
Laboratory Selection and Assessment of Resistance Risk in Drosophila suzukii (Diptera: Drosophilidae) to Spinosad and Malathion


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Selection of Drosophila suzukii Resistance to Spinosad and Malathion
2.3. Dose–Response Bioassay
2.4. Data Analysis
2.5. Estimation of Realized Heritability
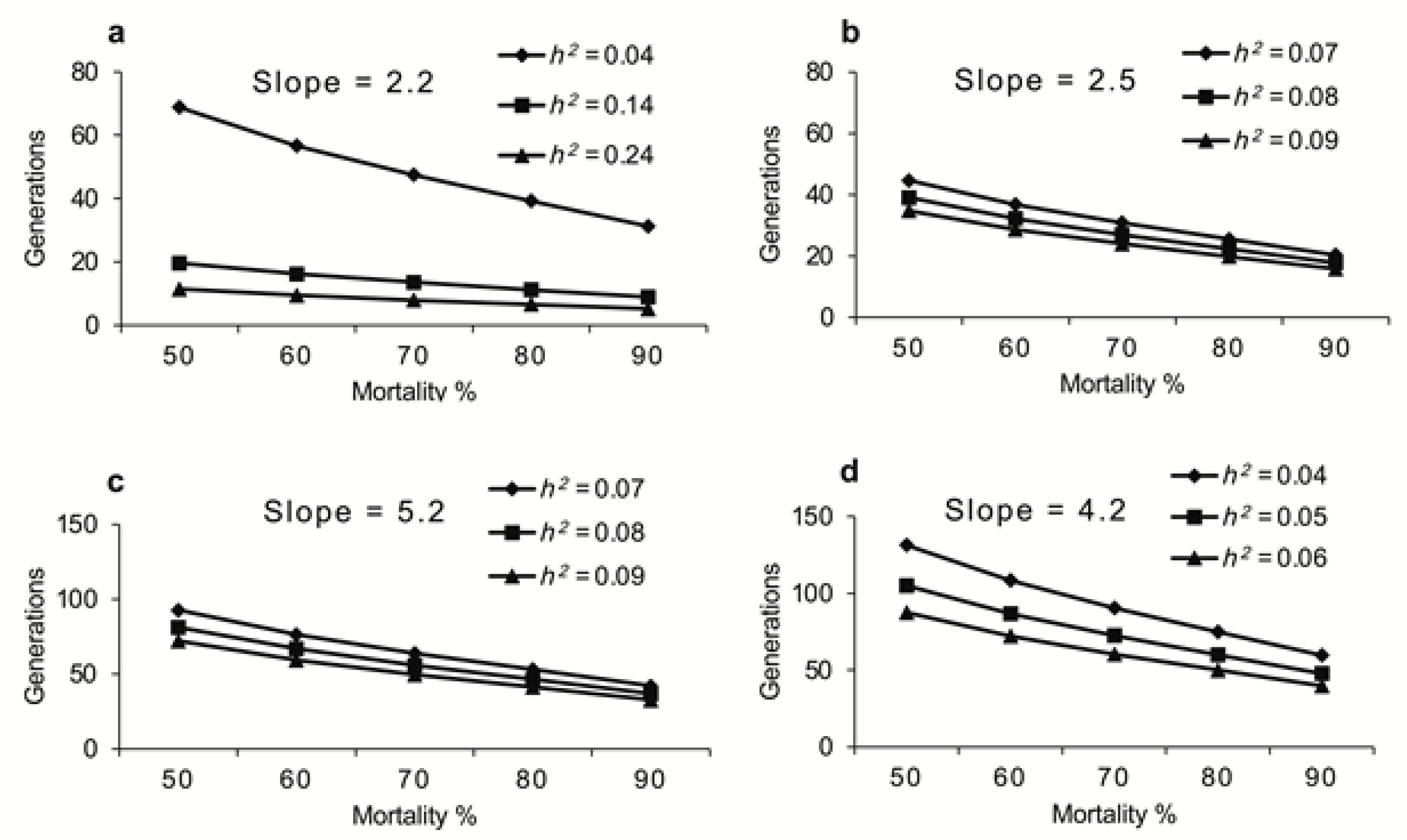
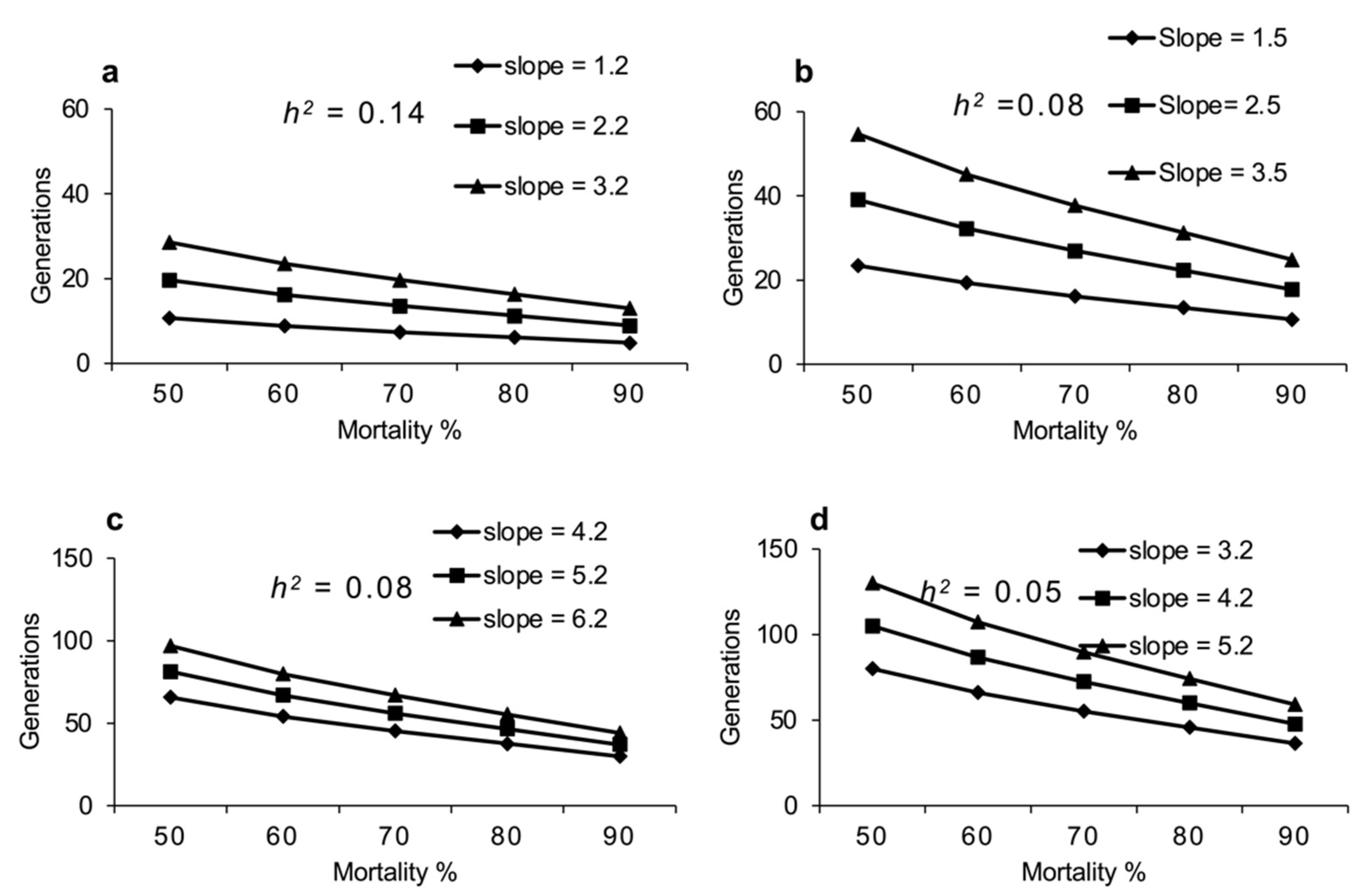
2.6. Projection of Number of Generations for 10-Fold Increase in Resistance Development
3. Results
3.1. Selection of Drosophila suzukii Resistance to Spinosad and Malathion and Dose–Response Bioassay
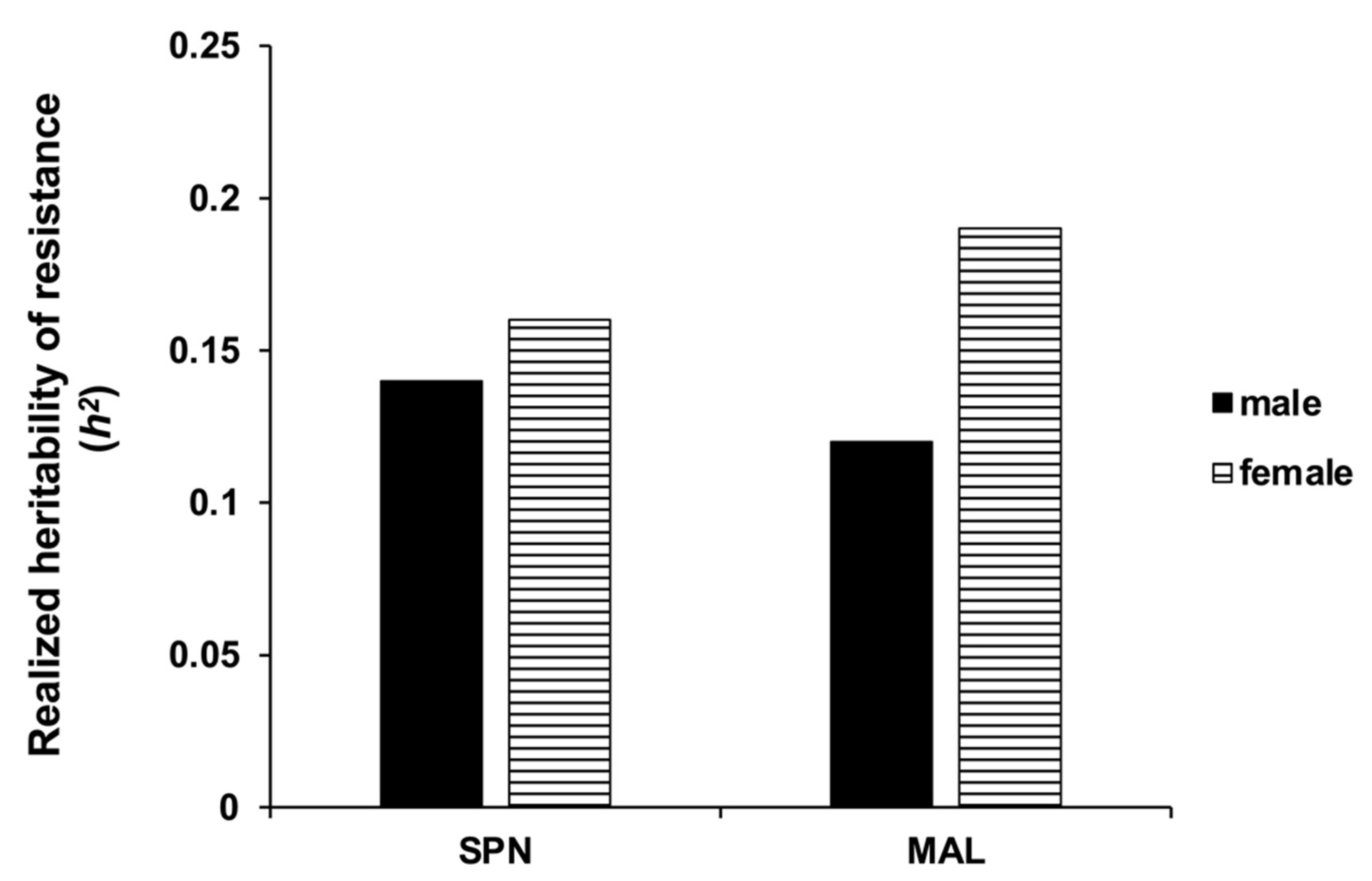
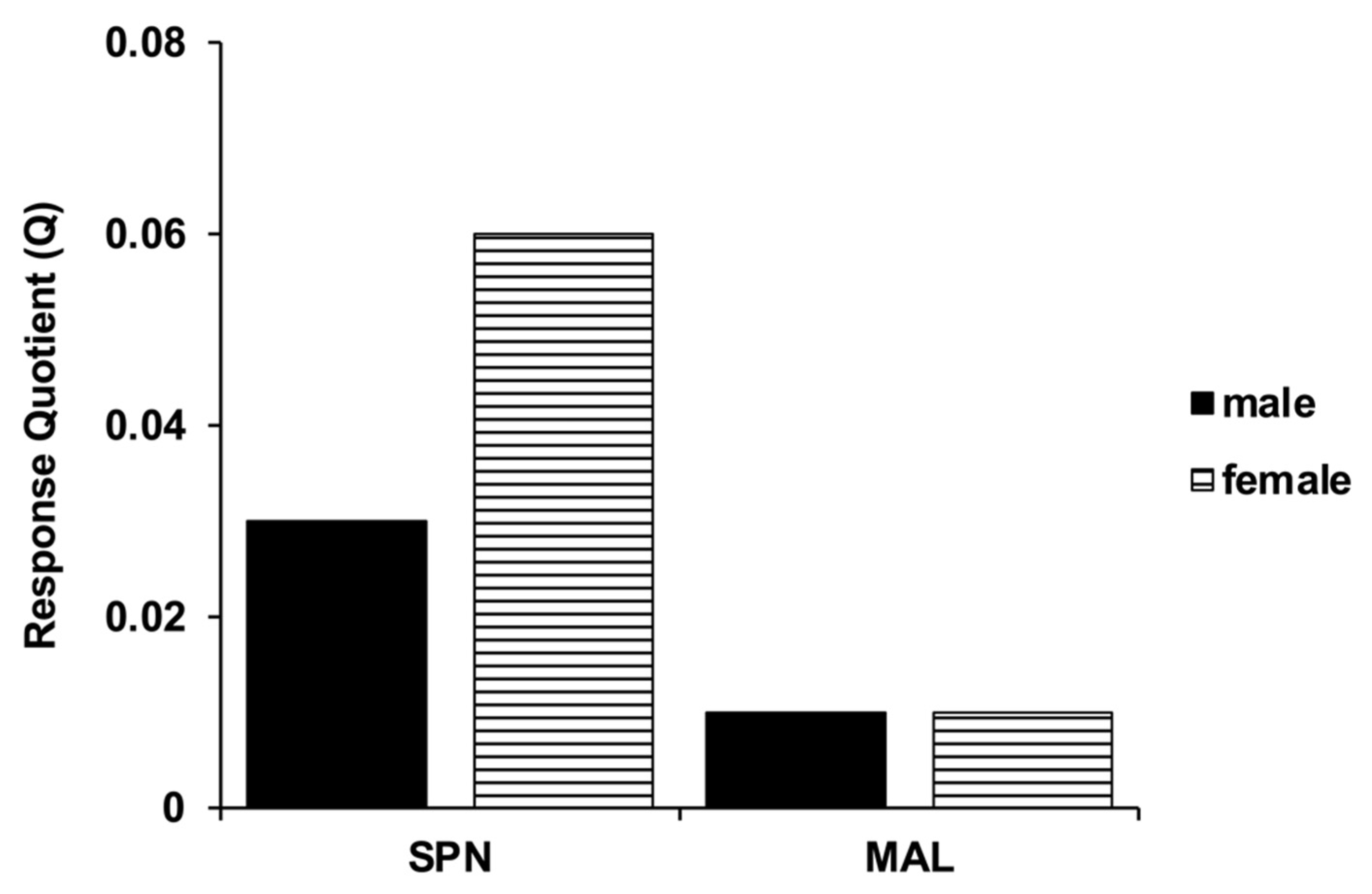
3.2. Estimation of Realized Heritability
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Bolda, M.P.; Goodhue, R.E.; Zalom, F.G. Spotted Wing Drosophila: Potential economic impact of a newly established pest. Agric. Resour. Econ. Updat. Univ. Calif. Giannini Found. 2010, 13, 5–8. [Google Scholar] [CrossRef]
- Boughdad, A.; Haddi, K.; El Bouazzati, A.; Nassiri, A.; Tahiri, A.; El Anbri, C.; Eddaya, T.; Zaid, A.; Biondi, A. First record of the invasive spotted wing Drosophila infesting berry crops in Africa. J. Pest Sci. 2021, 94, 261–271. [Google Scholar] [CrossRef]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive Pest of Ripening Soft Fruit Expanding its Geographic Range and Damage Potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Leach, H.; Van Timmeren, S.; Isaacs, R. Exclusion Netting Delays and Reduces Drosophila suzukii (Diptera: Drosophilidae) Infestation in Raspberries. J. Econ. Entomol. 2016, 109, 2151–2158. [Google Scholar] [CrossRef]
