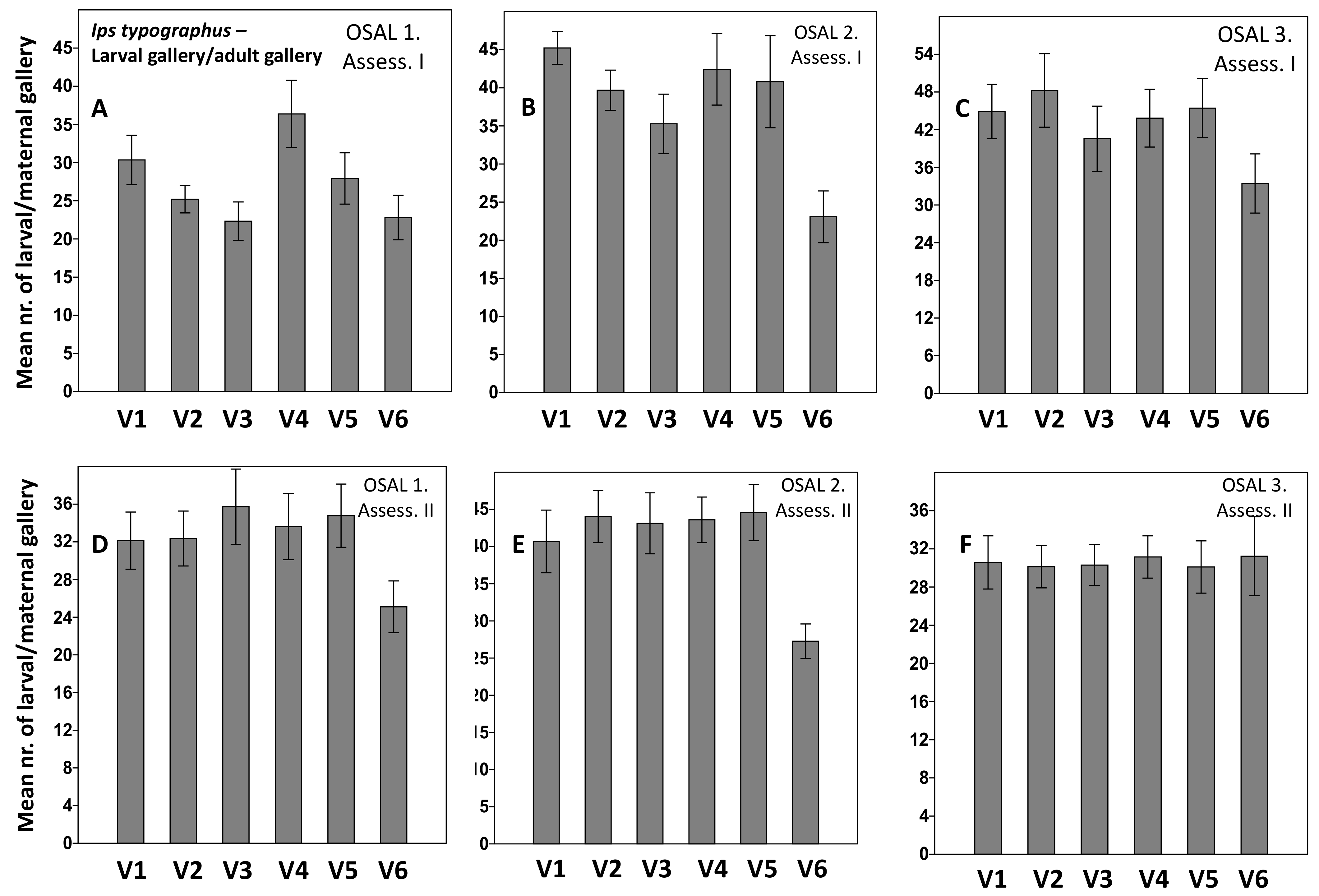
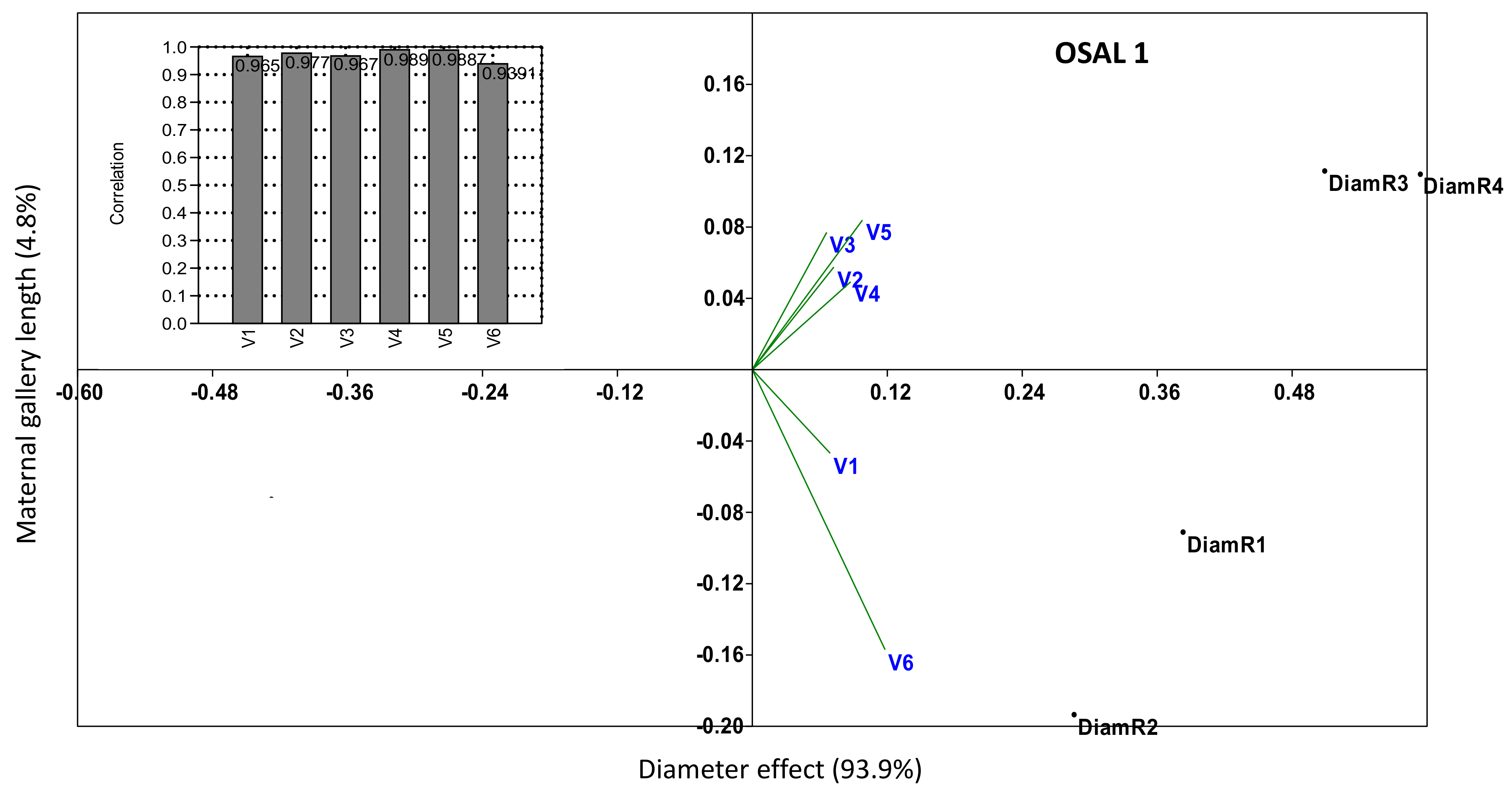
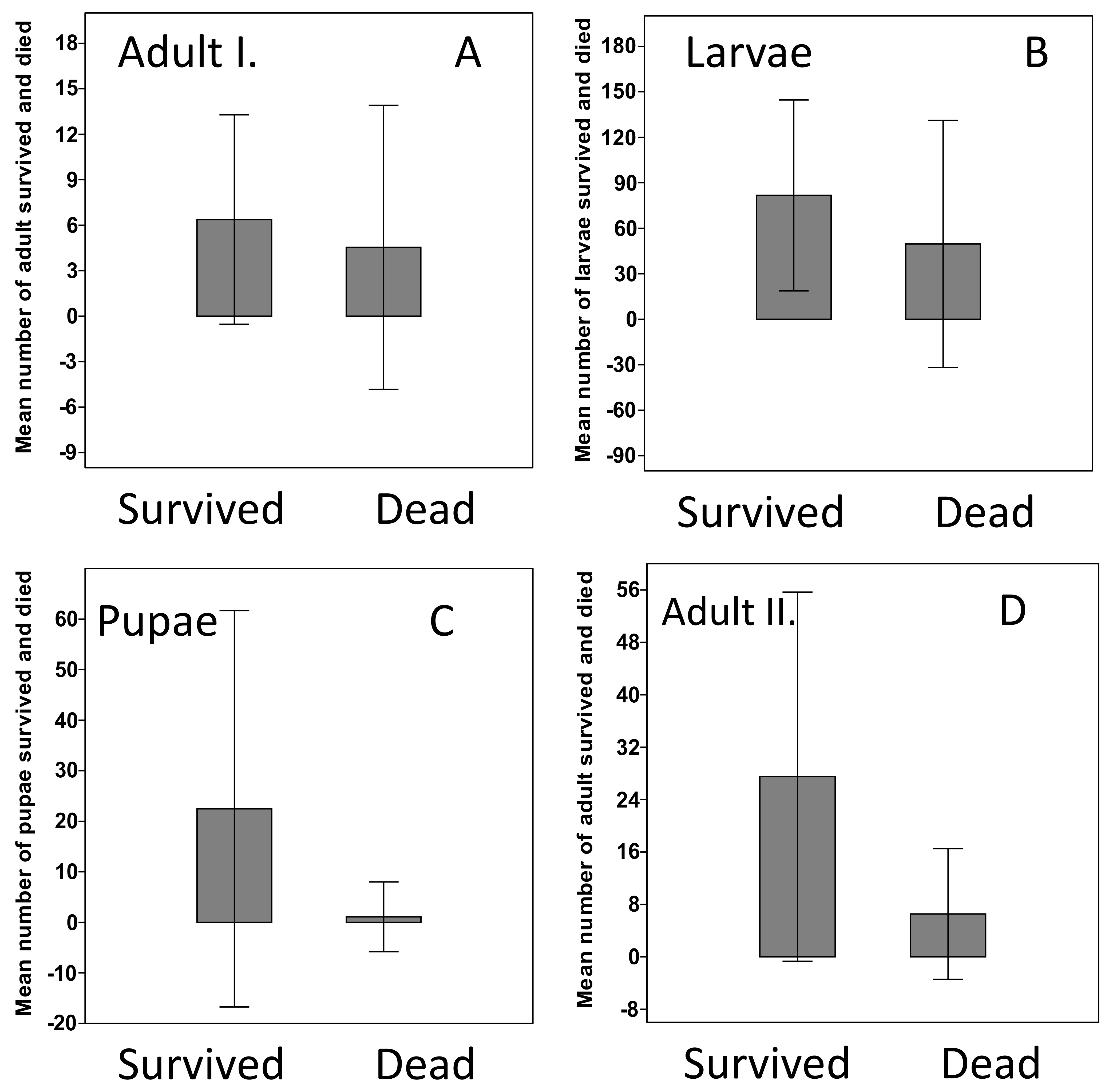
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess Forest Mortality Is Consistently Linked to Drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef] [PubMed]
- Sendall, K.M.; Reich, P.B.; Zhao, C.; Jihua, H.; Wei, X.; Stefanski, A.; Rice, K.; Rich, R.L.; Montgomery, R.A. Acclimation of Photosynthetic Temperature Optima of Temperate and Boreal Tree Species in Response to Experimental Forest Warming. Glob. Change Biol. 2015, 21, 1342–1357. [Google Scholar] [CrossRef] [PubMed]
- Senf, C.; Seidl, R. Storm and Fire Disturbances in Europe: Distribution and Trends. Glob. Change Biol. 2021, 27, 3605–3619. [Google Scholar] [CrossRef] [PubMed]
- Wegensteiner, R.; Stradner, A.; Händel, U. Occurrence of Pathogens in Ips Typographus (Coleoptera: Curculionidae) and in Other Spruce Bark Beetles from the Wilderness Reserve Dürrenstein (Lower Austria). Biologia 2014, 69, 92–100. [Google Scholar] [CrossRef]
- Kreutz, J.; Zimmermann, G.; Vaupel, O. Horizontal Transmission of the Entomopathogenic Fungus Beauveria Bassiana among the Spruce Bark Beetle, Ips Typographus (Col., Scolytidae) in the Laboratory and under Field Conditions. Biocontrol. Sci. Technol. 2004, 14, 837–848. [Google Scholar] [CrossRef]
