
Modelling the Potential Geographic Distribution of Two Trissolcus Species for the Brown Marmorated Stink Bug, Halyomorpha halys


 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
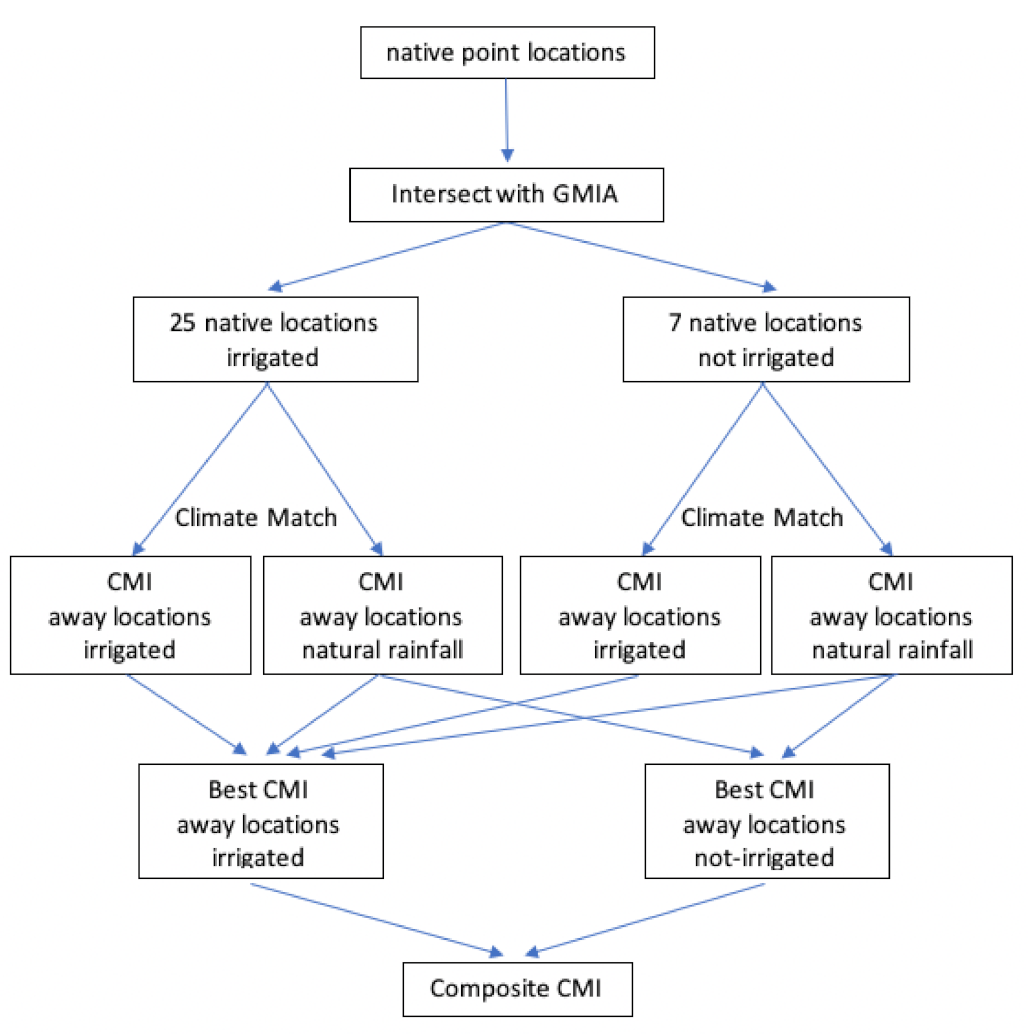
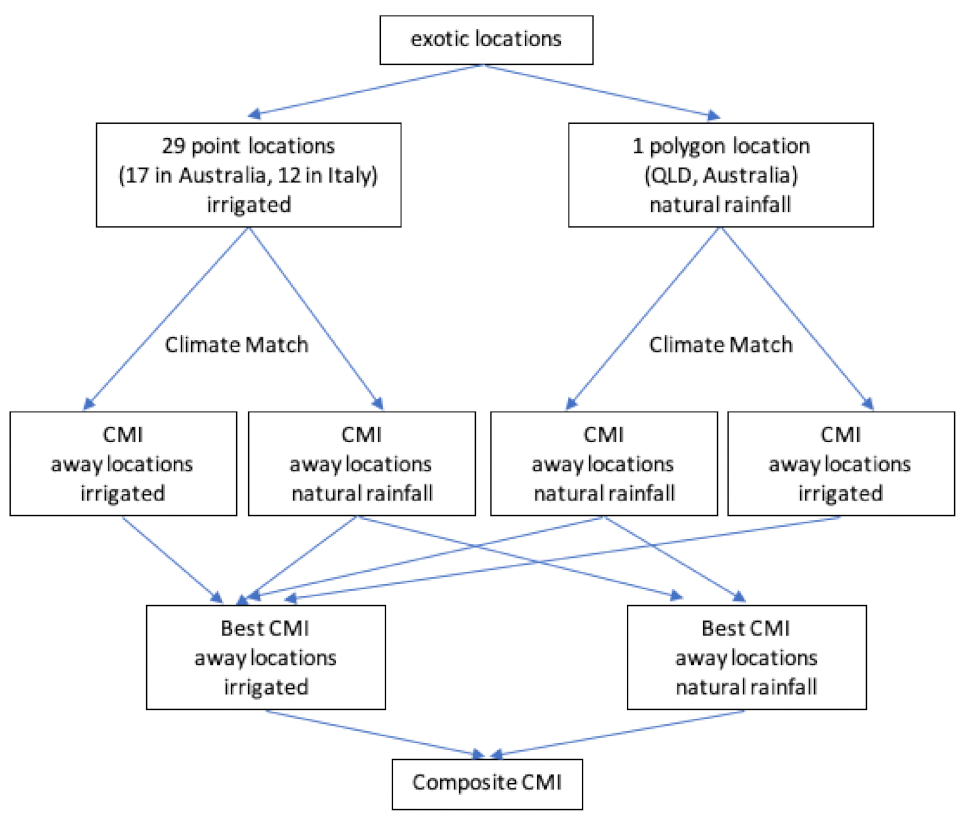
