
Life Table and Preference Choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for Kidney Bean Plants Treated by Exogenous Calcium

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Plant
2.3. Life Table Study
2.3.1. Thrips Development Duration and Nymphal Survival
2.3.2. Thrips Longevity and Fecundity
2.4. Choice Test
2.5. Data Analysis
2.5.1. Life Table Analyses
2.5.2. Choice Tests
3. Results
3.1. Developmental and Reproductive Parameters
3.1.1. Development Duration
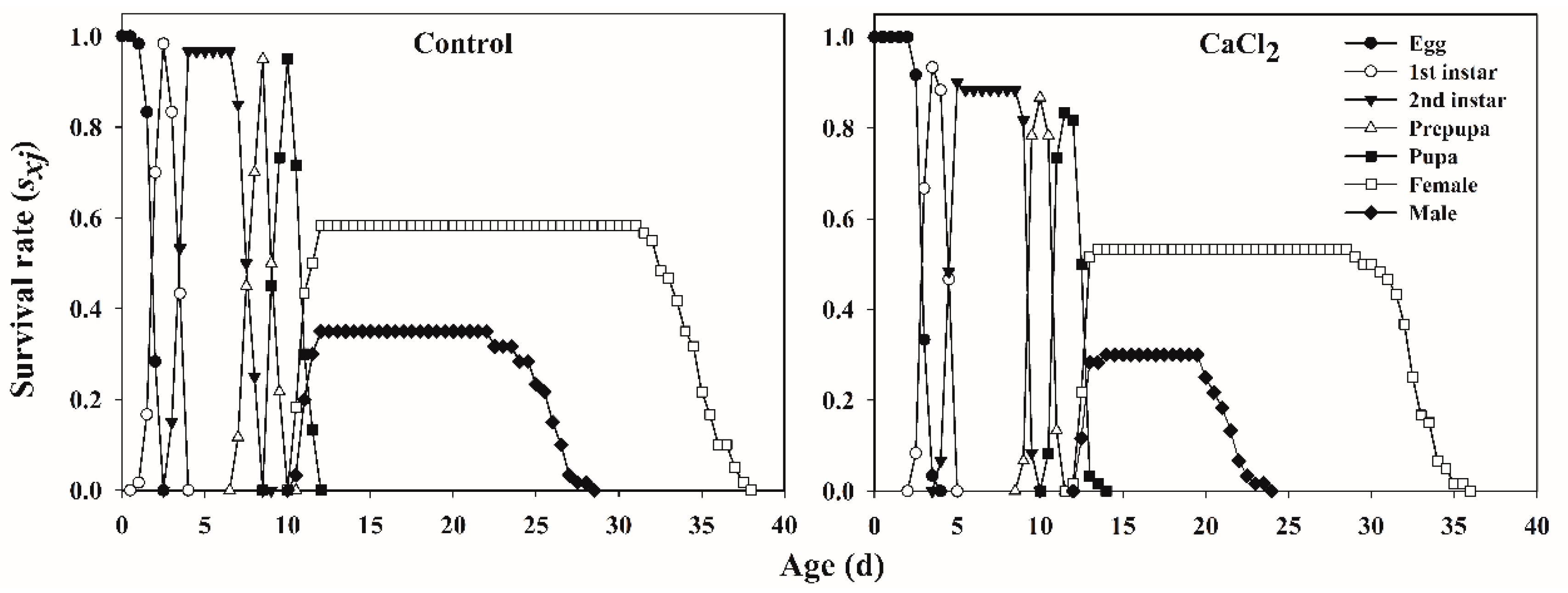
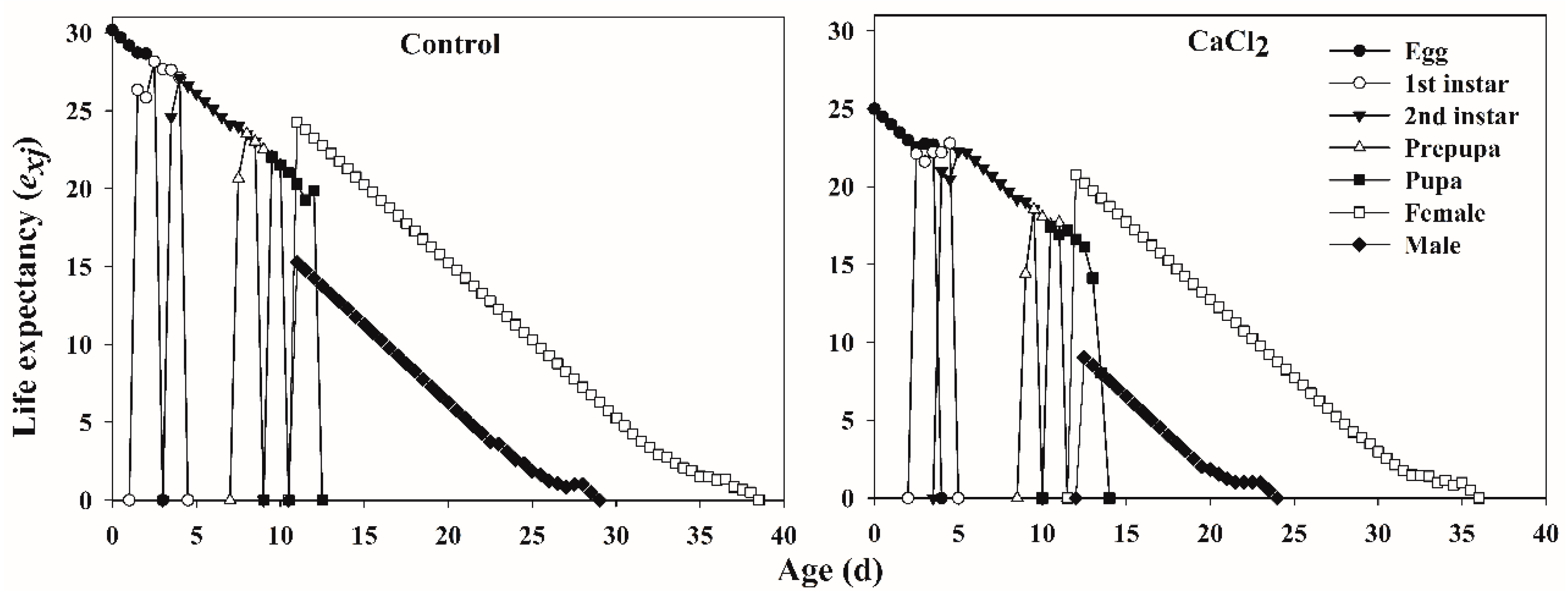
3.1.2. Age-Specific Survivorship, Life Expectancy, and Fecundity
3.1.3. Population Parameters
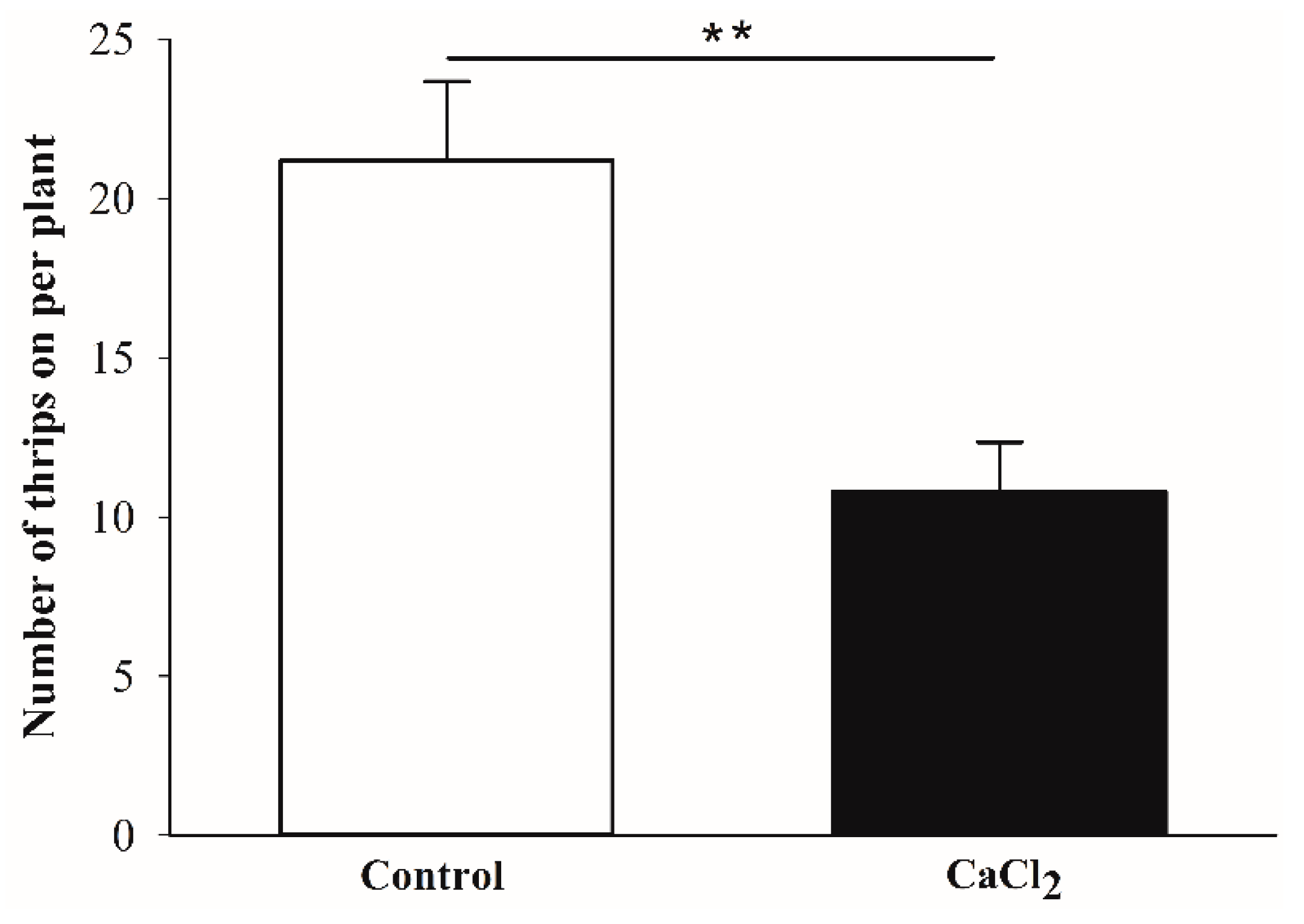
3.2. Choice Test
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agric. For. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.L.; Kirk, W.D.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Shimoda, T.; Ohnishi, J.; Kugimiya, S.; Narusaka, M.; Seo, S.; Narusaka, Y.; Tsuda, S.; Kobayashi, M. Jasmonate-dependent plant defense restricts thrips performance and preference. BMC Plant Biol. 2009, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R.; Diffie, S. Thrips vectors of tospoviruses. J. Integr. Pest Manag. 2011, 2, I1–I10. [Google Scholar] [CrossRef]
- Wu, S.Y.; Tang, L.D.; Zhang, X.R.; Xing, Z.L.; Lei, Z.R.; Gao, Y.L. A decade of a thrips invasion in China: Lessons learned. Ecotoxicology 2017, 27, 1032–1038. [Google Scholar] [CrossRef]
