
Inheritance of Resistance to Cry1A.105 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sources of H. zea Populations
2.2. Genetic Crosses
2.3. Diet Overlay Bioassays with Cry1A.105 Toxin
2.4. Data Analysis
3. Results
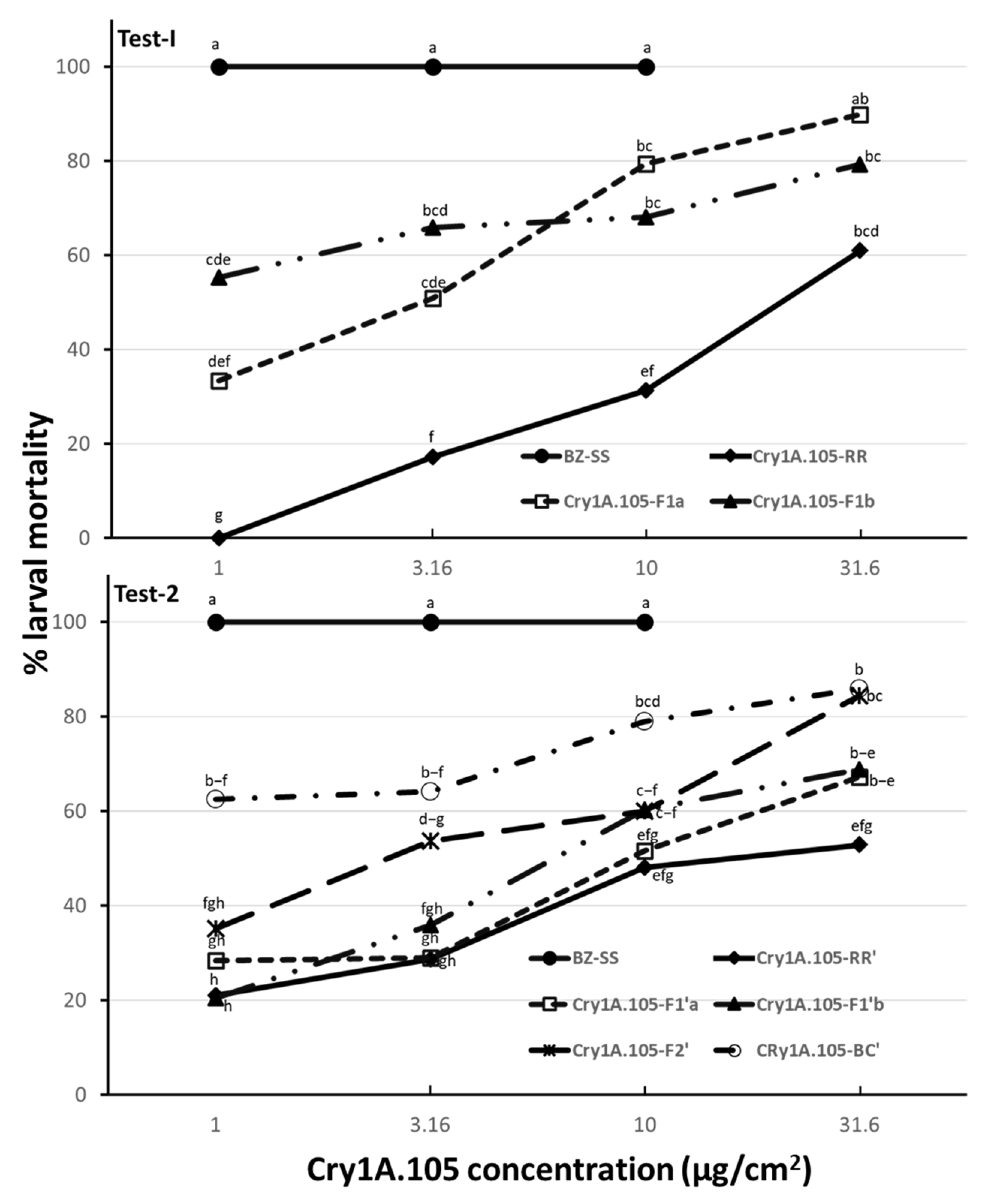
3.1. F2 Screen-Isolated Cry1A.105 Resistance Was Autosomal in H. zea
3.2. F2 Screen-Isolated Cry1A.105 Resistance Was Nonrecessive in H. zea
3.3. F2 Screen-Isolated Cry1A.105 Resistance Was Monogenic in H. zea
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- US-EPA (U.S. Environmental Protection Agency). Biopesticide Registration Action Document: Bacillus thuringiensis Cry1A.105 and Cry2Ab2 Insecticidal Proteins and the Genetic Material Necessary for Their Production in Corn. 2010. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/pip/mon-89034-brad.pdf (accessed on 1 August 2022).
