
A Novel Genetic Sexing Strain of Anastrepha ludens for Cost-Effective Sterile Insect Technique Applications: Improved Genetic Stability and Rearing Efficiency

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
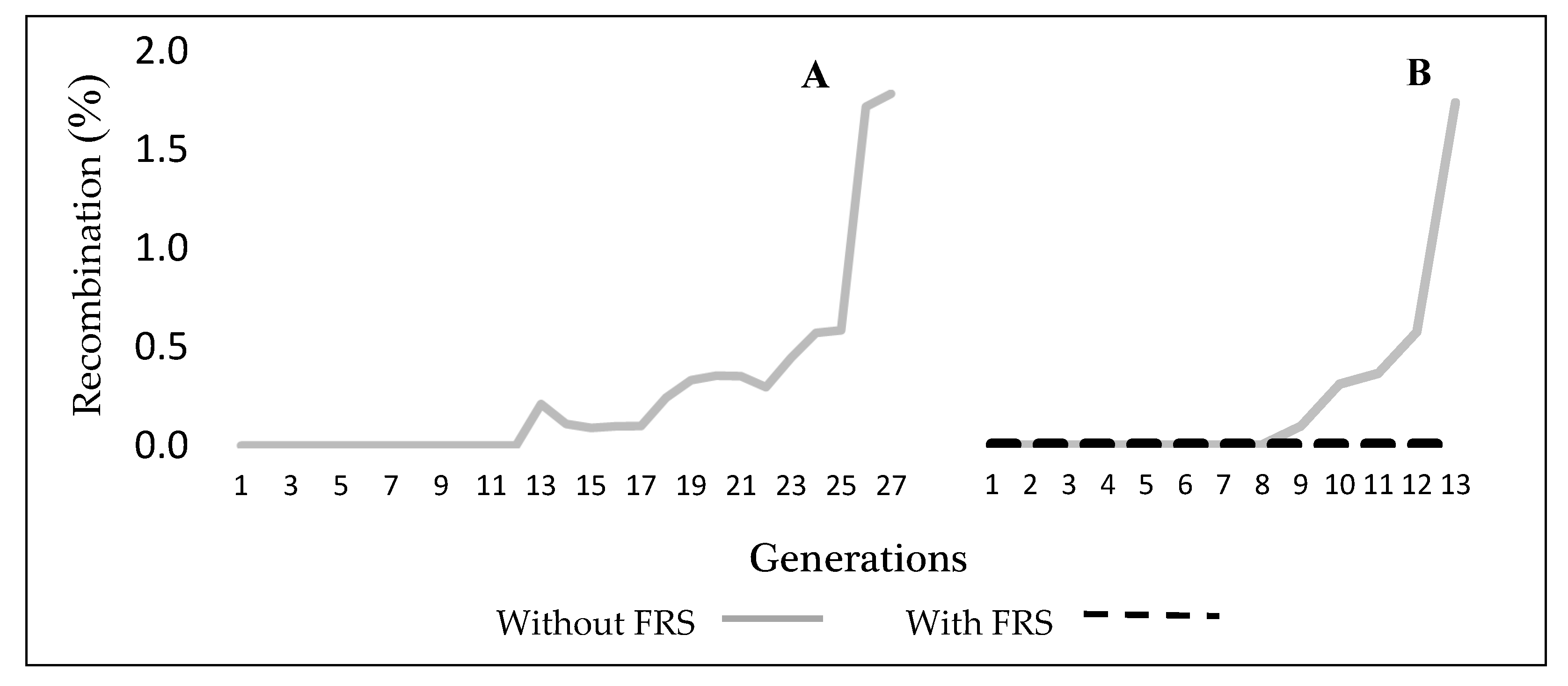
3.1. Induction of New Translocations and Selection of New Genetic Sexing Strain (GSS)
3.2. Fecundity and Fertility
3.3. Genetic Stability and Productivity of the Selected GSS GUA10
3.4. Quality Control Analysis under Laboratory Conditions
3.5. Irradiation doses Response Curve for GUA10
3.6. Field Cage Tests
3.7. Cytogenetic Analysis of GUA10 Translocation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- APHIS. Anastrepha Ludens, Mexican Fruit Fly Host List. 2016. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/fruit_flies/downloads/host-lists/mexfly-host-list.pdf (accessed on 24 September 2019).