- Schöneberg, T.; Arsenault-benoit, A.; Taylor, C.M.; Butler, B.R.; Dalton, D.T.; Walton, V.M.; Petran, A.; Rogers, M.A.; Diepenbrock, L.M.; Burrack, H.J.; et al. Agriculture, Ecosystems and Environment Pruning of small fruit crops can affect habitat suitability for Drosophila suzukii. Agric. Ecosyst. Environ. 2020, 294, 106860. [Google Scholar] [CrossRef]
- Yousef, M.; Aranda-Valera, E.; Quesada-Moraga, E. Lure-and-infect and lure-and-kill devices based on Metarhizium brunneum for spotted wing Drosophila control. J. Pest Sci. 2018, 91, 227–235. [Google Scholar] [CrossRef]
- Tait, G.; Kaiser, C.; Rossi-Stacconi, M.V.; Dalton, D.T.; Anfora, G.; Walton, V.M. A food-grade gum as a management tool for Drosophila suzukii. Bull. Insectology 2018, 71, 295–307. [Google Scholar]
- Rossi Stacconi, M.V.; Tait, G.; Rendon, D.; Grassi, A.; Boyer, G.; Nieri, R.; Walton, V.M. Gumming up the Works: Field Tests of a New Food-Grade Gum as Behavioral Disruptor for Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2020, 113, 1872–1880. [Google Scholar] [CrossRef]
- Santoiemma, G.; Tonina, L.; Marini, L.; Duso, C. Integrated management of Drosophila suzukii in sweet cherry orchards. Entomol. Generalis 2020, 40, 297–305. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Lee, J.C.; Bruck, D.J. Reduced spray programs for Drosophila suzukii management in berry crops. Int. J. Pest Manag. 2016, 62, 368–377. [Google Scholar] [CrossRef]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Mota-Sanchez, D.; Wise, J.C.; Isaacs, R. Baseline susceptibility of spotted wing Drosophila (Drosophila suzukii) to four key insecticide classes. Pest Manag. Sci. 2018, 74, 78–87. [Google Scholar] [CrossRef]