- Siebert, M.W.; Nolting, S.P.; Hendrix, W.; Dhavala, S.; Craig, C.; Leonard, B.R.; Stewart, S.D.; All, J.; Musser, F.; Buntin, G.D.; et al. Evaluation of corn hybrids expressing Cry1F, Cry1A.105, Cry2Ab2, Cry34Ab1/Cry35Ab1, and Cry3Bb1 against southern United States insect pests. J. Econ. Entomol. 2012, 105, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Rule, D.M.; Nolting, S.P.; Prasifka, P.L.; Storer, N.P.; Hopkins, B.W.; Scherder, E.F.; Siebert, M.W.; Hendrix, W.H., III. Efficacy of pyramided Bt proteins Cry1F, Cry1A.105, and Cry2Ab2 expressed in SmartStax corn hybrids against lepidopteran insect pests in the northern United States. J. Econ. Entomol. 2014, 107, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Head, G.P.; Leonard, B.R.; Levy, R.; Niu, Y.; Huang, F. A challenge for the seed mixture refuge strategy in Bt maize: Impact of cross-pollination on an ear-feeding pest, corn earworm. PLoS ONE 2014, 9, e112962. [Google Scholar] [CrossRef]
- Yang, F.; Kerns, D.L.; Head, G.P.; Leonard, B.R.; Niu, Y.; Huang, F. Occurrence, distribution, and ear damage of Helicoverpa zea (Lepidoptera: Noctuidae) in mixed plantings of non-Bt and Bt corn containing Genuity® SmartStaxTM traits. Crop Protect. 2014, 55, 127–132. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Finkenbinder, C. Field-evolved resistance in corn earworm to Cry proteins expressed by transgenic sweet corn. PLoS ONE 2016, 11, e0169115. [Google Scholar] [CrossRef]
- Dively, G.P.; Kuhar, T.P.; Taylor, S.; Doughty, H.B.; Holmstrom, K.; Gilrein, D.; Nault, B.A.; Ingerson-Mahar, J.; Whalen, J.; Reisig, D.; et al. Sweet corn sentinel monitoring for lepidopteran field-evolved resistance to Bt toxins. J. Econ. Entomol. 2020, 114, 207–319. [Google Scholar] [CrossRef]
- Bilbo, T.R.; Reay-Jones, F.P.F.; Reisig, D.D.; Greene, J.K. Susceptibility of corn earworm (Lepidoptera: Noctuidae) to Cry1A.105 and Cry2Ab2 in North and South Carolina. J. Econ. Entomol. 2019, 112, 1845–1857. [Google Scholar] [CrossRef]
- Kaur, G.; Guo, J.; Brown, S.; Head, G.P.; Price, P.A.; Paula-Moraes, S.; Ni, X.; Dimase, M.; Huang, F. Field-evolved resistance of Helicoverpa zea (Boddie) to transgenic maize expressing pyramided Cry1A.105/Cry2Ab2 proteins in northeast Louisiana, the United States. J. Invertebr. Pathol. 2019, 163, 11–20. [Google Scholar] [CrossRef]
- Yang, F.; González, J.C.S.; Williams, J.; Cook, D.C.; Gilreath, R.T.; Kerns, D.L. Occurrence and ear damage of Helicoverpa zea on transgenic Bacillus thuringiensis maize in the field in Texas, U.S. and its susceptibility to Vip3A protein. Toxins 2019, 11, 102. [Google Scholar] [CrossRef]
- Yu, W.; Lin, S.; Dimase, M.; Niu, Y.; Brown, S.; Head, G.P.; Price, P.A.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; et al. Extended investigation of field-evolved resistance of the corn earworm Helicoverpa zea (Lepidoptera: Noctuidae) to Bacillus thuringiensis Cry1A.105 and Cry2Ab2 proteins in the southeastern United States. J. Invertebr. Path. 2021, 183, 107560. [Google Scholar] [CrossRef]
- Yu, W.; Head, G.P.; Price, P.; Brown, S.; Cook, D.; Ni, X.; Reay-Jones, F.P.F.; Dimase, M.; Huang, F. Estimation of resistance allele frequencies to Cry1A.105 and Cry2Ab2 in the corn earworm (Lepidoptera: Noctuidae) with F2 isolines generated from a mass-mating method. Crop Prot. 2022, 161, 106054. [Google Scholar] [CrossRef]
