
Impact of Diets Including Agro-Industrial By-Products on the Fatty Acid and Sterol Profiles of Larvae Biomass from Ephestia kuehniella, Tenebrio molitor and Hermetia illucens



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Effect of Diet Composition on Fatty Acids Profiles
3.1.1. Hermetia illucens
3.1.2. Tenebrio molitor
3.1.3. Ephestia kuehniella
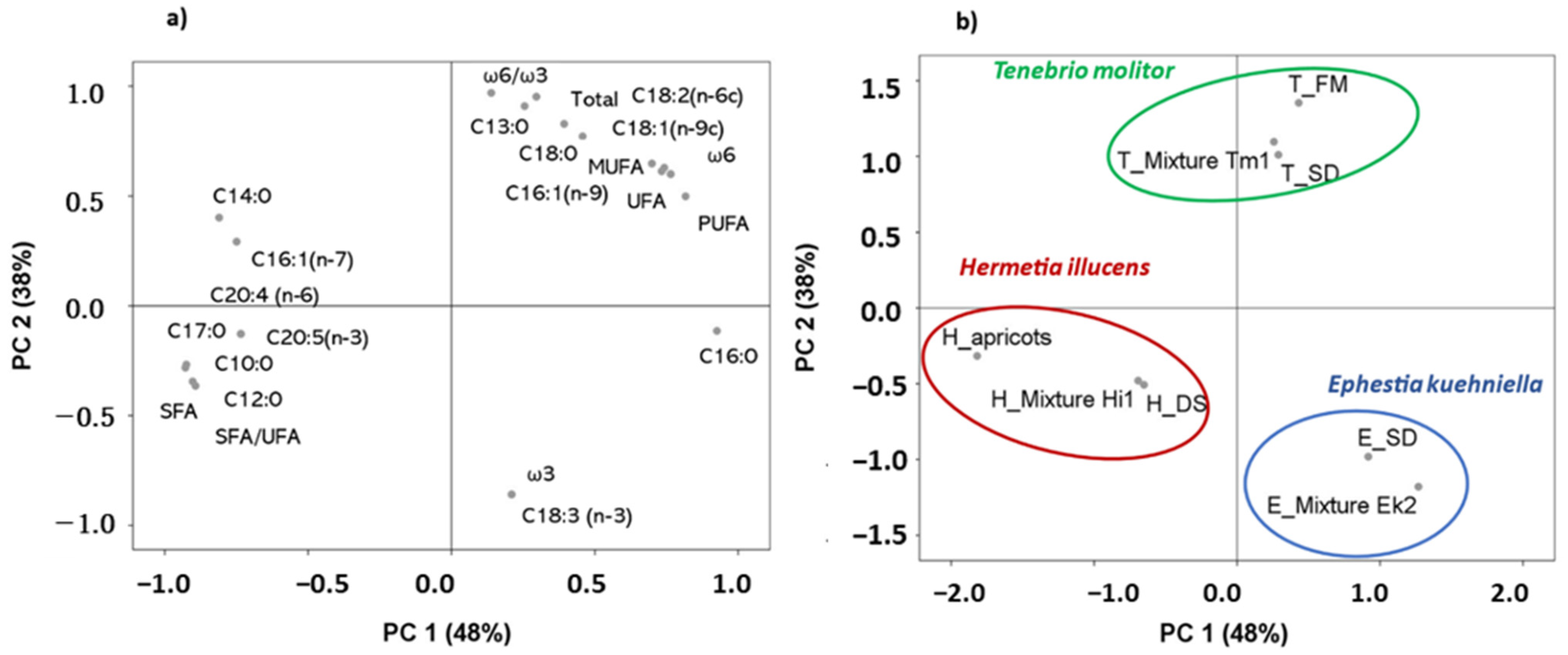
3.2. Differences in Fatty Acid Profiles Between Insect Species
- H. illucens: margaric, capric, lauric, EPA, SFA, and SFA:UFA;
- T. molitor: ω6:ω3, tridecylic, stearic, total fatty acids;
- E. kuehniella: ω3, and alpha linolenic.