- Copeland, R.S.; Wharton, R.A.; Luke, Q.; Meyer, M. Indigenous hosts of Ceratitis capitata (Diptera: Tephritidae) in Kenya. Ann. Ent. Soc. Am. 2002, 95, 672–694. [Google Scholar] [CrossRef]
- Molina-Nery, M.; Ruiz-Montoya, L.; Zepeda-Cisneros, C.; Liedo, P. Genetic structure of populations of Anastrepha ludens (Diptera: Tephritidae) in Mexico. Fla. Entomol. 2014, 97, 1648–1661. [Google Scholar] [CrossRef]
- Hernández, E.; Aceituno-Medina, M.; Toledo, J.; Bravo, B.; Caro-Corrales, J.; Montoya, P.; Mangan, R. The Effects of a Modified Hot Water Treatment on Anastrepha ludens (Diptera: Tephritidae)-Infested Mango. J. Econ. Entomol. 2017, 1–9. [Google Scholar] [CrossRef]
- Aluja, M.; Birke, A.; Ceymann, M.; Guillén, M.; Arrigoni, E.; Baumgartner, D.; Pascacio-Villafán, C.; Samietz, J. Agroecosystem resilience to an invasive insect species that could expand its geographical range in response to global climate change. Agric. Ecosist. Environ. 2014, 186, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.F. Sterile-male method of population control. Science 1959, 130, 902–904. [Google Scholar] [CrossRef]
- Knipling, E.F. Sterile insect technique as a screwworm control measure: The concept and its development. In Symposium on Eradication of the Screwworm from the United States and Mexico; Graham, O.H., Ed.; Miscellaneous Publications of the Entomological Society of America: College Park, MD, USA, 1985; pp. 4–7. [Google Scholar]
- Klassen, W.; Curtis, C.F. History of the Sterile Insect Technique. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 3–36. [Google Scholar]
- Dominiak, B.C.; Westcott, A.E.; Barchia, I.M. Release of sterile Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae), at Sydney, Australia. Aust. J. Exp. Agric. 2003, 43, 519–528. [Google Scholar] [CrossRef]
- Kuba, H.; Kohama, T.; Kakinohana, H.; Yamagishi, M.; Kinjo, K. The successful eradication programs of the melon fly in Okinawa. In Fruit Fly Pests: A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1996; pp. 534–550. [Google Scholar]
- František, M.; Vreysen, M.J.B. Advances and Challenges of Using the Sterile Insect Technique for the Management of Pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Cáceres, C.; Cayol, J.P.; Enkerlin, W.; Franz, G.; Hendrichs, J.; Robinson, A.S.; Barnes, B.N. Comparison of Mediterranean fruit fly (Ceratitis capitata) (Tephritidae) bisexual and genetic sexing strains: Development, evaluation and economics. In Proceedings of the 6th International Symposium on Fruit Flies of Economic Importance, Sellenbosch, South Africa, 6–10 May 2002; Isteg Scientific Publications: Irene, South Africa, 2004; pp. 367–381. [Google Scholar]
- Klassen, W. Area-Wide Integrated Pest Management and the Sterile Insect Technique, Chapter 2.1 In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest. Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; 787p. [Google Scholar]
- Hendrichs, J.; Franz, G.; Rendon, P. Increased effectiveness and applicability of the sterile insect technique through male-only releases for control of Mediterranean fruit flies during fruiting seasons. J. Appl. Entomol. 1995, 119, 371–377. [Google Scholar] [CrossRef]
- Franz, G.; Kerremans, P. Radiation induced chromosome aberrations for the genetic analysis and manipulation of the Mediterranean fruit fly, Ceratitis capitata. In Proceedings of the International Symposium on Management of Insect Pests. Nuclear and Related Molecular and Genetic Techniques, Vienna, Austria, 19–23 October 1992; International Atomic Energy Agency: Vienna, Austria, 1992; pp. 187–194. [Google Scholar]
- Franz, G. Genetic sexing strains in Mediterranean fruit fly, an example for other species amenable to largescale rearing for the sterile insect technique. In Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 427–451. [Google Scholar]
- Robinson, A.S.; Van Heemert, C. Ceratitis capitata—A suitable case for genetic sexing. Genetica 1982, 58, 229–237. [Google Scholar] [CrossRef]