- US-EPA (United States Environmental Protection Agency). White Paper on Resistance in Lepidopteran Pests of Bacillus thuringiensis (Bt) Plant Incorporated Protectants in the United States. 2018. Available online: https://www.epa.gov/sites/production/files/2018-7/documents/position_paper_07132018.pdf (accessed on 1 August 2022).
- Reay-Jones, F.P.F. Pest status and management of corn earworm (Lepidoptera: Noctuidae) in field corn in the United States. J. Integr. Pest Manag. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Amir Aghaee, M.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long term empirical and observational evidence of practical Helicoverpa zea resistance to cotton with pyramided Bt toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef]
- Huang, F.; Buschman, L.L.; Higgins, R.A.; McGaughey, W.H. Inheritance of resistance to Bacillus thuringiensis toxin (Dipel ES) in European corn borer. Science 1999, 284, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Alstad, D.; Andow, D.A. Managing the evolution of insect resistance to transgenic plants. Science 1995, 268, 1894–1896. [Google Scholar] [CrossRef] [PubMed]
- Anilkumar, K.J.; Pusztai-Carey, M.; Moar, W.J. Fitness costs associated with Cry1Ac-resistant Helicoverpa zea (Lepidoptera: Noctuidae): A factor countering selection for resistance to Bt cotton? J. Econ. Entomol. 2008, 101, 1421–1431. [Google Scholar] [CrossRef]
- Lin, S.; Head, P.G.; Niu, Y.; Huang, F. Relative fitness of susceptible and Cry1A.105/Cry2A2b2-single/dual-gene resistant Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) on non-Bt diet and a diet containing a low concentration of two Bt proteins. Ins. Sci. 2022. [Google Scholar] [CrossRef]
- Yang, F.; Head, G.P.; Price, P.A.; Santiago González, J.C.; Kerns, D.L. Inheritance of Bacillus thuringiensis Cry2Ab2 protein resistance in Helicoverpa zea (Lepidoptera: Noctuidae). Pest Manag. Sci. 2020, 76, 3676–3684. [Google Scholar] [CrossRef]
- Yang, F.; Santiago González, J.C.; Sword, G.A.; Kerns, D.L. Genetic basis of resistance to the Vip3Aa Bt protein in Helicoverpa zea. Pest Manag. Sci. 2021, 77, 1530–1535. [Google Scholar] [CrossRef]
- Benowitz, K.M.; Allan, C.W.; Degain, B.A.; Li, X.; Fabrick, J.A.; Tabashnik, B.E.; Carrière, Y.; Matzkin, L.M. Novel genetic basis of resistance to Bt toxin Cry1Ac in Helicoverpa zea. Genetics 2022, 221, iyac037. [Google Scholar] [CrossRef]
- Guo, J.; Oyediran, I.; Rice, M.E.; Brown, S.; Dimase, M.; Lin, S.; Walker, W.; Yu, W.; Niu, Y.; Huang, F. Seed blends of pyramided Cry/Vip maize reduce Helicoverpa zea populations from refuge ears. J. Pest Sci. 2021, 94, 959–968. [Google Scholar] [CrossRef]
- Niu, Y.; Oyediran, I.; Yu, W.; Lin, S.; Dimase, M.; Brown, S.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; Thrash, B.; et al. Populations of Helicoverpa zea (Boddie) in the southeastern United States are commonly resistant to Cry1Ab, but still susceptible to Vip3Aa20 expressed in MIR 162 corn. Toxins 2021, 13, 63. [Google Scholar] [CrossRef] [PubMed]
- Huang, F. Dominance and fitness costs of insect resistance to genetically modified Bacillus thuringiensis crops. GM Crops Food 2021, 12, 192–211. [Google Scholar] [CrossRef]
- Marçon, P.C.R.G.; Young, L.J.; Steffey, K.L.; Siegfried, B.D. Baseline susceptibility of European corn borer (Lepidoptera: Crambidae) to Bacillus thuringiensis toxins. J. Econ. Entomol. 1999, 92, 279–285. [Google Scholar] [CrossRef]