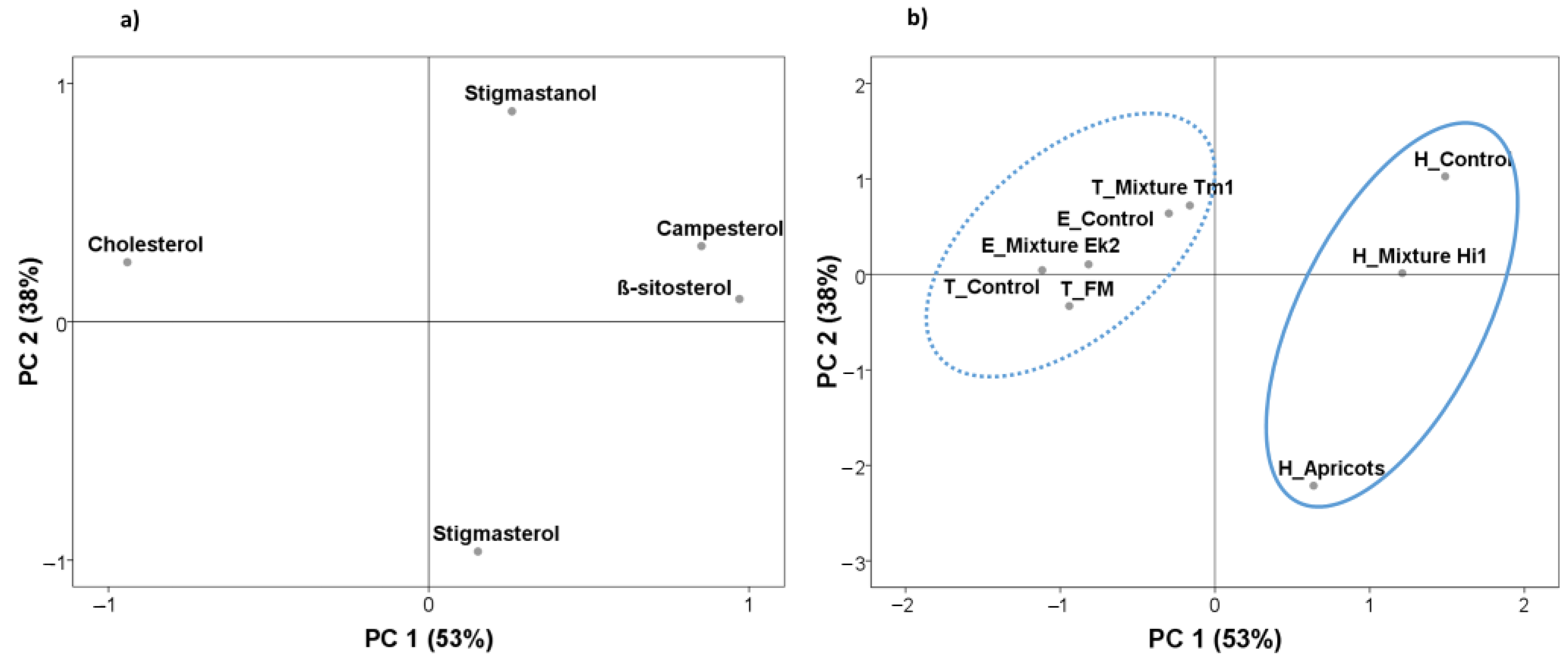
3.3. Effect of Diet Composition on Sterols Profiles
3.4. Differences in Fatty Sterol Profiles between Insects Species
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez-García, R.; Campos, D.A.; Aguilar, C.N.; Madureira, A.R.; Pintado, M. Valorization of melon fruit (Cucumis melo L.) by-products: Phytochemical and Biofunctional properties with Emphasis on Recent Trends and Advances. Trends Food Sci. Technol. 2020, 99, 507–519. [Google Scholar] [CrossRef]
- Correddu, F.; Lunesu, M.F.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can agro-industrial by-products rich in polyphenols be advantageously used in the feeding and nutrition of dairy small ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae produced at an industrial scale. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.C.; Pereira, R.N.; Rodrigues, A.S.; Teixeira, J.A.; Pintado, M.E. The use of emergent technologies to extract added value compounds from grape by-products. Trends Food Sci. Technol. 2020, 106, 182–197. [Google Scholar] [CrossRef]
- Barcelos, M.C.S.; Ramos, C.L.; Kuddus, M.; Rodriguez-Couto, S.; Srivastava, N.; Ramteke, P.W.; Mishra, P.K.; Molina, G. Enzymatic potential for the valorization of agro-industrial by-products. Biotechnol. Lett. 2020, 42, 1799–1827. [Google Scholar] [CrossRef] [PubMed]
- Ur Rehman, K.; Rehman, A.; Cai, M.; Zheng, L.; Xiao, X.; Somroo, A.A.; Wang, H.; Li, W.; Yu, Z.; Zhang, J. Conversion of mixtures of dairy manure and soybean curd residue by black soldier fly larvae (Hermetia illucens L.). J. Clean. Prod. 2017, 154, 366–373. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black Soldier Fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.; Ur Rehman, K.; Li, W.; Cai, M.; Li, Q.; Mazza, L.; Zhang, J.; et al. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Van Huis, A. Edible insects are the future? Proc. Nutr. Soc. 2016, 75, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect rearing: Potential, challenges, and circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)–Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Fasel, N.J.; Mene-Saffrane, L.; Ruczynski, I.; Komar, E.; Christe, P. Diet Induced Modifications of Fatty-Acid Composition in Mealworm Larvae (Tenebrio molitor). J. Food Res. 2017, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Jajić, I.; Popović, A.; Urošević, M.I.; Krstović, S.; Petrović, M.; Guljaš, D.; Samardžić, M. Fatty and amino acid profile of mealworm larvae (Tenebrio molitor L.). Biotechnol. Anim. Husb. 2020, 36, 167–180. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19. [Google Scholar] [CrossRef]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth Performance and Nutrient Composition of Mealworms (Tenebrio molitor) Fed on Fresh Plant Materials-Supplemented Diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Self-selection of two diet components by Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and its impact on fitness. Environ. Entomol. 2011, 40, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.C.; Ruiz, A.; Morales-Ramos, J.; Thomas, M.; Rojas, M.; Tomberlin, J.; Yi, L.; Han, R.; Giroud, L.; Jullien, R. Insect mass production technologies. In Insects as Sustainable Food Ingredients; Academic Press: Cambridge, MA, USA, 2016; pp. 153–201. [Google Scholar]