- Rössler, Y. Automated sexing of Ceratitis capitata [Dip.: Tephritidae]: The development of strains with inherited, sex-limited pupal color dimorphism. Entomophaga 1979, 24, 411–416. [Google Scholar] [CrossRef]
- Franz, G.; Bourtzis, K.; Cáceres, C. Practical and Operational Genetic Sexing Systems Based on Classical Genetic Approaches in Fruit Flies, an Example for Other Species Amenable to Large-Scale Rearing for the Sterile Insect Technique. In Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Ratón, FL, USA, 2021; pp. 575–604. [Google Scholar]
- Fisher, K.; Caceres, C.; Rendón, P. A filter rearing system for mass reared sexing strains of Mediterranean fruit fly (Diptera: Tephritidae). In Area-Wide Control of Fruit Flies and Other Pests; Tan, K.H., Ed.; Penerbit Universiti Sains Malaysia: Penang, Malaysia, 2000; pp. 543–550. [Google Scholar]
- Franz, G.; Gencheva, E.; Kerremans, P. Improved stability of genetic sex-separation strains for the Mediterranean fruit-fly, Ceratitis capitata. Genome 1994, 37, 72–82. [Google Scholar] [CrossRef]
- Zepeda-Cisneros, C.S.; Meza-Hernández, J.S.; García-Martínez, V.; Ibañez-Palacios, J.; Zacharopoulou, A.; Franz, G. Development, genetic and cytogenetic analyses of genetic sexing strains of the Mexican fruit fly. Anastrepha ludens Loew (Diptera: Tephritidae). BMC Genet. 2014, 15 (Suppl. 2), 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zepeda-Cisneros, C.S. Desarrollo de cepas de sexado genético. In Moscas de la Fruta: Fundamentos y Procedimientos para su Manejo; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: México City, Mexico, 2010; pp. 333–342. [Google Scholar]
- Braga, R.; Caceres, C.; Islam, A.; Wornoayporn, V.; Enkerlin, W. Diets based on soybean protein for Mediterranean fruit fly. Pesq. Agropec. Bras. 2006, 41, 705–708. [Google Scholar] [CrossRef]
- Orozco-Dávila, D.; Quintero-Fong, L.; Hernández, E.; Solís, E.; Artiaga, T.; Hernández, R.; Ortega, C.; Montoya, P. Mass rearing and sterile insect releases for the control of Anastrepha spp. pests in Mexico—A review. Entomol. Exp. Appl. 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies, Version 6.0. 2014; International Atomic Energy Agency: Vienna, Austria, 2014; 164p. [Google Scholar]
- Resilva, S.S.; Hernandez, E.; Obra, G.B. Radiation Sterilization of Mexican Fruit Fly Anastrepha ludens (Loew) Based on Pupal Eye Color. Philipp. J. Crop. Sci. 2018, 148, 45–50. [Google Scholar]
- Zacharopoulou, A. Cytogenetic analysis of mitotic and salivary gland chromosomes in the medfly C. capitata. Genome 1987, 29, 67–71. [Google Scholar] [CrossRef]
- Zacharopoulou, A. Polytene chromosome maps in the medfly C. capitata. Genome 1990, 33, 184–197. [Google Scholar] [CrossRef]
- García-Martinez, V.; Hernandez-Ortiz, E.; Zepeta Cisneros, C.S.; Robinson, A.S.; Zacharopoulou, A.; Franz, G. Mitotic and polytene chromosome analysis in the Mexican fruit fly, Anastrepha ludens (Loew) (Diptera: Tephritidae). Genome 2009, 52, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Casanoves, F.; Di Rienzo, J.A.; Robledo, C.W. InfoSat Statistical Software: Manual del Usuario; Editorial Brujas: Córdoba, Argentina, 2008; 336p. [Google Scholar]
- Orozco-Dávila, D.; Adriano-Amaya, M.; Quintero-Fong, L.; Salvador-Figueroa, M. Sterility and Sexual Competitiveness of Tapachula-7 Anastrepha ludens Males Irradiated at Different Doses. PLoS ONE 2015. [Google Scholar] [CrossRef] [Green Version]
- Rendón, P.; McInnis, D.O.; Lance, D.; Stewart, J. Medfly (Diptera: Tephritidae) genetic sexing: Large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Santos, E.; Rendón, P.; Ruiz-Montoya, L.; Toledo, J.; Liedo, P. Effect of Irradiation Doses on Sterility and Biological Security in a Genetically Modified Strain of the Mediterranean Fruit Fly (Diptera: Tephritidae). J. Econ. Entomol. 2017, 110, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Hendrichs, J.; Robinson, A.S.; Cayol, J.P.; Enkerlin, W. Medfly area wide sterile insect technique programmes for prevention, suppression or eradication: The importance of mating behavior studies. Fla. Entomol. 2002, 85, 1–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | n | % Recombinants (♀ in Brown Pupae) | % Recombinants (♂ in Black Pupae) | Egg to Pupae Conversion Rate (E:P) |
