
Anatomy of the Nervous System in Chelifer cancroides (Arachnida: Pseudoscorpiones) with a Distinct Sensory Pathway Associated with the Pedipalps


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection
2.2. Staining of Chelal Cuticle Using Congo Red
2.3. Vibratome Sections, Immunohistochemistry, and Phalloidin Labeling
2.4. Lipophilic Dye Labeling of Pedipalpal Nerves
2.5. Antibody Characterization
2.6. Microscopy and Image Acquisition
3. Results
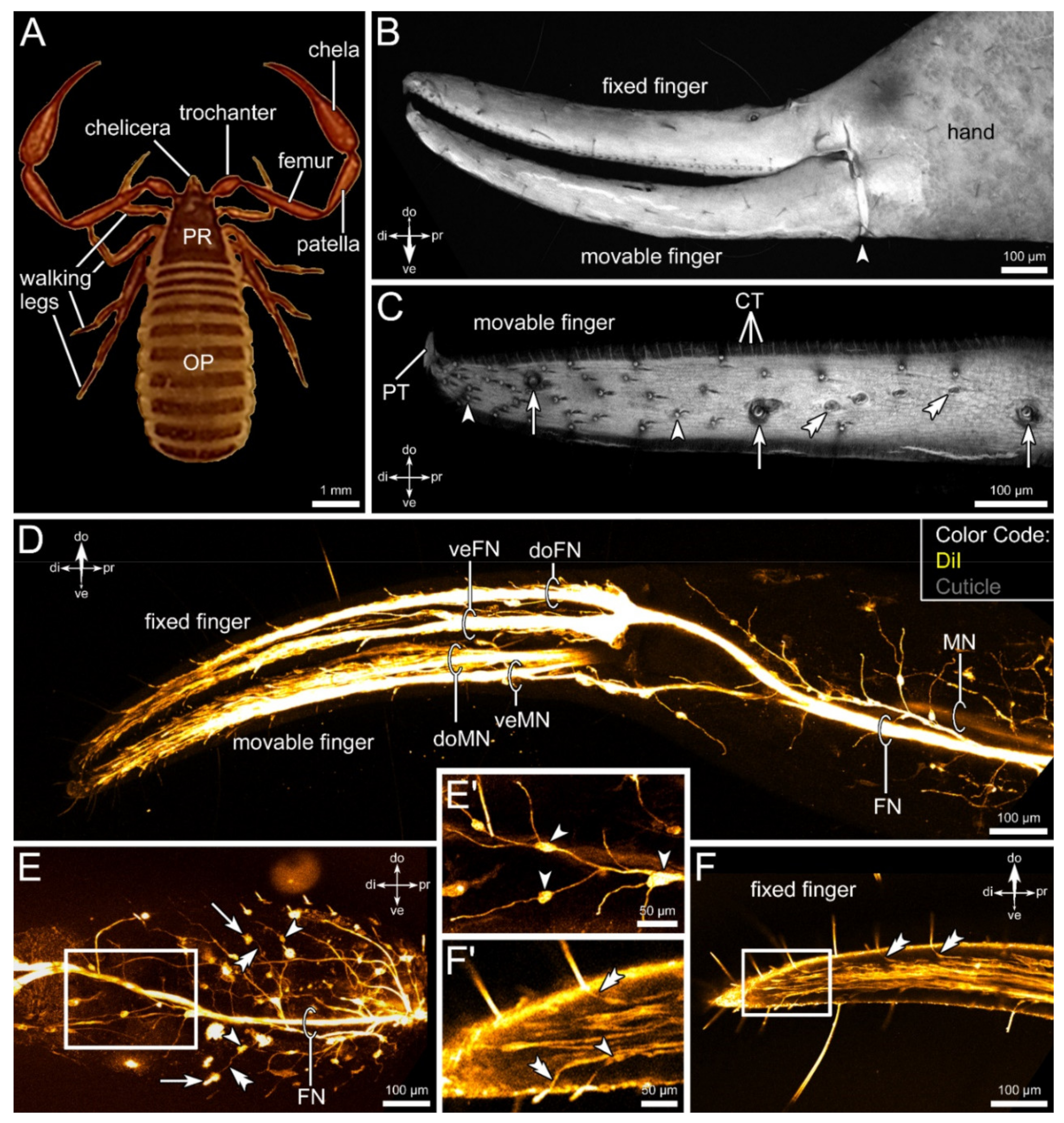
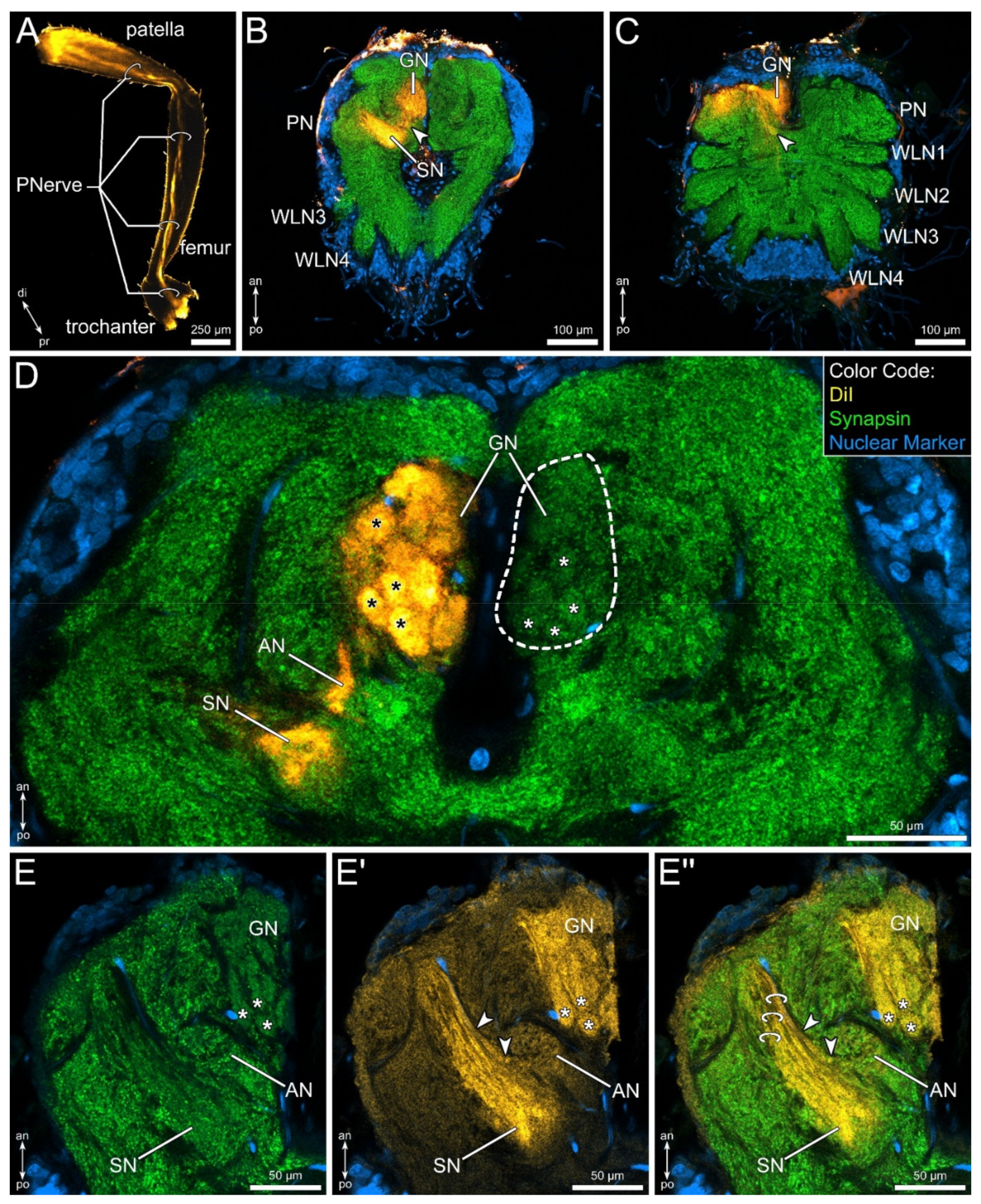
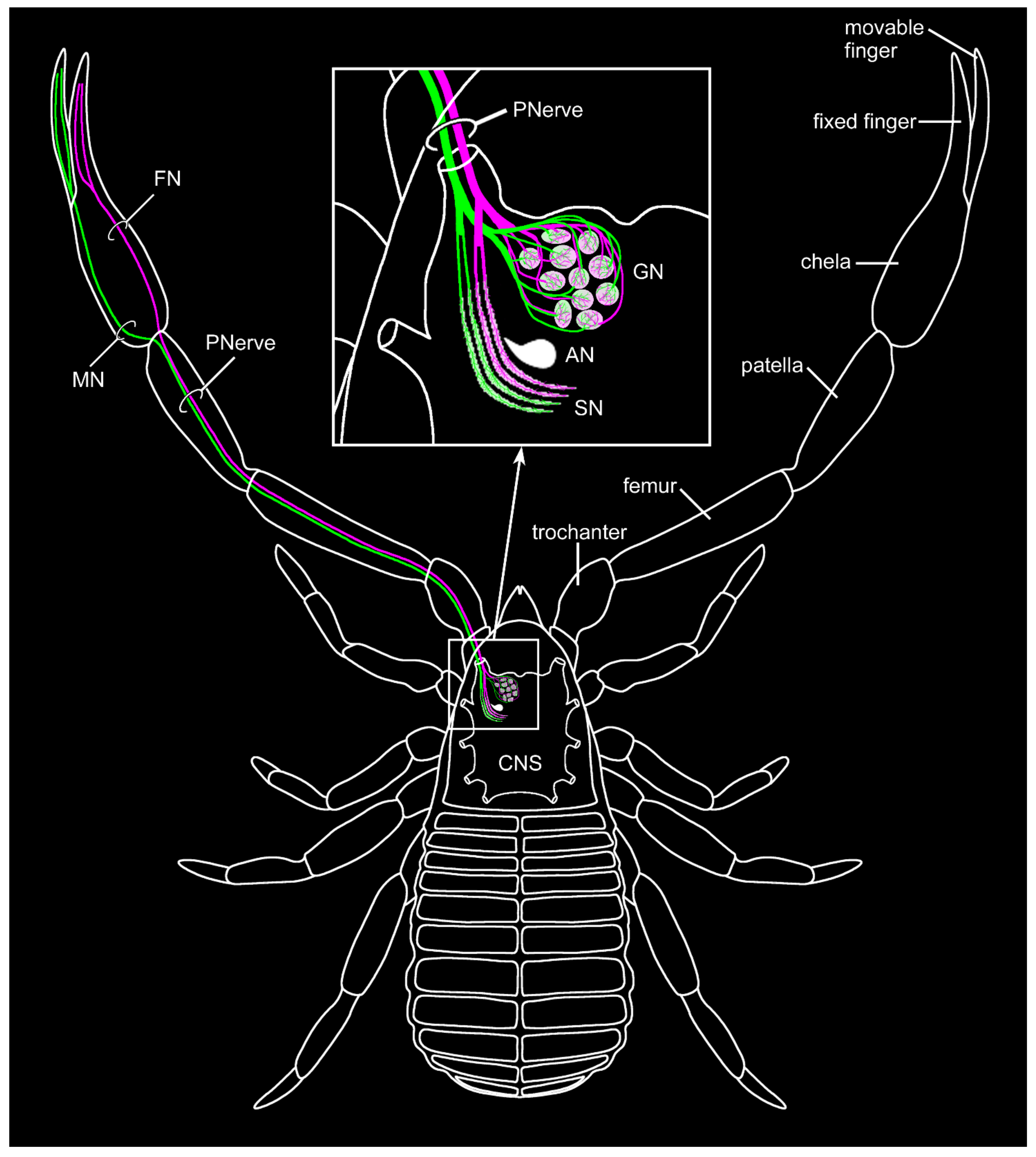
3.1. External Morphology and Innervation Pattern of the Pedipalps
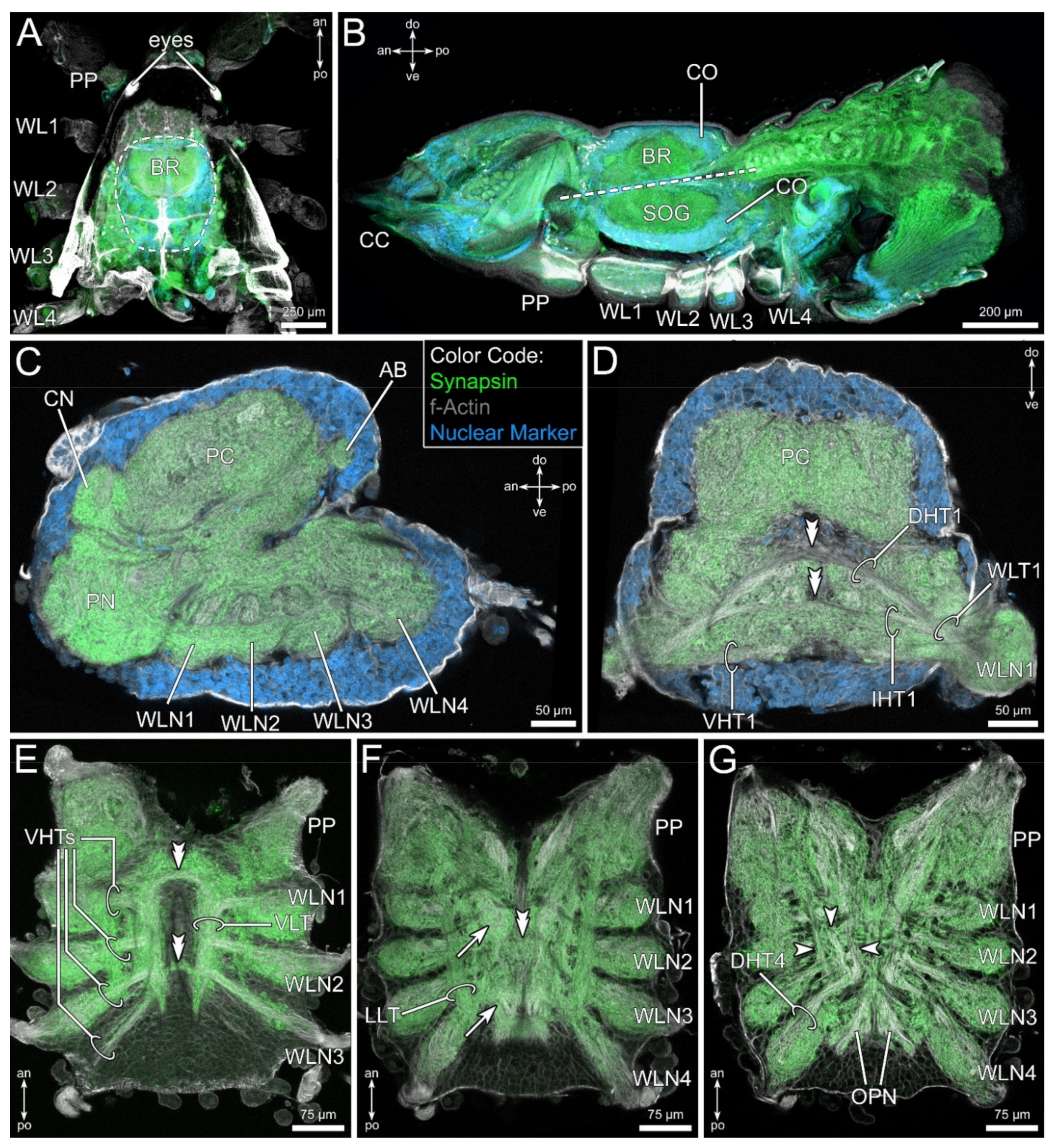
3.2. General Anatomy of the Central Nervous System
3.3. The Pedipalpal Neuropil Houses a Glomerular and a Stratified Neuropil
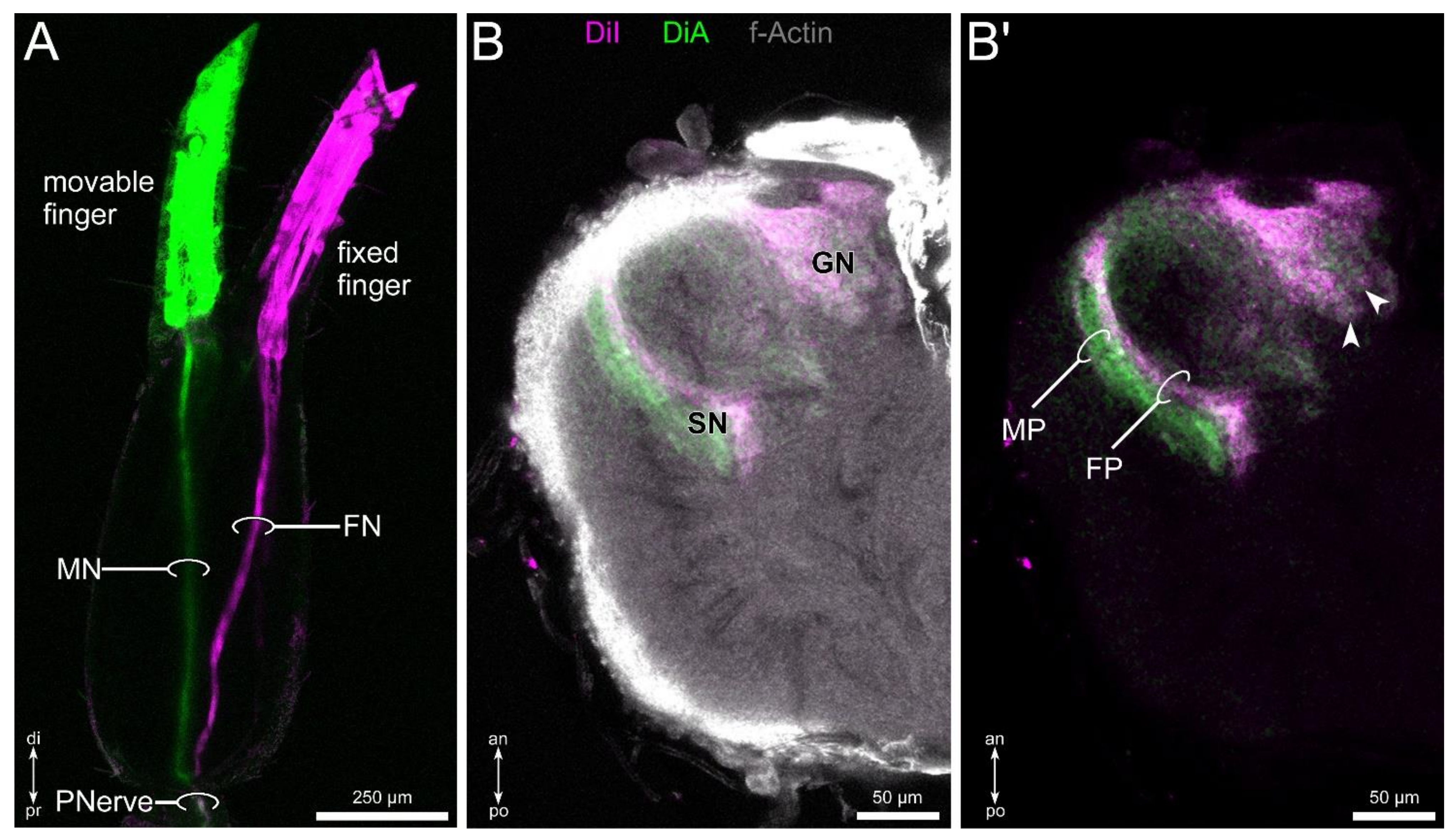
3.4. Lipophilic Dye Injection Reveal Connectivity of the Pedipalpal Nerve and Central Neuropilar Regions
3.5. Differential Innervation by Afferents from the Fixed and Movable Fingers
4. Discussion
4.1. Comparative Aspects of Prominent Protocerebral Neuropils
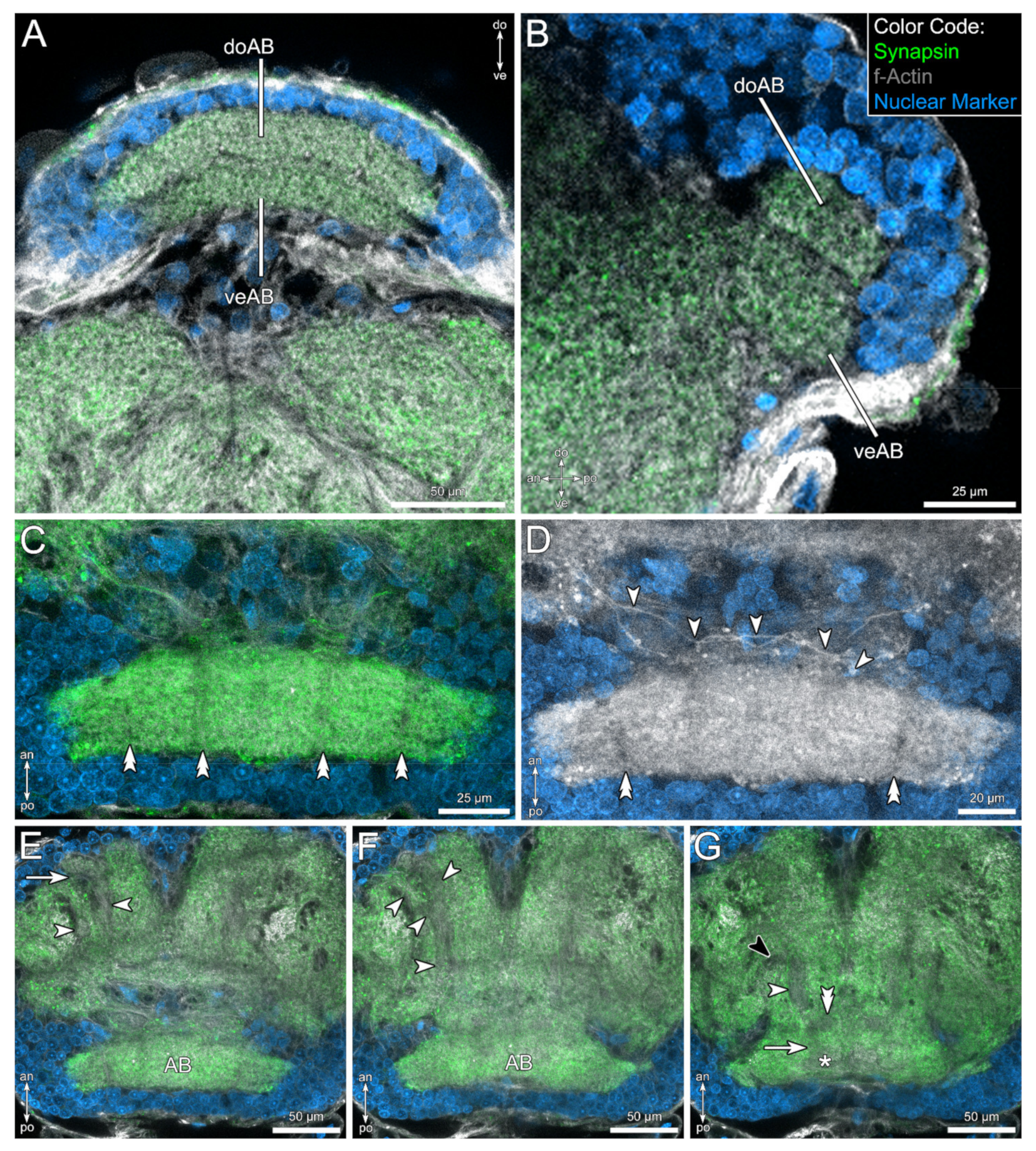
4.1.1. The Arcuate Body
4.1.2. The Mushroom Bodies
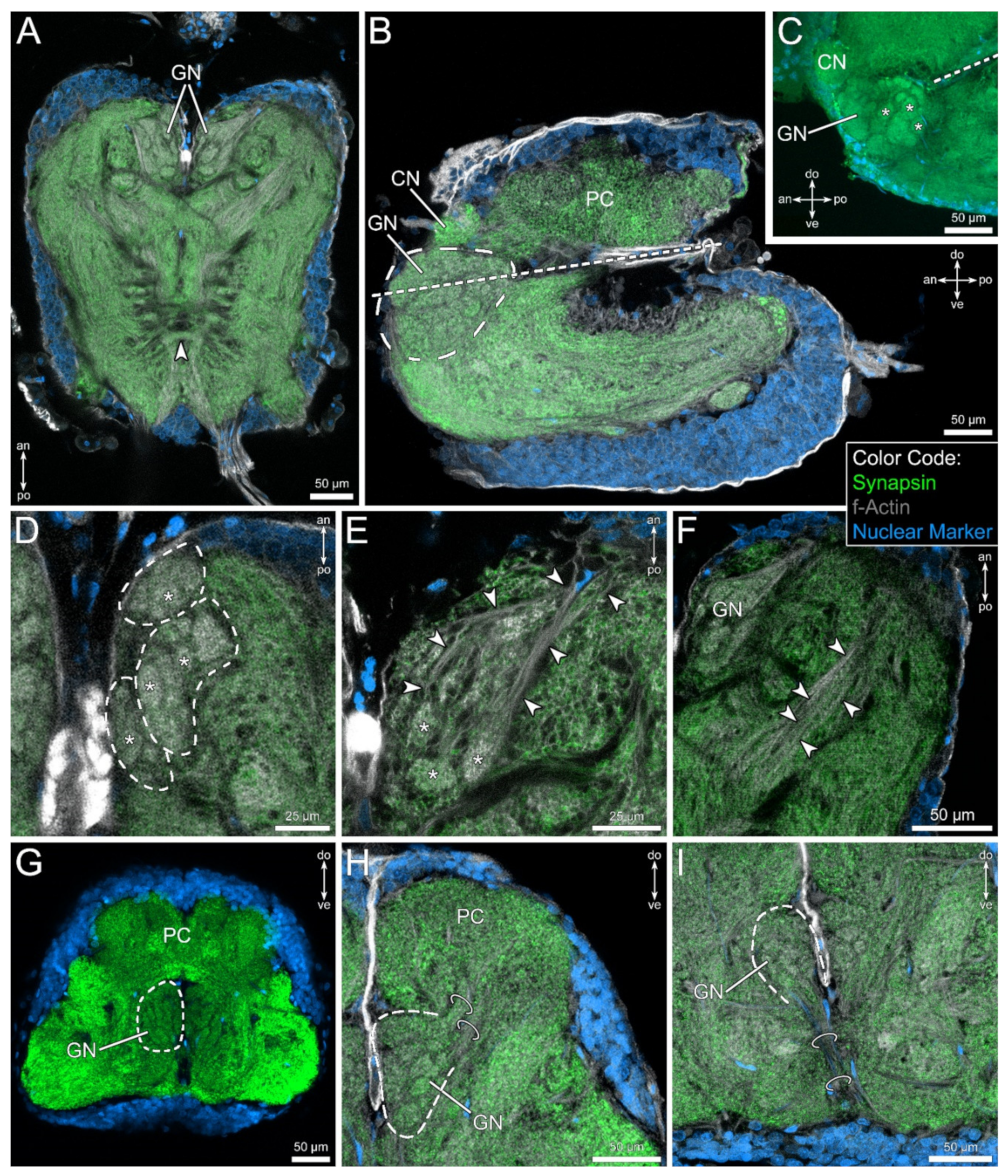
4.2. The Glomerular Neuropil Represents a First Integration Center for Chemosensory Cues
4.3. The Layered Neuropil Represents a First Integration Center for Mechanosensory Cues
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harvey, M.S. The smaller arachnid orders: Diversity, descriptions and distributions from Linnaeus to the present (1758 to 2007). Zootaxa 2007, 1668, 363–380. [Google Scholar] [CrossRef] [Green Version]
- Levi, H.W. Observation on two species of pseudoscorpions. Can. Entomol. 1953, 85, 55–62. [Google Scholar] [CrossRef]
- Weygoldt, P. Biology of Pseudoscorpions, 1st ed.; Harvard University Press: Harvard, UK, 1969; pp. 1–159. [Google Scholar]
- Brach, V. Social behavior in the pseudoscorpion Paratemnus elongatus (Banks) (Pseudoscorpionida, Atemnidae). Insect Soc. 1978, 25, 3–11. [Google Scholar] [CrossRef]
- Zeh, D.W. Life history consequences of sexual dimorphism in a chernetid pseudoscorpion. Ecology 1987, 68, 1495–1501. [Google Scholar] [CrossRef]
- Zeh, J.A.; Zeh, D.W. Cooperative Foraging for Large Prey by Paratemnus Elongatus (Pseudoscorpionida, Atemnidae). J. Arachnol. 1990, 18, 307–311. [Google Scholar]
- Weygoldt, P. Vergleichende Untersuchungen zur Fortpflanzungsbiologie der Pseudoscorpione. Beobachtungen über das Verhalten, die Samenübertragungsweisen und die Spermatophoren einiger einheimischer Arten. Zeitschrift Morphologie Ökologie Tiere 1966, 56, 39–92. [Google Scholar] [CrossRef]
- Beier, M. Phoresie und Phagophilie bei Pseudoscorpionen. Oesterreich. Zool. Z. 1948, 1, 441–497. [Google Scholar]
- Poinar, G.O.; Curcic, B.P.M.; Cokendolpher, J.C. Arthropod Phoresy Involving Pseudoscorpions in the Past and Present. Acta Arachnol. 1998, 47, 79–96. [Google Scholar] [CrossRef]