- Van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.B.; de Souza, R.G.; da Silva, S.R.; da Costa Feitosa, A.; Lopes, E.C.; Lima, S.B.P.; Dourado, L.R.B.; Pavan, B.E. Development of Tenebrio molitor (Coleoptera: Tenebrionidae) on Poultry Litter-Based Diets: Effect on Chemical Composition of Larvae. J. Insect Sci. 2021, 21. [Google Scholar] [CrossRef]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types. Animals 2019, 9, 281. [Google Scholar] [CrossRef] [Green Version]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Mancini, S.; Fratini, F.; Turchi, B.; Mattioli, S.; Dal Bosco, A.; Tuccinardi, T.; Nozic, S.; Paci, G. Former foodstuff products in Tenebrio molitor rearing: Effects on growth, chemical composition, microbiological load, and antioxidant status. Animals 2019, 9, 484. [Google Scholar] [CrossRef] [Green Version]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Riudavets, J.; Castañé, C.; Agustí, N.; Del Arco, L.; Diaz, I.; Castellari, M. Development and Biomass Composition of Ephestia kuehniella (Lepidoptera: Pyralidae), Tenebrio molitor (Coleoptera: Tenebrionidae), and Hermetia illucens (Diptera: Stratiomyidae) Reared on Different Byproducts of the Agri-Food Industry. J. Insect Sci. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Ichiki, R.T.; Shimoda, M.; Morioka, S. Small-scale rearing of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), in the laboratory: Low-cost and year-round rearing. Appl. Entomol. Zool. 2016, 51, 161–166. [Google Scholar] [CrossRef]
- N Ribeiro, M.A.R.C. A review of the scientific literature for optimal conditions for mass rearing Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2018, 53, 434–454. [Google Scholar] [CrossRef]
- Jacob, T.A.; Cox, P.D. The influence of temperature and humidity on the life-cycle of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Stored Prod. Res. 1977, 13, 107–118. [Google Scholar] [CrossRef]
- Barroso, F.G.; Sánchez-Muros, M.J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.L. Insects as food: Enrichment of larvae of Hermetia illucens with omega 3 fatty acids by means of dietary modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; de Meulenaer, B.; Michiels, J.; Eeckhout, M.; de Clercq, P.; de Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Galassi, G.; Jucker, C.; Parma, P.; Lupi, D.; Crovetto, G.M.; Savoldelli, S.; Colombini, S. Impact of Agro-industrial Byproducts on Bioconversion, Chemical Composition, in vitro Digestibility, and Microbiota of the Black Soldier Fly (Diptera: Stratiomyidae) Larvae. J. Insect Sci. 2021, 21. [Google Scholar] [CrossRef] [PubMed]
- Hoc, B.; Genva, M.; Fauconnier, M.L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Dayrit, F.M. The Properties of Lauric Acid and Their Significance in Coconut Oil. J. Am. Oil Chem. Soc. 2015, 92, 1–15. [Google Scholar] [CrossRef]