- Gao, Y.L.; Lei, Z.R.; Reitz, S.R. Western flower thrips resistance to insecticides: Detection, mechanisms and management strategies. Pest Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Mouden, S.; Sarmiento, K.F.; Klinkhamer, P.G.L.; Leiss, K.A. Integrated pest management in western flower thrips: Past, present and future. Pest Manag. Sci. 2017, 73, 813–822. [Google Scholar] [CrossRef]
- Steenbergen, M.; Abd-El-Haliem, A.; Bleeker, P.; Dicke, M.; Escobar-Bravo, R.; Cheng, G.; Leiss, K.A. Thrips advisor: Exploiting thrips-induced defences to combat pests on crops. J. Exp. Bot. 2018, 69, 1837–1848. [Google Scholar] [CrossRef]
- Mouden, S.; Leiss, K.A. Host plant resistance to thrips (Thysanoptera: Thripidae)—current state of art and future research avenues. Curr. Opin. Insect Sci. 2020, 45, 28–34. [Google Scholar] [CrossRef]
- Chen, G.; Escobar-Bravo, R.; Kim, H.K.; Leiss, K.A.; Klinkhamer, P.G.L. Induced resistance against western flower thrips by the Pseudomonas syringae-derived defense elicitors in tomato. Front. Plant Sci. 2018, 9, 1417. [Google Scholar] [CrossRef]
- Bittner, N.; Hundacker, J.; Achotegui-Castells, A.; Anderbrant, O.; Hilker, M. Defense of Scots pine against sawfly eggs (Diprion pini) is primed by exposure to sawfly sex pheromones. Proc. Natl. Acad. Sci. USA 2019, 116, 24668–24675. [Google Scholar] [CrossRef] [Green Version]
- Jeer, M.; Telugu, U.M.; Voleti, S.R.; Padmakumari, A.P. Soil application of silicon reduces yellow stem borer, Scirpophaga incertulas (Walker) damage in rice. J. Appl. Entomol. 2017, 141, 189–201. [Google Scholar] [CrossRef]
- Ye, M.; Song, Y.; Long, J.; Wang, R.; Baerson, S.R.; Pan, Z.Q.; Zhu-Salzman, K.; Xie, J.F.; Cai, K.; Luo, S.M.; et al. Priming of jasmonate-mediated antiherbivore defense responses in rice by silicon. Proc. Natl. Acad. Sci. USA 2013, 110, E3631–E3639. [Google Scholar] [CrossRef] [Green Version]