- Zeh, D.W.; Zeh, J.A. On the function of Harlequin Beetle-riding in the pseudoscorpion, Cordylochernes scorpioides (Pseudoscorpionida: Chernetidae). J. Arachnol. 1992, 20, 47–51. [Google Scholar]
- Zeh, D.W.; Zeh, J.A. Failed Predation or Transportation? Causes and Consequences of Phoretic Behavior in the Pseudoscorpion Dinocheirus arizonensis (Pseudoscorpionida: Chernetidae). J. Insect Behav. 1992, 5, 37–49. [Google Scholar] [CrossRef]
- Zeh, J.A.; Newcomer, S.D.; Zeh, D.W. Polyandrous females discriminate against previous mates. Proc. Natl. Acad. Sci. USA 1998, 95, 13732–13736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, M.M.; Zeh, D.W.; White, A.M.; Zeh, J.A. Discriminating Males and Unpredictable Females: Males Bias Sperm Allocation in Favor of Virgin Females. Ethology 2011, 117, 740–748. [Google Scholar] [CrossRef]
- Proctor, H.C. Mating biology resolves trichotomy for cheliferoid pseudoscorpions (Pseudoscorpionida, Cheliferoidea). J. Arachnol. 1993, 21, 156–158. [Google Scholar]
- Vachon, M. Recherches anatomiques et biologiques sur la reproduction et le développement des Pseudoscorpions. Ann. Sci. Nat. Zool. Biol. Anim. 1938, 11, 1–207. [Google Scholar]
- Harvey, M.S. A review and redescription of the cosmopolitan pseudoscorpion Chelifer cancroides (Pseudoscorpiones: Cheliferidae). J. Arachnol. 2014, 42, 86–104. [Google Scholar] [CrossRef] [Green Version]
- Weygoldt, P. Vergleichende Untersuchungen zur Fortpflanzungsbiologie der Pseudoscorpione II. J. Zool. Syst. Evol. Res. 1970, 8, 241–259. [Google Scholar] [CrossRef]
- Legg, G. Spermatophore formation in the pseudoscorpion Chthonius ischnocheles (Chthoniidae). J. Zool. 1973, 170, 367–394. [Google Scholar] [CrossRef]
- Legg, G. Taxonomy and the Dangers of Sex with special Reference to Pseudoscorpions. Adv. Arachnol. Dev. Biol. 2009, 14, 247–257. [Google Scholar]
- Judson, M.L.I. Roncocreagris murphyorum n. sp. and Occitanobisium nanum (Beier) n. comb. (Neobisiidae) from Iberia, with notes on the sternal glands of pseudoscorpions (Chelonethi). Bull. Br. Arachnol. Soc. 1992, 9, 26–30. [Google Scholar]
- Judson, M.L.I. A new and endangered pseudoscorpion of the genus Lagynochthonius (Arachnida, Chelonethi, Chthoniidae) from a cave in Vietnam, with notes on chelal morphology and the composition of the Tyrannochthoniini. Zootaxa 2007, 1627, 53–68. [Google Scholar] [CrossRef]
- Judson, M.L.I. Pseudoscorpions (Arachnida, Chelonethi) in Mexican amber, with a list of extant species associated with mangrove and Hymenaea trees in Chiapas. Bol. Soc. Geol. Mex. 2016, 68, 57–79. [Google Scholar] [CrossRef]
- Judson, M.L.I. A new subfamily of Feaellidae (Arachnida, Chelonethi, Feaelloidea) from Southeast Asia. Zootaxa 2017, 4258, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reboleira, A.S.P.S.; Zaragoza, J.A.; Goncalves, F.; Oromí, P. Titanobochica, surprising discovery of a new cave-dwelling genus from southern Portugal (Arachnida: Pseudoscorpiones: Bochicidae). Zootaxa 2010, 2681, 1–19. [Google Scholar] [CrossRef]
- Mahnert, V. A nature’s treasury: Pseudoscorpion diversity of the Canary Islands, with the description of nine new species (Pseudoscorpiones, Chthoniidae, Cheiridiidae) and new records. Revista Ibérica de Aracnologia 2011, 19, 27–45. [Google Scholar]