- Gress, B.E.; Zalom, F.G. Identification and risk assessment of spinosad resistance in a California population of Drosophila suzukii. Pest Manag. Sci. 2019, 75, 1270–1276. [Google Scholar] [CrossRef]
- Smirle, M.J.; Zurowski, C.L.; Ayyanath, M.M.; Scott, I.M.; MacKenzie, K.E. Laboratory studies of insecticide efficacy and resistance in Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) populations from British Columbia, Canada. Pest Manag. Sci. 2017, 73, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Armes, N.J.; Jadhav, D.R.; Bond, G.S.; King, A.B.S. Insecticide resistance in Helicoverpa armigera in South India. Pestic. Sci. 1992, 34, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Brouqui, P.; Parola, P.; Raoult, D. Insecticide resistance in mosquitoes and failure of malaria control. Expert Rev. Anti. Infect. Ther. 2012, 10, 1379–1381. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.C. Insecticide resistance in Heliothis armigera in Australia. Pestic. Sci. 1988, 23, 165–176. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Weintraub, P.G.; Ishaaya, I. Status of pesticide resistance in arthropod pests in Israel. Phytoparasitica 1998, 26, 231–240. [Google Scholar] [CrossRef]
- Kakani, E.G.; Zygouridis, N.E.; Tsoumani, K.T.; Seraphides, N.; Zalom, F.G.; Mathiopoulos, K.D. Spinosad resistance development in wild olive fruit fly Bactrocera oleae (Diptera: Tephritidae) populations in California. Pest Manag. Sci. 2010, 66, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Marra, S.O.D.; Guedes, R.N.C.; Bastos, C.S.; Marra, P.H.A.; Vivan, L.M.; de Moura Zanine, A. Insecticide resistance and control failure likelihood among populations of the boll weevil (Anthonomus grandis) from mato grosso (brazil). Acta Sci. Agron. 2019, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Lai, T.; Li, J. Susceptibility of field populations of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) in China to chlorantraniliprole and the activities of detoxification enzymes. Crop Prot. 2012, 42, 217–222. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Sial, A.A.; Lanka, S.K.; Spaulding, N.R.; Isaacs, R. Development of a rapid assessment method for detecting insecticide resistance in spotted wing Drosophila (Drosophila suzukii Matsumura). Pest Manag. Sci. 2019, 75, 1782–1793. [Google Scholar] [CrossRef]
- Jaramillo, S.L.; Mehlferber, E.; Moore, P.J. Life-history trade-offs under different larval diets in Drosophila suzukii (Diptera: Drosophilidae). Physiol. Entomol. 2015, 40, 2–9. [Google Scholar] [CrossRef]
- Disi, J.O.; Van Timmeren, S.; Gress, B.; Zalom, F.; Isaacs, R.; Sial, A. Insecticide residue longevity for on-site screening of Drosophila suzukii (Matsumura) resistance. Pest Manag. Sci. 2020, 76, 2918–2924. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Resistance Risk Assessment: Realized Heritability of Resistance to Bacillus thuringiensis in Diamondback Moth (Lepidoptera: Plutellidae), Tobacco Budworm (Lepidoptera: Noctuidae), and Colorado Potato Beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1992, 85, 1551–1559. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; McGaughey, W.H. Resistance risk assessment for single and multiple insecticides: Responses of indianmeal moth (Lepidoptera: Pyralidae) to Bacillus thuringiensis. J. Econ. Entomol. 1994, 87, 834–841. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Cushing, N.L. Quantitative Genetic Analysis of Insecticide Resistance: Variation in Fenvalerate Tolerance in a Diamondback Moth (Lepidoptera: Plutellidae) Population. J. Econ. Entomol. 1989, 82, 5–10. [Google Scholar] [CrossRef]
- Singh, R.; Morton, R.A. Selection for malathion-resistance in Drosophila melanogaster. Can. J. Genet. Cytol. 1981, 23, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Crow, J.F. Genetics of Insect Resistance to Chemicals. Annu. Rev. Entomol. 1957, 2, 227–246. [Google Scholar] [CrossRef]
- Crow, J.F. Analysis of a DDT-Resistant Strain of Drosophila. J. Econ. Entomol. 1954, 47, 393–398. [Google Scholar] [CrossRef]
- Kanga, L.H.B.; Pree, D.J.; Plapp, F.W.; Van Lier, J.L. Sex-linked altered acetylcholinesterase resistance to carbamate insecticides in adults of the Oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae). Pestic. Biochem. Physiol. 2001, 71, 29–39. [Google Scholar] [CrossRef]
- Pluthero, F.G.; Threlkeld, S.F. Genetic differences in malathion avoidance and resistance in Drosophila melanogaster. J. Econ. Entomol. 1981, 74, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Feng, Z.J.; Li, T.; Lu, X.P.; Zhao, J.J.; Niu, J.Z.; Smagghe, G.; Wang, J.J. Inheritance, realized heritability, and biochemical mechanisms of malathion resistance in Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2016, 109, 299–306. [Google Scholar] [CrossRef]