- Venter, E.; Mansoor, C.V.; Sibisi, P.; Botha, A.M. Potassium phosphate induces tolerance against the Russian wheat aphid (Diuraphis noxia; Homoptera: Aphididae) in wheat. Crop. Prot. 2014, 61, 43–50. [Google Scholar] [CrossRef]
- Jeer, M.; Yele, Y.; Sharma, K.C.; Prakash, N.B. Exogenous application of different silicon sources and potassium reduces pink stem borer damage and improves photosynthesis; yield and related parameters in wheat. Silicon 2021, 13, 901–910. [Google Scholar] [CrossRef]
- Chen, Y.; Li, R.; Li, B.P.; Meng, L. Biochar applications decrease reproductive potential of the English grain aphid Sitobion avenae and upregulate defense-related gene expression. Pest Manag. Sci. 2019, 75, 1310–1316. [Google Scholar] [CrossRef]
- Larkindale, J.; Knight, M.R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiol. 2002, 128, 682–695. [Google Scholar] [CrossRef]
- Erinle, K.O.; Jiang, Z.; Ma, B.; Li, J.M.; Chen, Y.K.; Ur-Rehman, K.; Shahla, A.; Zhang, Y. Exogenous calcium induces tolerance to atrazine stress in Pennisetum seedlings and promotes photosynthetic activity; antioxidant enzymes and psbA gene transcripts. Ecotoxicol. Environ. Saf. 2016, 132, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Mostofa, M.G.; Nahar, K.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous calcium alleviates cadmium-induced oxidative stress in rice (Oryza sativa L.) seedlings by regulating the antioxidant defense and glyoxalase systems. Braz. J. Bot. 2016, 39, 393–407. [Google Scholar] [CrossRef]