- Mahnert, V. Pseudoscorpions (Arachnida: Pseudoscorpiones) from the Galapagos Islands (Ecuador). Rev. Suisse Zool. 2014, 121, 135–210. [Google Scholar]
- Moro, S.D.; Geethabali. Distribution of cuticular sensory hairs on the legs and whip of Thelyphonus indicus stoliczka (Arachnida Uropygi). Ital. J. Zool. 1985, 19, 207–218. [Google Scholar]
- Igelmund, P. Morphology, sense organs, and regeneration of the forelegs (whips) of the whip spider Heterophrynus elaphus (Arachnida, Amblypygi). J. Morphol. 1987, 193, 75–89. [Google Scholar] [CrossRef]
- Schultz, J.W. Morphology of locomotor appendages in Arachnida: Evolutionary trends and phylogenetic implications. Zool. J. Linnean Soc. 1989, 97, 1–56. [Google Scholar] [CrossRef]
- Weygoldt, P. Whip Spiders. Their Biology, Morphology and Systematics (Chelicerata: Amblypygi), 1st ed.; Apollo Books: Stenstrup, Denmark, 2000; pp. 1–163. [Google Scholar]
- Carthy, J.D. Fine structure and function of the sensory pegs on the scorpion pecten. Experientia 1966, 22, 89–91. [Google Scholar] [CrossRef]
- Wolf, H. The pectine organs of the scorpion, Vaejovis spinigerus: Structure and (glomerular) central projections. Arthropod Struct. Dev. 2008, 37, 67–80. [Google Scholar] [CrossRef]
- Wolf, H. Scorpions pectines—Idiosyncratic chemo- and mechanosensory organs. Arthropod Struct. Dev. 2017, 46, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Brownell, P.H.; Farley, R.D. The organization of the malleolar sensory system in the solpugid, Chanbria sp. Tissue Cell 1974, 6, 471–485. [Google Scholar] [CrossRef]
- Lehmann, T.; Melzer, R.R.; Hörnig, M.K.; Michalik, P.; Sombke, A.; Harzsch, S. Arachnida (excluding Scorpiones). In Structure and Evolution of Invertebrate Nervous Systems, 1st ed.; Schmidt-Rhaesa, A., Harzsch, S., Purschke, G., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 453–477. [Google Scholar]
- Hilton, W.A. The nervous system of Chelifer. J. Entomol. Zool. 1913, 5, 93–95. [Google Scholar]
- Hilton, W.A. Nervous system and sense organs. Pseudoscorpionida. J. Entomol. Zool. 1931, 23, 67–75. [Google Scholar]
- Hanström, B. Zur Kenntnis des Zentralen Nervensystems der Arachnoiden und Pantopoden Nebst Schlussfolgerungen Betreffs der Phylogenie der Genannten Gruppen. Ph.D. Thesis, University of Lund, Lund, Sweden, 1919. [Google Scholar]
- Hanström, B. Vergleichende Anatomie des Nervensystems der Wirbellosen Tiere: Unter Berücksichtigung seiner Funktion, 1st ed.; Springer: Berlin, Germany, 1928; pp. 1–628. [Google Scholar]
- Strausfeld, N.J. Arthropod Brains; Evolution, Functional Elegance, and Historical Significance, 1st ed.; Belknap: Cambridge, UK, 2012; pp. 1–830. [Google Scholar]
- Wolff, G.H.; Strausfeld, N.J. The Insect Brain: A commentated primer. In Structure and Evolution of Invertebrate Nervous Systems, 1st ed.; Schmidt-Rhaesa, A., Harzsch, S., Purschke, G., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 597–639. [Google Scholar]