- Wu, X.; Leonard, B.R.; Zhu, Y.; Abel, C.A.; Head, G.P.; Huang, F. Susceptibility of Cry1Ab-resistant and -susceptible sugarcane borer (Lepidoptera: Crambidae) to four Bacillus thuringiensis toxins. J. Invert. Pathol. 2009, 100, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Leonard, B.R.; Gable, R.H. Comparative susceptibility of European corn borer, southwestern corn borer, and sugarcane borer (Lepidoptera: Crambidae) to Cry1Ab protein in a commercial Bt-corn hybrid. J. Econ. Entomol. 2006, 99, 194–202. [Google Scholar] [CrossRef]
- Abbott, W.S. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- SAS Institute. SAS/STAT: 9.3 User’s Third Edition; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Stone, B.F. A formula for determining degree of dominance in cases of monofactorial inheritance of resistance to chemicals. Bull. World Health Organ. 1968, 38, 325–326. [Google Scholar]
- Bourguet, D.; Genissel, A.; Raymond, M. Insecticide resistance and dominance levels. J. Econ. Entomol. 2000, 93, 1588–1595. [Google Scholar] [CrossRef]
- Lande, R. The minimum number of genes contributing to quantitative variation between and within populations. Genetics 1981, 99, 541–553. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Determining the mode of inheritance of pesticide resistance with backcross experiments. J. Econ. Entomol. 1991, 84, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac δ-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. 2005, 71, 948–954. [Google Scholar] [CrossRef]
- Liang, G.M.; Wu, K.M.; Yu, H.K.; Li, K.K.; Feng, X.; Guo, Y.Y. Changes of inheritance mode and fitness in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) along with its resistance evolution to Cry1Ac toxin. J. Invertebr. Pathol. 2007, 97, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Wei, Y.; Zhang, L.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Dominant resistance to Bt cotton and minor cross-resistance to Bt toxin Cry2Ab in cotton bollworm from China. Evol. Appl. 2013, 6, 1222–1235. [Google Scholar] [CrossRef] [PubMed]
- Kranthi, K.R.; Dhawad, C.S.; Naidu, S.R.; Mate, K.; Behere, G.T.; Wadaskar, R.M.; Kranthi, S. Inheritance of resistance in Indian Helicoverpa armigera (Hübner) to Cry1Ac toxin of Bacillus thuringiensis. Crop Prot. 2006, 25, 119–124. [Google Scholar] [CrossRef]
- Nair, R.; Kalia, V.; Aggarwal, K.K.; Gujar, G.T. Inheritance of Cry1Ac resistance and associated biological traits in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2010, 104, 31–38. [Google Scholar] [CrossRef]
- Kaur, P.; Dilawari, V.K. Inheritance of resistance to Bacillus thuringiensis Cry1Ac toxin in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) from India. Pest Manag. Sci. 2011, 67, 1294–1302. [Google Scholar] [CrossRef]
- Alvi, A.H.K.; Sayyed, A.H.; Naeem, M.; Ali, M. Field evolved resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) to Bacillus thuringiensis toxin Cry1Ac in Pakistan. PLoS ONE 2012, 7, e47309. [Google Scholar] [CrossRef]
- Akhurst, R.J.; James, W.; Bird, L.J.; Beard, C. Resistance to the Cry1Ac δ-endotoxin of Bacillus thuringiensis in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 1290–1299. [Google Scholar] [CrossRef]
- BCH (Biosafety Clearing-House). Cry1A.105. 2022. Available online: https://bch.cbd.int/en/database/43771 (accessed on 1 August 2022).