- Markussen, M.D.K.; Kristensen, M. Spinosad resistance in female Musca domestica L. from a field-derived population. Pest Manag. Sci. 2012, 68, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Gerber, A.S.; Donald, P. Field experiments and natural Experiments: One definition of field experimentation. In The Oxford Handbook of Political Science; Oxford University Press: Oxford, UK, 2013; pp. 1–28. [Google Scholar] [CrossRef]
- De Keyser, R.; Cassidy, C.; Laban, S.; Gopal, P.; Pickett, J.A.; Reddy, Y.K.; Prasad, M.; Prasad, G.; Chirukandoth, S.; Senthilven, K.; et al. Insecticidal effects of deltamethrin in laboratory and field populations of Culicoides species: How effective are host-contact reduction methods in India? Parasites Vectors 2017, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selected Generation | Pop | Sex | Slope (±SE) | LC50 (ppm) (95% FL) † | RR a |
|---|---|---|---|---|---|
| Unselected | Male | 2.05 (0.38) | 20.39 (8.78–41.59) | ||
| F3 | SPN-sel | Male | 3.88 (1.14) | 26.00 (19.00–36.93) | 1.28 (0.25–0.67) |
| F8 | SPN-sel | Male | 1.97 (0.35) | 60.45 (23.43–131.21) | 2.96 (0.21–0.72) |
| F10 | SPN-sel | Male | 2.91 (0.54) | 105.79 (71.11–153.15) | 5.19 (0.15–0.44) |
| Unselected | Female | 2.27 (0.50) | 22.14 (7.16–52.88) | ||
| F3 | SPN-sel | Female | 1.96 (0.34) | 50.72 (21.28–100.82) | 2.29 (0.13–0.41) |
| F8 | SPN-sel | Female | 1.71 (0.31) | 65.73 (25.30–164.18) | 2.97 (0.18–0.68) |
| F10 | SPN-sel | Female | 2.47 (0.54) | 167.25 (95.62–387.14) | 7.55 (0.11–0.33) |
| Selected Generation | Pop | Sex | Slope (±SE) | LC50 (ppm) (95% FL) † | RR a |