- Heisenberg, M. Mushroom body memoir: From maps to models. Nat. Rev. Neurosci. 2003, 4, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Strausfeld, N.J.; Sinakevitch, I.; Brown, S.M.; Farris, S.M. Ground plan of the insect mushroom body: Functional and evolutionary implications. J. Comp. Neurol. 2009, 513, 265–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, R.A.A.; Turner, G.C. The mushroom body. Curr. Biol. 2010, 20, R11–R12. [Google Scholar] [CrossRef] [Green Version]
- Aso, Y.; Hattori, D.; Yu, Y.; Johnston, R.M.; Iyer, N.A.; Ngo, T.T.B.; Dionne, H.; Abbott, L.F.; Axel, R.; Tanimoto, H.; et al. The neuronal architecture of the mushroom body provides a logic for associative learning. eLife 2014, 3, e04577. [Google Scholar] [CrossRef]
- Stopfer, M. Central processing in the mushroom bodies. Curr. Opin. Insect Sci. 2014, 6, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, T.; Melzer, R.R. A tiny visual systemdretinula axons and visual neuropils of Neobisium carcinoides (Hermann, 1804)(Chelicerata, Arachnida, Pseudoscorpiones). Zoologischer Anzeiger 2018, 273, 164–172. [Google Scholar] [CrossRef]
- Boissin, L.; Cazal, M. Étude du système nerveux et des glandes endocrines céphaliques de l’adulte femelle d’Hysterochelifer meridianus (L. Koch) (Arachnide, Pseudoscorpion, Cheliferidae). Bull. Soc. Zool. France 1969, 94, 263–268. [Google Scholar]
- Maurer, M.; Hladik, J.; Iliffe, T.M.; Stemme, T. Histaminergic interneurons in the ventral nerve cord: Assessment of their value for Euarthropod phylogeny. Zool. Lett. 2019, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Weygoldt, P. Vergleichend-embryologische Untersuchungen an Pseudoscorpionen (Chelonethi). Z. Morphol. Oekol. Tiere 1964, 54, 1–106. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Hildebrand, J.G. Olfactory systems: Common design, uncommon origins? Curr. Opin. Neurol. 1999, 9, 634–639. [Google Scholar] [CrossRef]
- Eisthen, H.L. Why Are Olfactory Systems of Different Animals So Similar? Brain Behav. Evol. 2002, 59, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Ache, B.W.; Young, J.M. Olfaction: Diverse Species, Conserved Principles. Neuron 2005, 48, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, K.S. Anatomy of the central nervous system of arachnids. Zool. Jb. Anat. 1965, 82, 1–35. [Google Scholar]
- Hill, D.E. The Structure of the Central Nervous System of Jumping Spiders of the Genus Phidippus (Araneae: Salticidae). Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1975. [Google Scholar]
- Babu, K.S.; Barth, F.G. Neuroanatomy of the central nervous system of the wandering spider, Cupiennius salei (Arachnida, Araneida). Zoomorphology 1984, 104, 344–359. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Barth, F.G. Two visual systems in one brain: Neuropils serving the secondary eyes of the spider Cupiennius salei. J. Comp. Neurol. 1993, 328, 43–62. [Google Scholar] [CrossRef]