- Batta, Y.A. Biocontrol of Almond Bark Beetle (Scolytus Amygdali Geurin-Meneville, Coleoptera: Scolytidae) Using Beauveria Bassiana (Bals.) Vuill. (Deuteromycotina: Hyphomycetes). J. Appl. Microbiol. 2007, 103, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Reay, S.D.; Brownbridge, M.; Cummings, N.J.; Nelson, T.L.; Souffre, B.; Lignon, C.; Glare, T.R. Isolation and Characterization of Beauveria Spp. Associated with Exotic Bark Beetles in New Zealand Pinus Radiata Plantation Forests. Biol. Control. 2008, 46, 484–494. [Google Scholar] [CrossRef]
- Yan, Z.; Sun, J.; Don, O.; Zhang, Z. The Red Turpentine Beetle, Dendroctonus Valens LeConte (Scolytidae): An Exotic Invasive Pest of Pine in China. Biodivers. Conserv. 2005, 14, 1735–1760. [Google Scholar] [CrossRef]
- Krischik, V.A.; Davidson, J. IPM (Integrated Pest Management) of Midwest Landscapes. In Minnesota Agricultural Experiment Station (MAES); Minnesota Agricultural Experiment Station (MAES): St. Paul, MN, USA, 2004. [Google Scholar]
- Simionescu, A. Protecţia Pădurilor; Muşatinii: Suceava, Romania, 2000. [Google Scholar]
- Simionescu, A.; Mihalache, G.; Mihalciuc, V.; Ciornei, C.; Chira, D.; Olenici, N.; Lupu, D.; Nețoiu, C.; Vlăduleasa, A.; Iliescu, M.; et al. Protecția Pădurilor; Mușatinii: Suceava, Romania, 2003. [Google Scholar]