|---|---|---|---|---|---|
| Unselected | Male | 3.92 (0.75) | 2.72 (1.71–3.29) | ||
| F7 | Mal-sel | Male | 3.76 (0.69) | 3.23 (1.39–4.67) | 1.19 (0.41–0.85) |
| F9 | Mal-sel | Male | 4.15 (0.83) | 3.01 (1.53–4.16) | 1.11 (0.47–1.05) |
| F11 | Mal-sel | Male | 5.72 (1.02) | 5.49 (3.55–7.28) | 2.02 (0.49–0.87) |
| Unselected | Female | 4.70 (1.03) | 3.50 (2.23–4.55) | ||
| F7 | Mal-sel | Female | 3.95 (0.74) | 3.72 (2.07–5.02) | 1.06 (0.58–1.13) |
| F9 | Mal-sel | Female | 4.71 (0.10) | 3.69 (2.77–4.45) | 1.05 (0.52–1.04) |
| F11 | Mal-sel | Female | 8.58 (1.64) | 7.81 (6.61–9.12) | 2.23 (0.48–0.78) |
| Selected Generation | Estimation of Response to Selection | Estimation of Selection Differential | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Sex | Initial LC50 (95% FL) | Final LC50 (95% FL) | R | i | Initial Slope (±SE) | Final Slope (±SE) | σp | s | |
| F3 | Male | 20.39 (8.78–41.59) | 26.00 (19.00–36.93) | 0.01 | 1.74 | 2.05(0.38) | 3.88 (1.14) | 0.34 | 0.59 |
| F8 | Male | 20.39 (8.78–41.59) | 60.45 (23.43–131.21 | 0.07 | 1.76 | 2.05(0.38) | 1.97 (0.35) | 0.5 | 0.88 |
| F10 | Male | 20.39 (8.78–41.59) | 105.79 (71.11–153.15) | 0.07 | 1.21 | 2.05(0.38) | 2.91 (0.54) | 0.4 | 0.49 |
| F3 | Female | 22.14 (7.16–52.88) | 50.72 (21.28–100.82) | 0.12 | 1.45 | 2.27(0.50) | 1.96 (0.34) | 0.47 | 0.69 |
| F8 | Male | 20.39 (8.78–41.59) | 60.45 (23.43–131.21 | 0.07 | 1.76 | 2.05(0.38) | 1.97 (0.35) | 0.5 | 0.88 |
| F10 | Female | 22.14 (7.16–52.88) | 167.25 (95.62–387.14) | 0.08 | 1.21 | 2.27(0.50) | 2.47 (0.54) | 0.42 | 0.51 |