- Sinakevitch, I.; Long, S.M.; Gronenberg, W. The central nervous system of whip spiders (Amblypygi): Large mushroom bodies receive olfactory and visual input. J. Comp. Neurol. 2021, 529, 1642–1658. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, P.O.M.; Uhl, G.; Harzsch, S.; Sombke, A. Visual pathways in the brain of the jumping spider Marpissa muscosa. J. Comp. Neurol. 2020, 528, 1883–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, S.M. Variations on a theme: Morphological variation in the secondary eye visual pathway across the order of Araneae. J. Comp. Neurol. 2021, 529, 259–280. [Google Scholar] [CrossRef] [PubMed]
- Schneeberg, K.; Bauernfeind, R.; Pohl, H. Comparison of cleaning methods for delicate insect specimens for scanning electron microscopy. Microsc. Res. Tech. 2017, 80, 1199–1204. [Google Scholar] [CrossRef]
- Michels, J.; Büntzow, M. Assessment of Congo red as a fluorescence marker for the exoskeleton of small crustaceans and the cuticle of polychaetes. J. Microsc. 2010, 238, 95–101. [Google Scholar] [CrossRef]
- Stemme, T.; Eickhoff, R.; Bicker, G. Olfactory projection neuron pathways in two species of marine Isopoda (Peracarida, Malacostraca, Crustacea). Tissue Cell 2014, 46, 260–263. [Google Scholar] [CrossRef]
- Harzsch, S.; Hansson, B.S. Brain architecture in the terrestrial hermit crab Coenobita clypeatus (Anomura, Coenobitidae), a crustacean with a good aerial sense of smell. BMC Neurosci. 2008, 9, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Groh, C.; Rössler, W. Comparison of microglomerular structures in the mushroom body calyx of neopteran insects. Arthropod Struct. Dev. 2011, 40, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Sombke, A.; Harzsch, S.; Hansson, B.S. Organization of deutocerebral neuropils and olfactory behavior in the centipede Scutigera coleoptrata (Linnaeus, 1758) (Myriapoda: Chilopoda). Chem. Senses 2011, 36, 43–61. [Google Scholar] [CrossRef] [Green Version]
- Stemme, T.; Iliffe, T.M.; von Reumont, B.M.; Koenemann, S.; Harzsch, S.; Bicker, G. Serotonin-immunoreactive neurons in the ventral nerve cord of Remipedia (Crustacea): Support for a sister group relationship of Remipedia and Hexapoda? BMC Evol. Biol. 2013, 13, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian-Fine, R.; Volknandt, W.; Seyfarth, E.A. Peripheral synapses at identifiable mechanosensory neurons in the spider Cupiennius salei: Synapsin-like immunoreactivity. Cell Tissue Res. 1999, 295, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, P.O.M.; Sombke, A.; Liedtke, J.; Schneider, J.M.; Harzsch, S.; Uhl, G. The synganglion of the jumping spider Marpissa muscosa (Arachnida: Salticidae): Insights from histology, immunohistochemistry and microCT analysis. Arthropod Struct. Dev. 2017, 46, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Drozd, D.; Wolf, H.; Stemme, T. Structure of the pecten neuropil pathway and its innervation by bimodal peg afferents in two scorpion species. PLoS ONE 2020, 15, e0243753. [Google Scholar] [CrossRef] [PubMed]
- Strausfeld, N.J.; Weltzien, P.; Barth, F.G. Two visual systems in one brain: Neuropils serving the principal eyes of the spider Cupiennius salei. J. Comp. Neurol. 1993, 328, 63–75. [Google Scholar] [CrossRef]