- Zeiger, K.; Popp, J.; Becker, A.; Hankel, J.; Visscher, C.; Klein, G.; Meemken, D. Lauric acid as feed additive—An approach to reducing Campylobacter spp. in broiler meat. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belghit, I.; Waagbø, R.; Lock, E.-J.; Liland, N.S. Insect-based diets high in lauric acid reduce liver lipids in freshwater Atlantic salmon. Aquac. Nutr. 2019, 25, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, L.; Varriale, L.; Dipineto, L.; Pace, A.; Menna, L.F.; Fioretti, A. Insect Derived Lauric Acid as Promising Alternative Strategy to Antibiotics in the Antimicrobial Resistance Scenario. Front. Microbiol. 2021, 12, 330. [Google Scholar] [CrossRef] [PubMed]
- Dvm, H. Dietary Fat and Heart Health: In Search of the Ideal Fat. Asia Pacific J. Clin. Nutr. 2002, 11, S394–S400. [Google Scholar]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Lawal, K.G.; Kavle, R.R.; Akanbi, T.O.; Mirosa, M.; Agyei, D. Enrichment in specific fatty acids profile of Tenebrio molitor and Hermetia illucens larvae through feeding. Futur. Foods 2021, 3, 100016. [Google Scholar] [CrossRef]
- Pasotto, D.; van Emmenes, L.; Cullere, M.; Giaccone, V.; Pieterse, E.; Hoffman, L.C.; Dalle Zotte, A. Inclusion of Hermetia illucens larvae reared on fish offal to the diet of broiler quails: Effect on immunity and caecal microbial populations. Czech J. Anim. Sci. 2020, 65, 213–223. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [Green Version]
- Rumpold, B.A.; Fröhling, A.; Reineke, K.; Knorr, D.; Boguslawski, S.; Ehlbeck, J.; Schlüter, O. Comparison of volumetric and surface decontamination techniques for innovative processing of mealworm larvae (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2014, 26, 232–241. [Google Scholar] [CrossRef]
- Rumpold, B.; Schlüter, O. Insect-based protein sources and their potential for human consumption: Nutritional composition and processin. Anim. Front. 2015, 5, 20–24. [Google Scholar] [CrossRef]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio molitor larvae as an alternative food source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Sobczuk-Szul, M.; Wroński, M.; Wielgosz-Groth, Z.; Mochol, M.; Rzemieniewski, A.; Nogalski, Z.; Pogorzelska-Przybyłek, P.; Purwin, C. The effect of slaughter season on the fatty acid profile in four types of fat deposits in crossbred beef bulls. Asian-Australas. J. Anim. Sci. 2013, 26, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional Value of Mealworm, Tenebrio molitor as Food Source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, F.; Ogihara, M.H.; Iga, M.; Kataoka, H. Cholesterol internalization and metabolism in insect prothoracic gland, a steroidogenic organ, via lipoproteins. Steroids 2018, 134, 110–116. [Google Scholar] [CrossRef]
- Matthäus, B.; Piofczyk, T.; Katz, H.; Pudel, F. Renewable Resources from Insects: Exploitation, Properties, and Refining of Fat Obtained by Cold-Pressing from Hermetia illucens (Black Soldier Fly) Larvae. Eur. J. Lipid Sci. Technol. 2019, 121. [Google Scholar] [CrossRef]