- Hernández-Rodríguez, C.S.; Hernández-Martínez, P.; Van Rie, J.; Escriche, B.; Ferré, J. Shared midgut binding sites for Cry1A.105, Cry1Aa, Cry1Ab, Cry1Ac and Cry1Fa proteins from Bacillus thuringiensis in two important corn pests, Ostrinia nubilalis and Spodoptera frugiperda. PLoS ONE 2013, 8, e68164. [Google Scholar] [CrossRef]


{kind=link}
| Insect Population | Source of Insect Population |
|---|---|
| Test I with the original Cry1A.105-RR population without backcrosses and reselection | |
| BZ-SS | A known Bt susceptible H. zea laboratory population provided by Benzon Research Inc (Carlisle, PA, USA). |
| Cry1A.105-RR | A single-toxin Cry1A.105-resistant H. zea population isolated from an F2 screen of isoline families collected in a maize field in 2019 in Louisiana. |
| Cry1A.105-F1a | A heterozygous Cry1A.105-resistant population formed by crossing Cry1A.105-RR♂ and BZ-SS♀. |
| Cry1A.105-F1b | A heterozygous Cry1A.105-resistant population formed by crossing Cry1A.105-RR♀ and BZ-SS♂. |
| Test II with a backcrossed-and-reselected Cry1A.105-resistant strain | |
| BZ-SS | A known Bt susceptible laboratory population provided by Benzon Research Inc (Carlisle, PA, USA). |
| Cry1A.105-RR’ | A backcrossed-and-reselected Cry1A.105-resistant H. zea population. |
| Cry1A.105-F1′a | A heterozygous Cry1A.105-resistant population formed by crossing Cry1A.105-RR’♂ and BZ-SS♀. |
| Cry1A.105-F1′b | A heterozygous Cry1A.105-resistant population formed by crossing Cry1A.105-RR’♀ and BZ-SS♂. |
| Cry1A.105-F2′ | A mixed F2 population formed by sib-mating within Cry1A.105-F1′a and Cry1A.105-F1′b |
| Cry1A.105-BC’ | A mixed population formed from reciprocal back-crosses between (Cry1A.105-F1′a + Cry1A.105-F1′b) and BZ-SS. |
| Insect Population | Number of Larvae Assayed | Slope ± SE | LC50(95%CL) (µg/cm2) | χ2 | p-Value | Resistance Ratio |
|---|---|---|---|---|---|---|
| Test-I | ||||||
| BZ-SS | 620 | 1.82 ± 0.20 | 0.008 (0.006, 0.010) | 14.75 | 0.6785 | - |
| Cry1A.105-RR | 640 | 1.50 ± 0.26 | 19.75 (13.12, 36.97) | 22.29 | 0.0729 | 2469 |
| Cry1A.105-F1a | 635 | 1.20 ± 0.11 | 2.49 (1.87, 3.35) | 26.75 | 0.2208 | 311 |
| Cry1A.105-F1b | 551 | 0.78 ± 0.12 | 1.63 (0.88, 2.93) | 36.69 | 0.0118 | 204 |
| Combined F1 | 1186 | 0.82 ± 0.10 | 1.73 (1.10, 2.55) | 63.35 | 0.0061 | 217 |
| Test-II | ||||||
| BZ-SS | 1408 | 2.75 ± 0.28 | 0.023 (0.019, 0.028) | 28.45 | 0.0124 | - |