- Loesel, R.; Nässel, D.R.; Strausfeld, N.J. Common design in a unique midline neuropil in the brains of arthropods. Arthropod Struct. Dev. 2002, 31, 77–91. [Google Scholar] [CrossRef]
- Loesel, R.; Seyfarth, E.A.; Bräunig, P.; Agricola, H.J. Neuroarchitecture of the arcuate body in the brain of the spider Cupiennius salei (Araneae, Chelicerata) revealed by allatostatin-, proctolin-, and CCAP-immunocytochemistry and its evolutionary implications. Arthropod Struct. Dev. 2011, 40, 210–220. [Google Scholar] [CrossRef]
- Wolf, H. Scorpiones. In Structure and Evolution of Invertebrate Nervous Systems, 1st ed.; Schmidt-Rhaesa, A., Harzsch, S., Purschke, G., Eds.; Oxford University Press: Oxford, UK, 2019; pp. 443–452. [Google Scholar]
- Loesel, R.; Wolf, H.; Kenning, M.; Harzsch, S.; Sombke, A. Architectural principles and evolution of the arthropod central nervous system. In Arthropod Biology and Evolution, 1st ed.; Minelli, A., Boxshall, G., Fusco, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 299–342. [Google Scholar]
- Wolff, G.H.; Strausfeld, N.J. Genealogical correspondence of a forebrain centre implies an executive brain in the protostome-deuterostome bilaterian ancestor. Philos. Trans. R. Soc. 2016, 371, 20150055. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, F.C. The meaning and structure of the so-called “mushroom bodies” of the hexapod brain. Am. Nat. 1896, 30, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Richter, S.; Loesel, R.; Purschke, G.; Schmidt-Rhaesa, A.; Scholtz, G.; Stach, T.; Vogt, L.; Wanninger, A.; Brenneis, G.; Döring, C.; et al. Invertebrate neurophylogeny: Suggested terms and definitions for a neuroanatomical glossary. Front. Zool. 2010, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, T.; Melzer, R.R. Outsourcing a visual neuropil—The central visual system of the median eyes of Galeodes granti Pocock, 1903 (Arachnida: Solifugae). Arthropod Struct. Dev. 2021, 60, 101024. [Google Scholar] [CrossRef]
- Sombke, A.; Klann, A.E.; Lipke, E.; Wolf, H. Primary processing neuropils associated with the malleoli of camel spiders (Arachnida, Solifugae): A re-evaluation of axonal pathways. Zool. Lett. 2019, 5, 26. [Google Scholar] [CrossRef]
- Demoll, R. Die Sinnesorgane der Arthropoden ihr Bau und ihre Funktion, 1st ed.; Vieweg & Sohn: Braunschweig, Germany, 1917; pp. 1–243. [Google Scholar]
- Hansson, B.S.; Anton, S. Function and Morphology of the Antennal Lobe: New Developments. Annu. Rev. Entomol. 2000, 45, 203–231. [Google Scholar] [CrossRef] [Green Version]
- Schachtner, J.; Schmidt, M.; Homberg, U. Organization and evolutionary trends of primary olfactory brain centers in Tetraconata (Crustacea + Hexapoda). Arthropod Struct. Dev. 2005, 34, 257–299. [Google Scholar] [CrossRef]
- Schmidt, M.; Mellon, D. Neuronal processing of chemical information in crustaceans. In Chemical Communication in Crustaceans, 1st ed.; Breithaupt, T., Thiel, M., Eds.; Springer: New York, NY, USA, 2011; pp. 123–147. [Google Scholar]
- Stegner, M.E.J.; Richter, S. Morphology of the brain in Hutchinsoniella macracantha (Cephalocarida, Crustacea). Arthropod Struct. Dev. 2011, 40, 221–243. [Google Scholar] [CrossRef]