- Otero, P.; Gutierrez-Docio, A.; Navarro del Hierro, J.; Reglero, G.; Martin, D. Extracts from the edible insects Acheta domesticus and Tenebrio molitor with improved fatty acid profile due to ultrasound assisted or pressurized liquid extraction. Food Chem. 2020, 314, 126200. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Bueno, R.P.; González-Fernández, M.J.; Sánchez-Muros-Lozano, M.J.; García-Barroso, F.; Guil-Guerrero, J.L. Fatty acid profiles and cholesterol content of seven insect species assessed by several extraction systems. Eur. Food Res. Technol. 2016, 242, 1471–1477. [Google Scholar] [CrossRef]
- Mlček, J.; Adámková, A.; Adámek, M.; Borkovcová, M.; Bednářová, M.; Knížková, I. Fat from Tenebrionidae bugs–Sterols content, fatty acid profiles, and cardiovascular risk indexes. Pol. J. Food Nutr. Sci. 2019, 69, 247–254. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Diet Description | Diet Composition | Mortality (%) | Weight (mg) | Development Time (Weeks) | |||
|---|---|---|---|---|---|---|---|---|
| Moisture (%) | Total Fat (% Dry Matter) | Total Protein (% Dry Matter) | Pupae | Adult | ||||
| H. illucens | ||||||||
| Control [26] | 24.4% wheat bran, 14.6% rabbit feed (Cryspy Muesli, Versele-Laga), 2.4% yeast, and 58.6%, water | 61.2 | 1.8 | 7.5 | 21.6 ± 6.6 | 225.7 ± 10.3 | 3 | 5 |
| AP-R | 100% apricots (raw) | 88.5 | 0.2 | 0.6 | 83.9 ± 2.9 | 309.9 ± 31.8 | 3 | 8 |
| Mixture Hi1 | 25% brewer’s spent grain (dried), 12.5% feed mill by-products and 62.5% brewer’s spent yeast (raw) | 59.1 | 3.0 | 9.6 | 32.1 ± 7.7 | 292.0 ± 11.9 | 3 | 5 |
| T. molitor | ||||||||
| Control [27] | 48.5% whole flour wheat, 36.7% wheat bran, and 14.7% pet food (Ultima dog food with chicken, Affinity Petcare) | 12.1 | 6.0 | 17.2 | 8.0 ± 5.8 | 156.5 ± 4.4 | 11 | 13 |
| FM | 100% feed mill by-products (broken cereal grains) | 8.7 | 2.9 | 11.3 | 39.7 ± 3.3 | 142.4 ± 5.5 | 16 | 18 |
| Mixture Tm1 | 36.8% brewer’s spent grain (dried), 52.5% feed mill by-products: and 10.5% brewer’s spent yeast (dried) | 7.3 | 4.9 | 19.3 | 22.8 ± 2.3 | 140.1 ± 12.1 | 11 | 13 |
| E. kuehniella | ||||||||
| Control [28] | 67% whole wheat flour and 33%, commercial yeast | 11.4 | 2.6 | 16.4 | 60.4 ± 2.4 | 43.5 ± 2.5 | 3 | 4 |
| Mixture Ek2 | 67% feed mill by-products and 33%, brewer’s spent yeast (dried) | 6.5 | 2.7 | 22.4 | 38.4 ± 7.5 | 18.3 ± 2.5 | 6 | 7 |
| Fatty Acids and Indices | Control | Mixture Hi1 | AP-R | Significance |
|---|---|---|---|---|
| Capric [C10:0] (%) | 0.57 ± 0.11 a | 0.52 ± 0.26 a | 1.36 ± 0.06 b | ** |
| Lauric [C12:0] (%) | 50.83 ± 2.74 a | 49.86 ± 2.43 a | 55.39 ± 0.62 b | * |
| Tridecylic |C13:0] (%) | n.d. | n.d. | n.d. | |
| Myristic [C14:0] (%) | 8.74 ± 0.32 | 8.34 ± 0.88 | 7.50 ± 0.14 | ns |
| Palmitic [C16:0] (%) | 11.92 ± 0.46 a | 13.96 ± 0.75 b | 11.99 ± 0.21 a | ** |
| Hypogeic [C16:1(n-9c)] (%) | 0.80 ± 0.24 b | 0.46 ± 0.07 a | 0.19 ± 0.00 a | ** |
| Palmitoleic [C16:1(n-7c)] (%) | 1.71 ± 0.14 a | 2.00 ± 0.35 a | 4.35 ± 0.07 b | *** |
| Margaric [C17:0] (%) | 0.18 ± 0.03 a | 0.22 ± 0.09 a | 0.38 ± 0.01 b | ** |
| Stearic [C18:0] (%) | 1.61 ± 0.08 a | 1.89 ± 0.34 ab | 2.18 ± 0.05 b | * |