- Roy, P.R.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Hossen, M.Z.; Hossain, M.A. Physiological mechanisms of exogenous calcium on alleviating salinity-induced stress in rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2019, 25, 611–624. [Google Scholar] [CrossRef]
- Debona, D.; Cruz, M.F.A.; Rodrigues, F.A. Calcium-triggered accumulation of defense-related transcripts enhances wheat resistance to leaf blast. Trop. Plant Pathol. 2017, 42, 309–314. [Google Scholar] [CrossRef]
- Yamazaki, H.; Kikuchi, S.; Hoshina, T.; Kimura, T. Effect of calcium concentration in nutrient solution on development of bacterial wilt and population of its pathogen Ralstonia solanacearum in grafted tomato seedlings. Soil Sci. Plant Nutr. 2000, 46, 535–539. [Google Scholar]
- Ehret, D.L.; Utkhede, R.S.; Frey, B.; Menzies, J.G.; Bogdanoff, C. Foliar applications of fertilizer salts inhibit powdery mildew on tomato. Can. J. Plant Pathol. 2002, 24, 437–444. [Google Scholar] [CrossRef]
- Keeping, M.G.; Meyer, J.H. Calcium silicate enhances resistance of sugarcane to the African stalk borer Eldana saccharina Walker (Lepidoptera: Pyralidae). Agric. For. Entomol. 2002, 4, 265–274. [Google Scholar] [CrossRef]
- Wang, J.; Song, J.; Wu, X.B.; Deng, Q.Q.; Zhu, Z.Y.; Ren, M.J.; Ye, M.; Zeng, R.S. Seed priming with calcium chloride enhances wheat resistance against wheat aphid Schizaphis graminum Rondani. Pest Manag. Sci. 2021, 77, 4709–4718. [Google Scholar] [CrossRef]