- Yeo, H.; Pell, J.K.; Alderson, P.G.; Clark, S.J.; Pye, B.J. Laboratory Evaluation of Temperature Effects on the Germination and Growth of Entomopathogenic Fungi and on Their Pathogenicity to Two Aphid Species. Pest. Manag. Sci. 2003, 59, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Bugeme, D.M.; Maniania, N.K.; Knapp, M.; Boga, H.I. Effect of Temperature on Virulence of Beauveria Bassiana and Metarhizium Anisopliae Isolates to Tetranychus Evansi. Exp. Appl. Acarol. 2008, 46, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.J.; Davis, T.S. Plant Secondary Metabolites and Low Temperature Are the Major Limiting Factors for Beauveria Bassiana (Bals.-Criv.) Vuill. (Ascomycota: Hypocreales) Growth and Virulence in a Bark Beetle System. Biol. Control. 2020, 141, 104130. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Nemčovič, M.; Šalamon, M.; Hyblerova, S.; Mazáň, M.; Salamon, I. Entomopathogenic Fungus Species Beauveria Bassiana (Bals.) and Metarhizium Anisopliae (Metsch.) Used as Mycoinsecticide Effective in Biological Control of Ips Typographus (L.). J. Microbiol. Biotechnol. Food Sci. 2013, 2, 2469. [Google Scholar]
- Zúbrik, M.; Raši, R.; Vakula, J.; Varínsky, J.; Nikolov, C.; Novotný, J. Bark Beetle (Ips Typographus L., Pityogenes Chalcographus L., Col.: Scolytidae) Pheromone Traps Spatial Distribution Optimisation in Central Slovakian Mountains. Lesn. Časopis-For. J. 2008, 2008, 235–248. [Google Scholar]