- Stemme, T.; Iliffe, T.M.; Bicker, G.; Harzsch, S.; Koenemann, S. Serotonin immunoreactive interneurons in the brain of the Remipedia: New insights into the phylogenetic affinities of an enigmatic crustacean taxon. BMC Evol. Biol. 2012, 12, 168. [Google Scholar] [CrossRef] [Green Version]
- Harzsch, S.; Krieger, J. Crustacean olfactory systems: A comparative review and a crustacean perspective on olfaction in insects. Prog. Neurobiol. 2018, 161, 23–60. [Google Scholar] [CrossRef] [PubMed]
- Sombke, A.; Lipke, E.; Kenning, M.; Müller, C.H.G.; Hansson, B.S. Comparative analysis of deutocerebral neuropils in Chilopoda (Myriapoda): Implications for the evolution of the arthropod olfactory system and support for the Mandibulata concept. BMC Neurosci. 2012, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownell, P.H. Glomerular Cytoarchitectures in chemosensory Systems of Arachnids. Ann. N. Y. Acad. Sci. 1998, 855, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Strausfeld, N.J.; Reisenman, C.E. Dimorphic Olfactory Lobes in the Arthropoda. Ann. N. Y. Acad. Sci. 2009, 1170, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Hummel, N.A.; Li, A.Y.; Witt, C.M. Serotonin-like immunoreactivity in the central nervous system of two ixodid tick species. Exp. Appl. Acarol. 2007, 43, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Menezes, K.M.F.; de Oliveira Filho, J.G.; Ferreira, L.L.; Borges, L.M.F. First neuronal projection from Haller’s organ to the synganglion and three-dimensional reconstruction of Amblyomma sculptum olfactory lobe. Tick Tick Borne Dis. 2021, 12, 101690. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B.S.; Christensen, T.A. Functional characteristics of the antennal lobe. In Insect Olfaction, 1st ed.; Hansson, B.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 125–161. [Google Scholar]
- Ignell, R.; Anton, S.; Hansson, B.S. The antennal lobe of Orthoptera–Anatomy and Evolution. Brain Behav. Evol. 2001, 57, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Polanska, M.A.; Tuchina, O.; Agricola, H.; Hansson, B.S.; Harzsch, S. Neuropeptide complexity in the crustacean central olfactory pathway: Immunolocalization of A-type allatostatins and RFamide-like peptides in the brain of a terrestrial hermit crab. Mol. Brain 2012, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Strausfeld, N.J. Crustacean–insect relationships: The use of brain characters to derive phylogeny amongst segmented invertebrates. Brain Behav. Evol. 1998, 52, 186–206. [Google Scholar] [CrossRef] [PubMed]
- Saint-Rémy, G. Contribution a l’étude du cerveau chez les arthropods trachéates. Arch. Zool. Exp. Gen. 1887, 2, 1–274. [Google Scholar]
- Fahlander, K. Beiträge zur Anatomie und systematischen Einteilung der Chilopoden. Zoologiska Bidrag från Uppsala 1938, 17, 1–148. [Google Scholar]
- Nguyen Duy-Jacquemin, M.N.; Arnold, G. Spatial organization of the antennal lobe in Cylindroiulus punctatus (Leach) (Myriapoda: Diplopoda). Int. J. Insect Morphol. Embryo. 1991, 20, 205–214. [Google Scholar] [CrossRef]