- Zeng, G.; Zhi, J.R.; Ye, M.; Yue, W.B.; Song, J. Inductive effects of exogenous calcium on the defense of kidney bean plants against Frankliniella occidentalis (Thysanoptera: Thripidae). Arthropod-Plant Interact. 2020, 14, 473–480. [Google Scholar] [CrossRef]
- van Rijn, P.C.J.; Mollema, C.; Steenhuis-Broers, G.M. Comparative life history studies of Frankliniella occidentalis and Thrip tabaci (Thysanoptera: Thripidae) on cucumber. Bull. Entomol. Res. 1995, 85, 285–297. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 18–25. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-Stage, Two-Sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Ding, T.; Chi, H.; Gökçe, A.; Gao, Y.L.; Zhang, B. Demographic analysis of arrhenotokous parthenogenesis and bisexual reproduction of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Atrchian, H.; Mahdian, K.; Shahidi, S. Effect of different natural and factitious diets on development and life history parameters of Chilocorus bipustulatus. BioControl 2016, 61, 703–715. [Google Scholar] [CrossRef]
- Arshad, M.; Ullah, M.I.; Khan, R.R.; Anjum, S.; Tahir, M.; Shamshad, A.; Rizwan, M. Demographic parameters of the reduviid predator, Rhynocoris marginatus (Reduviidae: Hemiptera) fed on two lepidopterous insect pests. BioControl 2021, 66, 227–235. [Google Scholar] [CrossRef]
- Ou, H.D.; Atlihan, R.; Wang, X.Q.; Li, H.X.; Yu, X.F.; Jin, X.; Yang, M.F. Host deprivation effects on population performance and paralysis rates of Habrobracon hebetor (Hymenoptera: Braconidae). Pest Manag. Sci. 2021, 77, 1851–1863. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2020; Available online: http://140.120.197.173/Ecology/ (accessed on 20 August 2020).