|---|---|---|---|---|
| 197 | 6 | 25.03 a ± 1.397 | 22.19 a ± 1.677 | 0.36 c ± 0.005 × |
| 109 | 17 | 0.28 c ± 0.153 | 0.20 c ± 0.110 | 0.31 c ± 0.019 × |
| 48 | 6 | 5.30 b ± 2.672 | 9.97 b ± 4.538 | 0.30 c ± 0.028 × |
| 45 | 14 | 0.27 c ± 0.220 | 0.00 c ± 0.000 | 0.34 c ± 0.016 × |
| 9 | 17 | 2.80 b ± 1.569 | 0.97 c ± 0.533 | 0.29 c ± 0.019 × |
| 87 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.34 c ± 0.012 ×× |
| 125 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.33 c ± 0.019 ×× |
| 116 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.32 c ± 0.013 ×× |
| 154 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.32 c ± 0.014 ×× |
| 103 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.32 c ± 0.025 ×× |
| 66 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.31 c ± 0.017 ×× |
| 10 | 17 | 0.00 c ± 0.000 | 0.00 c ± 0.000 | 0.41 b ± 0.016 ××× |
| TAP7 (Control) | 17 | 3.02 b ± 0.725 | 0.56 c ± 0.202 | 0.32 c ± 0.015 |
| Wild (Reference) | 13 | – | – | 0.74 a ± 0.029 |
| Cross | Low Density (without Stress) | High Density (with Stress) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Male | × | Female | n | Eggs/♀/Day | n | % Egg Hatch | n | Eggs/♀/Day | n | % Egg Hatch |
| GUA10 | × | GUA10 | 90 | 49.50 a± 1.33 | 105 | 86.92 a± 0.71 | 14 | 29.14 a± 2.30 | 10 | 85.54 a± 1.33 |
| TAP7 | × | TAP7 | 82 | 25.57 b± 1.39 | 96 | 74.68 b± 0.74 | 12 | 22.75 a± 2.49 | 9 | 70.67 b± 1.40 |
| Scale | Strain | Larval Recovery | Egg: Pupae Conversion Efficiency | Millions of Pupae/ton of Larval Diet |
|---|---|---|---|---|
| Medium | GUA10 | 0.18 a ± 0.003 | 0.26 a ± 0.01 | 4.17 a ± 0.102 |
| TAP7 | 0.13 b ± 0.007 | 0.17 b ± 0.008 | 2.81 b ± 0.090 | |
| Large | GUA10 | 0.14 a ± 0.003 | 0.28 a ± 0.005 | 3.60 a ± 0.081 |
| TAP7 | 0.11 b ± 0.005 | 0.21 b ± 0.009 | 2.78 b ± 0.117 |
| Males (Brown Pupae) | Females (Black Pupae) | Recombinants | |||
|---|---|---|---|---|---|
| Strain | Emergence | Flyers | Emergence | Flyers | (♀ in Brown Pupae) |
| GUA10 | 93.36 a ± 0.652 | 89.62 a± 0.924 | 89.54 a ± 0.618 | 81.65 a ± 1.033 | 1.25 a ± 0.199 |
| TAP7 | 78.67 b ± 0.681 | 78.76 b ± 0.364 | 81.59 b ± 0.571 | 77.37 b ± 0.669 | 12.65 b ± 1.740 |
| Cross | ||||||||
|---|---|---|---|---|---|---|---|---|
| (♂GUA10 × ♂TAP7 × ♂w) × ♀w | (♂GUA10 × ♂w) × ♀w | (♂TAP7 × ♂w) × ♀w | ||||||
| Male | n | % Matings ± SE | RMPI | n | % Matings ± SE | RSI | n | % Matings ± SE |
| GUA10 | 20 | 19.10 b ± 2.32 | 0.40 a ± 0.03 | 22 | 30.73 a ± 3.29 | 0.68 a ± 0.03 | ||
| TAP7 | 20 | 16.30 b ± 2.27 | 0.32 b ± 0.02 | 0.54 b ± 0.02 | 22 | 22.73 b ± 2.56 | ||
| W | 20 | 14.70 b ± 2.26 | 22 | 16.09 b ± 2.82 | 22 | 20.55 b ± 3.07 | ||
| Control | 11 | 51.27 a ± 8.55 | 11 | 41.45 a ± 7.37 | 11 | 41.45 a ± 7.37 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Santos, E.; Rendon, P.; Gouvi, G.; Zacharopoulou, A.; Bourtzis, K.; Cáceres, C.; Bloem, K. A Novel Genetic Sexing Strain of Anastrepha ludens for Cost-Effective Sterile Insect Technique Applications: Improved Genetic Stability and Rearing Efficiency. Insects 2021, 12, 499. https://doi.org/10.3390/insects12060499
Ramírez-Santos E, Rendon P, Gouvi G, Zacharopoulou A, Bourtzis K, Cáceres C, Bloem K. A Novel Genetic Sexing Strain of Anastrepha ludens for Cost-Effective Sterile Insect Technique Applications: Improved Genetic Stability and Rearing Efficiency. Insects. 2021; 12(6):499. https://doi.org/10.3390/insects12060499
Chicago/Turabian StyleRamírez-Santos, Edwin, Pedro Rendon, Georgia Gouvi, Antigone Zacharopoulou, Kostas Bourtzis, Carlos Cáceres, and Kenneth Bloem. 2021. "A Novel Genetic Sexing Strain of Anastrepha ludens for Cost-Effective Sterile Insect Technique Applications: Improved Genetic Stability and Rearing Efficiency" Insects 12, no. 6: 499. https://doi.org/10.3390/insects12060499