| Selected Generation | Estimation of Response to Selection | Estimation of Selection Differential | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Sex | Initial LC50 (95% FL) | Final LC50 (95% FL) | R | i | Initial Slope (±SE) | Final Slope (±SE) | σp | s | |
| F7 | Male | 2.72 (1.71–3.29) | 3.23 (1.39–4.67) | 0.01 | 1.27 | 3.92 (0.75) | 3.76 (0.69) | 0.26 | 0.33 |
| F9 | Male | 2.72 (1.71–3.29) | 3.01 (1.53–4.16) | 0.008 | 1.1 | 3.92 (0.75) | 4.15 (0.83) | 0.25 | 0.27 |
| F11 | Male | 2.72 (1.71–3.29) | 5.49 (3.55–7.28) | 0.03 | 1.23 | 3.92 (0.75) | 5.72 (1.02) | 0.21 | 0.26 |
| F7 | Female | 3.50 (2.23–4.55) | 3.72 (2.07–5.02) | 0.004 | 1.01 | 4.70 (1.03) | 3.95 (0.74) | 0.23 | 0.23 |
| F9 | Female | 3.50 (2.23–4.55) | 3.69 (2.77–4.45) | 0.003 | 1.08 | 4.70 (1.03) | 4.71 (0.10) | 0.21 | 0.23 |
| F11 | Female | 3.50 (2.23–4.55) | 7.81 (6.61–9.12) | 0.03 | 1.04 | 4.70 (1.03) | 8.58 (1.64) | 0.15 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Disi, J.O.; Sial, A.A. Laboratory Selection and Assessment of Resistance Risk in Drosophila suzukii (Diptera: Drosophilidae) to Spinosad and Malathion. Insects 2021, 12, 794. https://doi.org/10.3390/insects12090794
Disi JO, Sial AA. Laboratory Selection and Assessment of Resistance Risk in Drosophila suzukii (Diptera: Drosophilidae) to Spinosad and Malathion. Insects. 2021; 12(9):794. https://doi.org/10.3390/insects12090794
Chicago/Turabian StyleDisi, Joseph Onwusemu, and Ashfaq A. Sial. 2021. "Laboratory Selection and Assessment of Resistance Risk in Drosophila suzukii (Diptera: Drosophilidae) to Spinosad and Malathion" Insects 12, no. 9: 794. https://doi.org/10.3390/insects12090794