- Sandeep, J.S.; Klaas, B.; Jhon, V.M.; Anja, D.; Wilhelm, B.; Marcel, D. Involvement of sweet pepper CaLOX2 in jasmonate-dependent induced defence against western flower thrips. J. Integr. Plant Biol. 2019, 61, 1085–1098. [Google Scholar]
- Koschier, E.H.; Hoffmann, D.; Riefler, J. Influence of salicylaldehyde and methyl salicylate on post-landing behaviour of Frankliniella occidentalis Pergande. J. Appl. Entomol. 2007, 131, 362–367. [Google Scholar] [CrossRef]
- Escobar-Bravo, R.; Chen, G.; Kim, H.K.; Grosser, K.; van Dam, N.M.; Leiss, K.A.; Klinkhamer, P.G. Ultraviolet radiation exposure time and intensity modulate tomato resistance to herbivory through activation of jasmonic acid signaling. J. Exp. Bot. 2019, 70, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Bravo, R.; Ruijgrok, J.; Kim, H.K.; Grosser, K.; Van Dam, N.M.; Klinkhamer, P.G.; Leiss, K.A. Light intensity-mediated induction of trichome-associated allelochemicals increases resistance against thrips in tomato. Plant Cell Physiol. 2018, 59, 2462–2475. [Google Scholar] [CrossRef] [Green Version]
- Brodbeck, B.V.; Stavisky, J.; Funderburk, J.E.; Andersen, P.C.; Olson, S.M. Flower nitrogen status and populations of Frankliniella occidentalis feeding on Lycopersicon esculentum. Entomol. Exp. Appl. 2001, 99, 165–172. [Google Scholar] [CrossRef]
- Qian, L.; He, S.Q.; Liu, X.W.; Huang, Z.J.; Chen, F.J.; Gui, F.R. Effect of elevated CO2 on the interaction between invasive thrips, Frankliniella occidentalis, and its host kidney bean, Phaseolus vulgaris. Pest Manag. Sci. 2018, 74, 2773–2782. [Google Scholar] [CrossRef]
- Peneder, S.; Koschier, E.H. Toxic and behavioural effects of carvacrol and thymol on Frankliniella occidentalis larvae. J. Plant Dis. Prot. 2011, 118, 26–30. [Google Scholar] [CrossRef]
- Murata, M.; Kobayashi, T.; Seo, S. α-ionone, an apocarotenoid, induces plant resistance to western flower thrips, Frankliniella occidentalis, independently of jasmonic acid. Molecules 2020, 25, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outchkourov, N.S.; De Kogel, W.J.; Wiegers, G.L.; Abrahamson, M.; Jongsma, M.A. Engineered multidomain cysteine protease inhibitors yield resistance against western flower thrips (Frankliniella occidentalis) in greenhouse trials. Plant Biotechnol. J. 2004, 2, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, T.; Watanabe, K.; Yoshida, S.; Aino, M.; Furiki, M.; Shiono, M.; Matoh, T.; Biggs, A.R. Field application of calcium to reduce Phytophthora stem rot of soybean, and calcium distribution in plants. Plant Dis. 2010, 94, 812–819. [Google Scholar] [CrossRef] [Green Version]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar]
- Heitz, T.; Ryan, B.C.A. A gene encoding a chloroplast-targeted lipoxygenase in tomato leaves is transiently induced by wounding, systemin, and methyl jasmonate. Plant Physiol. 1997, 114, 1085–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Hamilton, W. Jasmonate-induced responses of Nicotiana sylvestris results in fitness costs due to impaired competitive ability for nitrogen. J. Chem. Ecol. 2000, 26, 915–952. [Google Scholar] [CrossRef]