| Oleic [C18:1(n-9c)] (%) | 9.17 ± 0.69 | 11.35 ± 1.07 | 9.64 ± 0.71 | ns |
| Linoleic [C18:2(n-6c)] (%) | 12.7 ± 1.74 c | 10.47 ± 0.33 b | 4.44 ± 0.09 a | * |
| Alpha linolenic [C18:3(n-3c)] (%) | 1.77 ± 0.08 b | 0.93 ± 0.09 a | 1.85 ± 0.06 b | *** |
| Arachidonic [C20:4(n-6c)] (%) | n.d. | n.d. | 0.17 ± 0.00 | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (%) | n.d. | n.d. | 0.57 ± 0.02 | |
| Capric [C10:0] (mg/100 mg) | 0.16 ± 0.04 a | 0.13 ± 0.07 a | 0.37 ± 0.02 b | ** |
| Lauric [C12:0] (mg/100 mg) | 13.83 ± 1.81 a | 11.84 ± 0.9 a | 15.07 ± 0.31 b | * |
| Tridecylic |C13:0] (mg/100 mg) | n.d. | n.d. | n.d. | |
| Myristic [C14:0] (mg/100 mg) | 2.38 ± 0.26 | 1.98 ± 0.23 | 2.04 ± 0.01 | ns |
| Palmitic [C16:0] (mg/100 mg) | 3.23 ± 0.12 a | 3.33 ± 0.45 b | 3.26 ± 0.02 a | ** |
| Hypogeic [C16:1(n-9c)] (mg/100 mg) | 0.21 ± 0.05 b | 0.11 ± 0.01 a | 0.05 ± 0.00 a | ** |
| Palmitoleic [C16:1(n-7c)] (mg/100 mg) | 0.46 ± 0.01 a | 0.47 ± 0.04 a | 1.18 ± 0.01 b | *** |
| Margaric [C17:0] (mg/100 mg) | 0.05 ± 0.00 a | 0.05 ± 0.04 a | 0.10 ± 0.00 b | ** |
| Stearic [C18:0] (mg/100 mg) | 0.44 ± 0.05 a | 0.45 ± 0.11 ab | 0.59 ± 0.00 b | * |
| Oleic [C18:1(n-9c)] (mg/100 mg) | 2.48 ± 0.04 | 2.71 ± 0.43 | 2.63 ± 0.24 | ns |
| Linoleic [C18:2(n-6c)] (mg/100 mg) | 3.43 ± 0.28 c | 2.49 ± 0.28 b | 1.21 ± 0.04 a | * |
| Alpha linolenic [C18:3(n-3c)] (mg/100 mg) | 0.48 ± 0.03 b | 0.22 ± 0.03 a | 0.50 ± 0.01 b | *** |
| Arachidonic [C20:4(n-6c)] (mg/100 mg) | n.d. | n.d. | 0.05 ± 0.00 | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (mg/100 mg) | n.d. | n.d. | 0.15 ± 0.00 | |
| Sum of fatty acids (mg/100 mg) | 27.15 ± 2.03 | 23.78 ± 2.07 | 27.21 ± 0.53 | ns |
| SFA (mg/100 mg) | 20.08 ± 2.27 | 17.77 ± 1.46 | 21.44 ± 0.33 | ns |
| MUFA (mg/100 mg) | 3.16 ± 0.06 a | 3.29 ± 0.38 a | 3.86 ± 0.25 b | * |
| PUFA (mg/100 mg) | 3.90 ± 0.27 c | 2.72 ± 0.31 b | 1.71 ± 0.04 a | *** |
| ω6 (mg/100 mg) | 3.43 ± 0.28 c | 2.49 ± 0.30 b | 1.21 ± 0.04 a | *** |
| ω3 (mg/100 mg) | 0.48 ± 0.03 b | 0.22 ± 0.03 a | 0.5 ± 0.01 b | *** |
| ω6:ω3 | 7.17 ± 0.74 b | 11.33 ± 1.69 c | 2.4 ± 0.05 a | *** |
| UFA (mg/100 mg) | 7.06 ± 0.31 b | 6.00 ± 0.67 a | 5.57 ± 0.29 a | * |
| SFA:UFA | 2.85 ± 0.43 a | 2.97 ± 0.17 a | 3.86 ± 0.18 b | * |
| Fatty Acids and Indices | Control | FM | Mixture Tm1 | Significance |
|---|---|---|---|---|
| Capric [C10:0] (%) | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 | ns |
| Lauric [C12:0] (%) | 0.46 ± 0.03 b | 0.37 ± 0.02 a | 0.41 ± 0.06 b | ** |
| Tridecylic [C13:0] (%) | 0.05 ± 0.00 | 0.03 ± 0.00 | 0.04 ± 0.01 | ns |
| Myristic [C14:0] (%) | 4.61 ± 0.11 b | 4.19 ± 0.14 a | 4.68 ± 0.39 b | * |
| Palmitic [C16:0] (%) | 18.20 ± 0.48 | 16.12 ± 0.48 | 17.40 ± 0.82 | ns |
| Hypogeic [C16:1(n-9c)] (%) | 1.69 ± 0.07 b | 1.54 ± 0.05a | 1.53 ± 0.15 a | ** |
| Palmitoleic [C16:1(n-7c)] (%) | 1.68 ± 0.01 | 1.60 ± 0.03 | 2.05 ± 0.15 | ns |
| Margaric [C17:0] (%) | n.d. | n.d. | n.d. | |
| Stearic [C18:0] (%) | 3.58 ± 0.12 a | 3.64 ± 0.18 b | 3.38 ± 0.57 b | ** |
| Oleic [C18:1(n-9c)] (%) | 49.63 ± 0.50 a | 52.37 ± 0.60 b | 51.95 ± 0.94 b | *** |