| Cry1A.105-RR’ | 2285 | 0.65 ± 0.10 | 17.01 (10.75, 35.96) | 42.99 | 0.0586 | 740 |
| Cry1A.105-F1′a | 1214 | 0.94 ± 0.12 | 9.29 (6.16, 16.39) | 28.85 | 0.0503 | 404 |
| Cry1A.105-F1′b | 1215 | 1.04 ± 0.10 | 7.04 (5.39, 9.68) | 24.40 | 0.1423 | 306 |
| Combined F1 | 2429 | 0.99 ± 0.08 | 8.04 (6.24, 10.88) | 54.62 | 0.0394 | 350 |
| Cry1A.105-F2′ | 1204 | 0.56 ± 0.05 | 2.13 (1.45, 3.32) | 29.49 | 0.2892 | 93 |
| Cry1A.105-BC’ | 1088 | 0.37 ± 0.07 | 0.10 (0.02, 0.23) | 48.68 | 0.0045 | 4 |
| Resistant Population | Calculation Method | Dominance Level | Functional |
|---|---|---|---|
| Cry1A.105-RR | Stone’s D value | 0.376 | Incompletely dominant |
| DML at 1.00 µg/cm2 | 0.557 | Codominant | |
| DML at 3.16 µg/cm2 | 0.504 | Codominant | |
| DML at 10.0 µg/cm2 | 0.383 | Incompletely recessive | |
| DML at 31.6 µg/cm2 | 0.396 | Incompletely recessive | |
| Cry1A.105-RR’ | Stone’s D value | 0.773 | Incompletely dominant |
| DML at 1.00 µg/cm2 | 0.958 | Near completely dominant | |
| DML at 3.16 µg/cm2 | 0.947 | Near completely dominant | |
| DML at 10.0 µg/cm2 | 0.850 | Incompletely dominant | |
| DML at 31.6 µg/cm2 | 0.678 | Incompletely dominant |
| Insect Population | Bt Concentration (µg/cm2) | Number of Larvae Assayed | Observed Number of Dead Larvae | Expected Number of Dead Larvae | χ2 | p-Value |
|---|---|---|---|---|---|---|
| Cry1A.105-F2′ | 1.00 | 128 | 45 | 54.3 | 2.77 | 0.0961 |
| 3.16 | 125 | 67 | 60.4 | 1.40 | 0.2367 | |
| 10.0 | 119 | 71 | 77.3 | 1.47 | 0.2253 | |
| 31.6 | 64 | 54 | 46.2 | 4.73 | 0.0296 | |
| Cry1A.105-BC’ | 1.00 | 128 | 80 | 79.6 | 0.005 | 0.9436 |
| 3.16 | 128 | 82 | 84.7 | 0.25 | 0.6171 | |
| 10.0 | 128 | 101 | 99.8 | 0.07 | 0.7913 | |
| 31.6 | 64 | 55 | 53.8 | 0.17 | 0.6801 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, W.; Head, G.P.; Huang, F. Inheritance of Resistance to Cry1A.105 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae). Insects 2022, 13, 875. https://doi.org/10.3390/insects13100875
Yu W, Head GP, Huang F. Inheritance of Resistance to Cry1A.105 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae). Insects. 2022; 13(10):875. https://doi.org/10.3390/insects13100875
Chicago/Turabian StyleYu, Wenbo, Graham P. Head, and Fangneng Huang. 2022. "Inheritance of Resistance to Cry1A.105 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae)" Insects 13, no. 10: 875. https://doi.org/10.3390/insects13100875