- Shoji, T.; Yamada, Y.; Hashimoto, T. Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol. 2000, 41, 831–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, K.L.; Kästner, J.; Bodenhausen, N.; Schramm, K.; Paetz, C.; Vassão, D.G.; Reichelt, M.; von Knorre, D.; Bergelson, J.; Erb, M.; et al. The role of glucosinolates and the jasmonic acid pathway in resistance of Arabidopsis thaliana against molluscan herbivores. Mol. Ecol. 2014, 23, 1188–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar-Bravo, R.; Alba, J.M.; Pons, C.; Granell, A.; Kant, M.R.; Moriones, E.; Fernández-Muñoz, R. A jasmonate-inducible defense trait transferred from wild into cultivated tomato establishes increased whitefly resistance and reduced viral disease incidence. Front. Plant Sci. 2016, 7, 1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.J.; Zhang, Y.J.; Xu, B.Y.; Zhu, G.R. The biological character, damege and management of an invasive insect pest, Frankliniella occidentalis. Chin. J. Appl. Entomol. 2005, 42, 11–14. [Google Scholar]
- Hopkins, R.J.; van Dam, N.M.; van Loon, J.J. Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.W.; Gratton, C.; Wolkowski, R.P.; Hogg, D.B.; Wedberg, J.L. Effect of soil potassium availability on soybean aphid (Hemiptera: Aphididae) population dynamics and soybean yield. J. Econ. Entomol. 2005, 98, 113–120. [Google Scholar] [CrossRef]
- Tripathy, S.; Rath, L.K. Non-preference mechanism of induced resistance in rice to white backed plant hopper through application of zinc. J. Entomol. Zool. Stud. 2019, 7, 1240–1243. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | n | Control | n | Ca-Treated |
|---|---|---|---|---|
| Egg (d) | 60 | 2.55 ± 0.05 b | 60 | 3.14 ± 0.04 a |
| First instar (d) | 59 | 1.57 ± 0.03 a | 57 | 1.58 ± 0.03 a |
| Second instar (d) | 58 | 4.17 ± 0.05 b | 54 | 4.79 ± 0.05 a |
| Prepupa (d) | 57 | 1.51 ± 0.03 a | 52 | 1.53 ± 0.04 a |
| Pupa (d) | 56 | 1.75 ± 0.04 a | 50 | 1.79 ± 0.05 a |
| Immature stages (d) | 56 | 11.61 ± 0.06 b | 50 | 12.82 ± 0.05 a |
| Preadult survival rate | 56 | 0.93 ± 0.03 a | 50 | 0.83 ± 0.05 a |
| Longevity/Female (d) | 35 | 24.72 ± 0.26 a | 32 | 19.94 ± 0.25 b |
| Longevity/Male (d) | 21 | 14.52 ± 0.33 a | 18 | 8.67 ± 0.28 b |
| APOP (d) | 56 | 1.97 ± 0.09 a | 50 | 2.0 ± 0. 0.13 a |
| TPOP (d) | 56 | 13.51 ± 0.13 b | 50 | 14.79 ± 0.14 a |
| Fecundity (eggs/female) | 35 | 80.29 ± 7.29 a | 32 | 55.34 ± 8.09 b |
| Oviposition days (d) | 35 | 19.88 ± 0.14 a | 32 | 16.14 ± 0.13 b |
| Treatments | r (d−1) | λ (d−1) | R0 (Offspring) | T (d−1) |
|---|---|---|---|---|
| Control | 0.19 ± 0.01 a | 1.21 ± 0.01 a | 47.17 ± 5.20 a | 19.93 ± 0.15 b |
| Ca-treated | 0.16 ± 0.01 b | 1.17 ± 0.01 b | 29.62 ± 3.66 b | 21.04 ± 0.13 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, G.; Zhi, J.-R.; Ye, M.; Xie, W.; Zhang, T.; Li, D.-Y.; Liu, L.; Wu, X.-B.; Cao, Y. Life Table and Preference Choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for Kidney Bean Plants Treated by Exogenous Calcium. Insects 2021, 12, 838. https://doi.org/10.3390/insects12090838
Zeng G, Zhi J-R, Ye M, Xie W, Zhang T, Li D-Y, Liu L, Wu X-B, Cao Y. Life Table and Preference Choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for Kidney Bean Plants Treated by Exogenous Calcium. Insects. 2021; 12(9):838. https://doi.org/10.3390/insects12090838
Chicago/Turabian StyleZeng, Guang, Jun-Rui Zhi, Mao Ye, Wen Xie, Tao Zhang, Ding-Ying Li, Li Liu, Xiao-Bao Wu, and Yu Cao. 2021. "Life Table and Preference Choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for Kidney Bean Plants Treated by Exogenous Calcium" Insects 12, no. 9: 838. https://doi.org/10.3390/insects12090838