| Linoleic [C18:2(n-6c)] (%) | 19.72 ± 0.11 b | 19.82 ± 0.17 b | 18.21 ± 0.28 a | ** |
| Alpha linolenic [C18:3(n-3c)] (%) | 0.36 ± 0.02 | 0.29 ± 0.01 | 0.34 ± 0.02 | ns |
| Arachidonic [C20:4(n-6c)] (%) | n.d. | n.d. | n.d. | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (%) | n.d. | n.d. | n.d. | |
| Capric [C10:0] (mg/100 mg) | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ns |
| Lauric [C12:0] (mg/100 mg) | 0.15 ± 0.02 | 0.14 ± 0.01 | 0.14 ± 0.02 | ns |
| Tridecylic [C13:0] (mg/100 mg) | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ns |
| Myristic [C14:0] (mg/100 mg) | 1.47 ± 0.11 | 1.59 ± 0.16 | 1.61 ± 0.19 | ns |
| Palmitic [C16:0] (mg/100 mg) | 5.80 ± 0.17 | 6.11 ± 0.60 | 5.98 ± 0.30 | ns |
| Hypogeic [C16:1(n-9c)] (mg/100 mg) | 0.54 ± 0.04 | 0.58 ± 0.02 | 0.53 ± 0.07 | ns |
| Palmitoleic [C16:1(n-7c)] (mg/100 mg) | 0.53 ± 0.03 a | 0.61 ± 0.05 ab | 0.71 ± 0.07 b | * |
| Margaric [C17:0] (mg/100 mg) | n.d. | n.d. | n.d. | |
| Stearic [C18:0] (mg/100 mg) | 1.14 ± 0.02 | 1.38 ± 0.10 | 1.16 ± 0.16 | ns |
| Oleic [C18:1(n-9c)] (mg/100 mg) | 15.83 ± 0.92 a | 19.82 ± 1.25 b | 17.85 ± 0.67 b | ** |
| Linoleic [C18:2(n-6c)] (mg/100 mg) | 6.29 ± 0.32 a | 7.50 ± 0.56 b | 6.26 ± 0.30 a | * |
| Alpha linolenic [C18:3(n-3c)] (mg/100 mg) | 0.11 ± 0.01 | 0.11 ± 0.00 | 0.12 ± 0.01 | ns |
| Arachidonic [C20:4(n-6c)] (mg/100 mg) | n.d. | n.d. | n.d. | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (mg/100 mg) | n.d. | n.d. | n.d. | |
| Sum of fatty acids (mg/100 mg) | 31.89 ± 1.60 a | 37.86 ± 2.71 b | 34.36 ± 1.21 ab | * |
| SFA (mg/100 mg) | 8.58 ± 0.31 | 9.24 ± 0.84 | 8.91 ± 0.35 | ns |
| MUFA (mg/100 mg) | 16.91 ± 0.98 a | 21.01 ± 1.32 b | 19.08 ± 0.81 b | ** |
| PUFA (mg/100 mg) | 6.41 ± 0.33 a | 7.62 ± 0.56 b | 6.38 ± 0.30 a | * |
| ω6 (mg/100 mg) | 6.29 ± 0.32 a | 7.50 ± 0.56 b | 6.26 ± 0.30 a | * |
| ω3 (mg/100 mg) | 0.11 ± 0.01 | 0.11 ± 0.00 | 0.12 ± 0.01 | ns |
| ω6:ω3 | 54.91 ± 2.82 a | 66.74 ± 2.65 b | 53.97 ± 1.50 a | ** |
| UFA (mg/100 mg) | 23.31 ± 1.30 a | 28.62 ± 1.87 b | 25.46 ± 1.06 a | * |
| SFA:UFA | 0.37 ± 0.01 b | 0.32 ± 0.01 a | 0.35 ± 0.02 b | * |
| Fatty Acids and Indices | Control | Mixture Ek2 | Significance |
|---|---|---|---|
| Capric [C10:0] (%) | n.d. | n.d. | |
| Lauric [C12:0] (%) | n.d. | n.d. | |
| Tridecylic [C13:0] (%) | n.d. | n.d. | |
| Myristic [C14:0] (%) | 0.19 ± 0.01 | 0.32 ± 0.04 | ** |
| Palmitic [C16:0] (%) | 29.02 ± 0.12 | 30.34 ± 0.54 | * |
| Hypogeic [C16:1(n-9c)] (%) | n.d. | n.d. | |
| Palmitoleic [C16:1(n-7c)] (%) | 1.24 ± 0.03 | 1.21 ± 0.06 | ns |
| Margaric [C17:0] (%) | n.d. | n.d. | |
| Stearic [C18:0] (%) | 1.69 ± 0.05 | 3.44 ± 0.49 | ** |
| Oleic [C18:1(n-9c)] (%) | 51.42 ± 0.53 | 40.68 ± 2.47 | ** |
| Linoleic [C18:2(n-6c)] (%) | 14.43 ± 0.28 | 20.72 ± 1.25 | ** |
| Alpha linolenic [C18:3(n-3c)] (%) | 2.01 ± 0.10 | 3.29 ± 0.24 | ** |
| Arachidonic [C20:4(n-6c)] (%) | n.d. | n.d. | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (%) | n.d. | n.d. | |
| Capric [C10:0] (mg/100 mg) | n.d. | n.d. | |
| Lauric [C12:0] (mg/100 mg) | n.d. | n.d. | |
| Tridecylic C13:0 (mg/100 mg) | n.d. | n.d. | |