- Schmidt, M.; Van Ekeris, L.; Ache, B.W. Antennular projections to the midbrain of the spiny lobster. I. Sensory innervation of the lateral and medial antennular neuropils. J. Comp. Neurol. 1992, 318, 277–290. [Google Scholar] [CrossRef]
- Tuchina, O.; Koczan, S.; Harzsch, S.; Rybak, J.; Wolff, G.; Strausfeld, N.J.; Hansson, B.S. Central projections of antennular chemosensory and mechanosensory afferents in the brain of the terrestrial hermit crab (Coenobita clypeatus; Coenobitidae, Anomura). Front. Neuroanat. 2015, 9, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflüger, H.J.; Bräunig, P.; Hustert, R. The organization of mechanosensory neuropiles in locust thoracic ganglia. Phil. Trans. R. Soc. Lond. B. 1988, 321, 1–26. [Google Scholar] [CrossRef]
- Newland, P.L. Morphology and somatotopic organisation of the central projections of afferents from tactile hairs on the hind leg of the locust. J. Comp. Neurol. 1991, 312, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Newland, P.L.; Rogers, S.M.; Gaaboub, I.; Matheson, T. Parallel somatotopic maps of gustatory and mechanosensory neurons in the central nervous system of an insect. J. Comp. Neurol. 2000, 425, 82–96. [Google Scholar] [CrossRef]
- Babu, K.S.; Barth, F.G. Central nervous projections of mechanoreceptors in the spider Cupiennius salei Keys. Cell Tissue Res. 1989, 258, 69–82. [Google Scholar] [CrossRef]
- Anton, S.; Barth, F.G. Central nervous projection patterns of trichobothria and other cuticular sensilla in the wandering spider Cupiennius salei (Arachnida, Araneae). Zoomorphology 1993, 113, 21–32. [Google Scholar] [CrossRef]
- Gorb, S.N.; Anton, S.; Barth, F.G. Central projections of cheliceral mechanoreceptors in the spider Cupiennius salei (Arachnida, Araneae). J. Morphol. 1993, 217, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Babu, K.S.; Sreenivasulu, K.; Sekhar, V. Sensory projections of identified coxal hair sensilla of the scorpion Heterometrus fulvipes (Scorpionidae). J. Biosci. 1993, 18, 247–259. [Google Scholar] [CrossRef]
- Kirchmair, G.; Raspotnig, G. Mating behavior of Dactylochelifer latreillii latreillii (Pseudoscorpiones: Cheliferidae): A quantitative study. J. Arachnol. 2021, 49, 198–204. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stemme, T.; Pfeffer, S.E. Anatomy of the Nervous System in Chelifer cancroides (Arachnida: Pseudoscorpiones) with a Distinct Sensory Pathway Associated with the Pedipalps. Insects 2022, 13, 25. https://doi.org/10.3390/insects13010025
Stemme T, Pfeffer SE. Anatomy of the Nervous System in Chelifer cancroides (Arachnida: Pseudoscorpiones) with a Distinct Sensory Pathway Associated with the Pedipalps. Insects. 2022; 13(1):25. https://doi.org/10.3390/insects13010025
Chicago/Turabian StyleStemme, Torben, and Sarah E. Pfeffer. 2022. "Anatomy of the Nervous System in Chelifer cancroides (Arachnida: Pseudoscorpiones) with a Distinct Sensory Pathway Associated with the Pedipalps" Insects 13, no. 1: 25. https://doi.org/10.3390/insects13010025