- Steinwender, B.; Krenn, H.; Wegensteiner, R. Different Effects of the Insectpathogenic Fungus Beauveria Bassiana (Deuteromycota) on the Bark Beetle Ips Sexdentatus (Coleoptera: Curculionidae) and on Its Predator Thanasimus Formicarius (Coleoptera: Cleridae). J. Plant. Dis. Prot. 2010, 117, 33–38. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fora, C.G.; Boja, N.; Moatăr, M.; Tóth, F.; Balog, A. Effect of Entomopathogenic Fungi, Beauveria bassiana (Cordycipitaceae), on the Bark Beetle, Ips typographus (L.), under Field Conditions. Insects 2022, 13, 885. https://doi.org/10.3390/insects13100885
Fora CG, Boja N, Moatăr M, Tóth F, Balog A. Effect of Entomopathogenic Fungi, Beauveria bassiana (Cordycipitaceae), on the Bark Beetle, Ips typographus (L.), under Field Conditions. Insects. 2022; 13(10):885. https://doi.org/10.3390/insects13100885
Chicago/Turabian StyleFora, Ciprian George, Nicușor Boja, Mihaela Moatăr, Ferenc Tóth, and Adalbert Balog. 2022. "Effect of Entomopathogenic Fungi, Beauveria bassiana (Cordycipitaceae), on the Bark Beetle, Ips typographus (L.), under Field Conditions" Insects 13, no. 10: 885. https://doi.org/10.3390/insects13100885