| Myristic [C14:0] (mg/100 mg) | 0.05 ± 0.01 | 0.09 ± 0.01 | ** |
| Palmitic [C16:0] (mg/100 mg) | 8.11 ± 0.47 | 8.65 ± 0.45 | ns |
| Hypogeic [C16:1(n-9c)] (mg/100 mg) | n.d. | n.d. | |
| Palmitoleic [C16:1(n-7c)] (mg/100 mg) | 0.35 ± 0.02 | 0.34 ± 0.04 | ns |
| Margaric [C17:0] (mg/100 mg) | n.d. | n.d. | |
| Stearic [C18:0] (mg/100 mg) | 0.47 ± 0.04 | 0.98 ± 0.09 | ** |
| Oleic [C18:1(n-9c)] (mg/100 mg) | 14.35 ± 0.73 | 11.63 ± 1.45 | * |
| Linoleic [C18:2(n-6c)] (mg/100 mg) | 4.03 ± 0.26 | 5.90 ± 0.24 | ** |
| Alpha linolenic [C18:3(n-3c)] (mg/100 mg) | 0.56 ± 0.05 | 0.94 ± 0.06 | ** |
| Arachidonic [C20:4(n-6c)] (mg/100 mg) | n.d. | n.d. | |
| Eicosapentaenoic (EPA) [C20:5(n-3c)] (mg/100 mg) | n.d. | n.d. | |
| Sum of fatty acids (mg/100 mg) | 27.92 ± 1.54 | 28.53 ± 1.95 | ns |
| SFA (mg/100 mg) | 8.63 ± 0.51 | 9.72 ± 0.42 | * |
| MUFA (mg/100 mg) | 14.70 ± 0.75 | 11.98 ± 1.49 | * |
| PUFA (mg/100 mg) | 4.59 ± 0.31 | 6.83 ± 0.30 | ** |
| ω6 (mg/100 mg) | 4.03 ± 0.26 | 5.90 ± 0.24 | ** |
| ω3 (mg/100 mg) | 0.56 ± 0.05 | 0.94 ± 0.06 | ** |
| ω6:ω3 | 7.21 ± 0.02 | 6.03 ± 0.13 | ** |
| UFA (mg/100 mg) | 19.29 ± 1.03 | 18.81 ± 1.56 | ns |
| SFA:UFA | 0.45 ± 0.00 | 0.52 ± 0.02 | *** |
| Sterols | H. illucens | T. molitor | E. kuehniella | |||||
|---|---|---|---|---|---|---|---|---|
| Control | Mixture Hi1 | Apricots | Control | FM | Mixture Tm1 | Control | Mixture Ek2 | |
| Cholesterol | 0.031 ± 0.004 a | 0.225 ± 0.022 c | 0.048 ± 0.008 b | 1.037 ± 0.058 b | 0.935 ± 0.048 b | 0.646 ± 0.103 a | 0.873 ± 0.15 a | 1.13 ± 0.127 b |
| Campesterol | 0.554 ± 0.054 c | 0.478 ± 0.017 b | 0.122 ± 0.0 4 a | 0.078 ± 0.014 | 0.083 ± 0.020 | 0.073 ± 0.092 | 0.132 ± 0.012 | 0.136 ± 0.010 |
| Stigmasterol | n.d. | 0.043 ± 0.003 a | 0.154 ± 0.010 b | 0.018 ± 0.002 a | 0.044 ± 0.003 b | 0.029 ± 0.012 a | 0.014 ± 0.006 a | 0.035 ± 0.008 b |
| β-sitosterol | 1.035 ± 0.090 b | 1.017 ± 0.012 a | n.d. | 0.168 ± 0.027 a | 0.209 ± 0.027 b | 0.171 ± 0.072 ab | 0.383 ± 0.018 | 0.364 ± 0.028 |
| Stigmastanol | 0.113 ± 0.012 b | 0.062 ± 0.023 a | n.d. | 0.039 ± 0.006 a | 0.034 ± 0.004 a | 0.123 ± 0.008 b | 0.091 ± 0.027 b | 0.054 ± 0.016 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boukid, F.; Riudavets, J.; del Arco, L.; Castellari, M. Impact of Diets Including Agro-Industrial By-Products on the Fatty Acid and Sterol Profiles of Larvae Biomass from Ephestia kuehniella, Tenebrio molitor and Hermetia illucens. Insects 2021, 12, 672. https://doi.org/10.3390/insects12080672
Boukid F, Riudavets J, del Arco L, Castellari M. Impact of Diets Including Agro-Industrial By-Products on the Fatty Acid and Sterol Profiles of Larvae Biomass from Ephestia kuehniella, Tenebrio molitor and Hermetia illucens. Insects. 2021; 12(8):672. https://doi.org/10.3390/insects12080672
Chicago/Turabian StyleBoukid, Fatma, Jordi Riudavets, Lidia del Arco, and Massimo Castellari. 2021. "Impact of Diets Including Agro-Industrial By-Products on the Fatty Acid and Sterol Profiles of Larvae Biomass from Ephestia kuehniella, Tenebrio molitor and Hermetia illucens" Insects 12, no. 8: 672. https://doi.org/10.3390/insects12080672