2. Materials and Methods
2.1. Location Records: Trissolcus mitsukurii
2.2. Location Records: Trissolcus japonicus
2.3. Location Records: Halyomorpha halys
2.4. Meteorological Data
2.5. CLIMEX Regional Match Climates: Trissolcus mitsukurii
2.6. CLIMEX Compare Locations
2.6.1. Moisture Index
2.6.2. Temperature Index
2.6.3. Cold Stress
2.6.4. Heat Stress
2.6.5. Dry Stress
2.6.6. Wet Stress
2.6.7. Number of Generations (PDD)
2.6.8. Model Runs
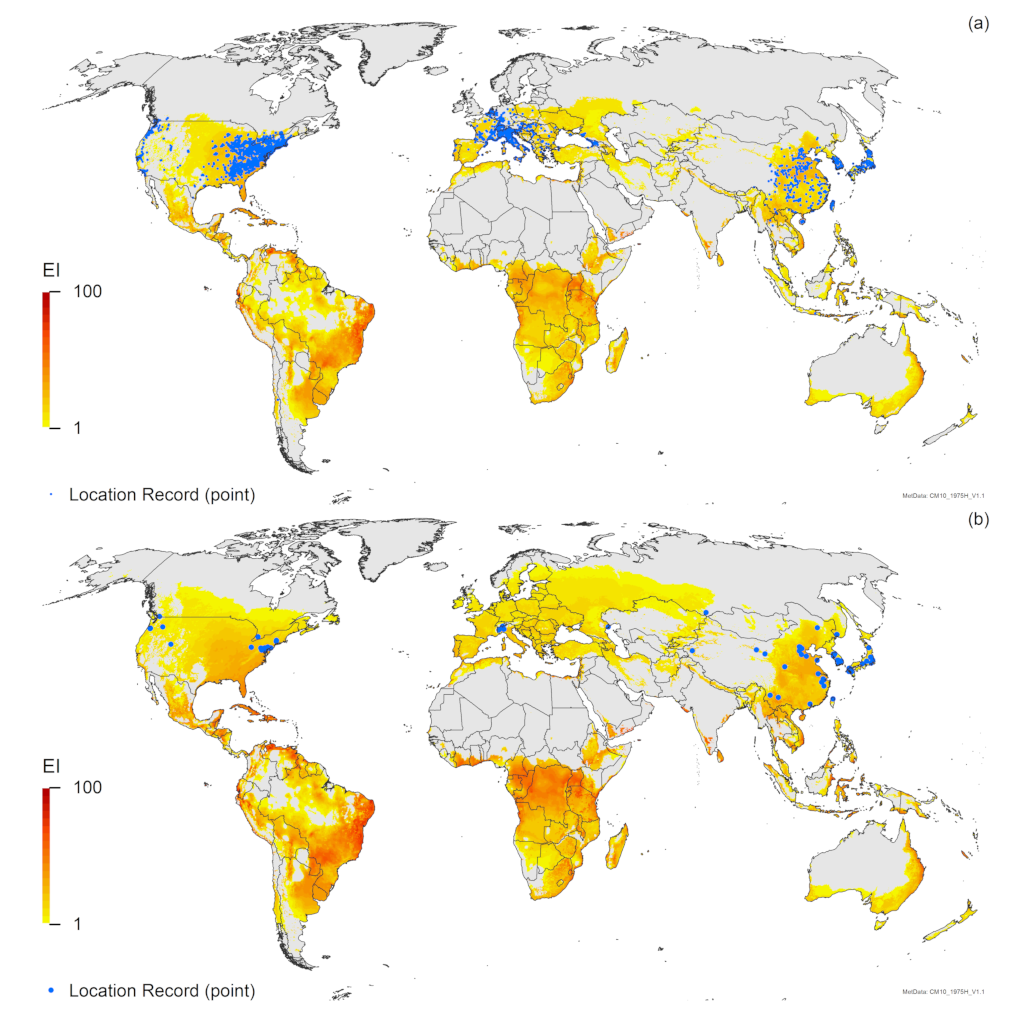
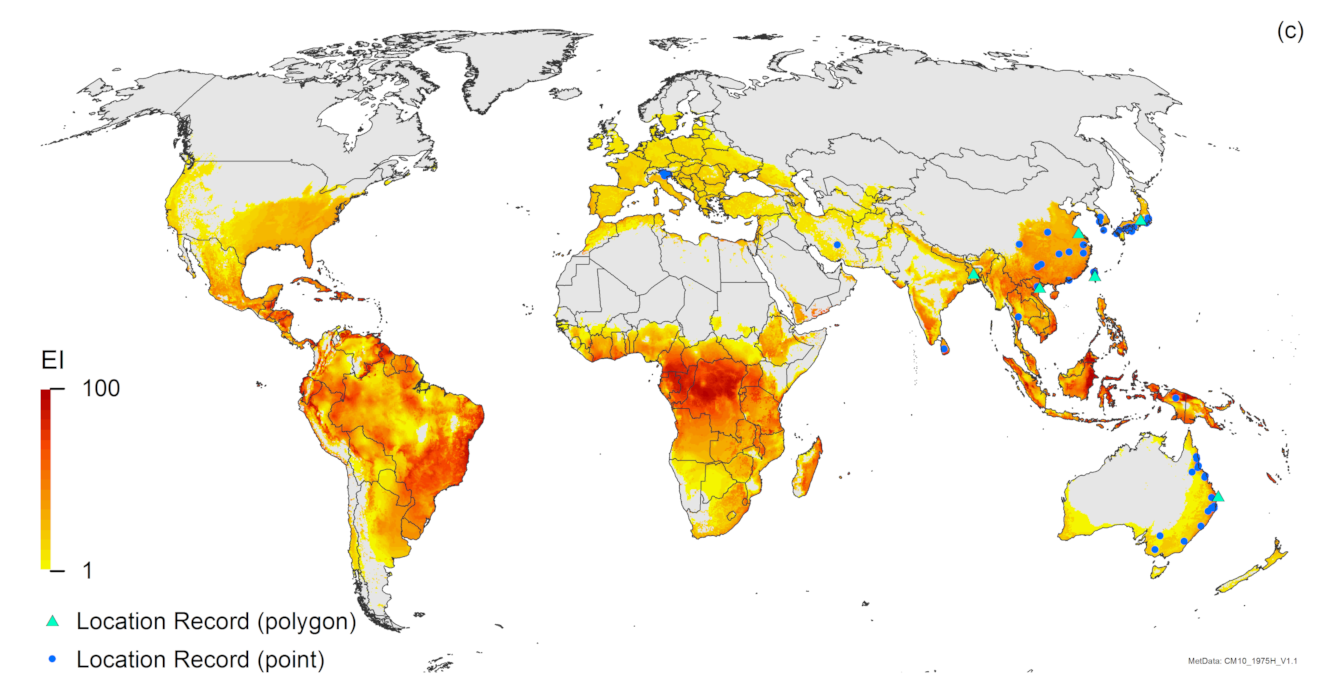
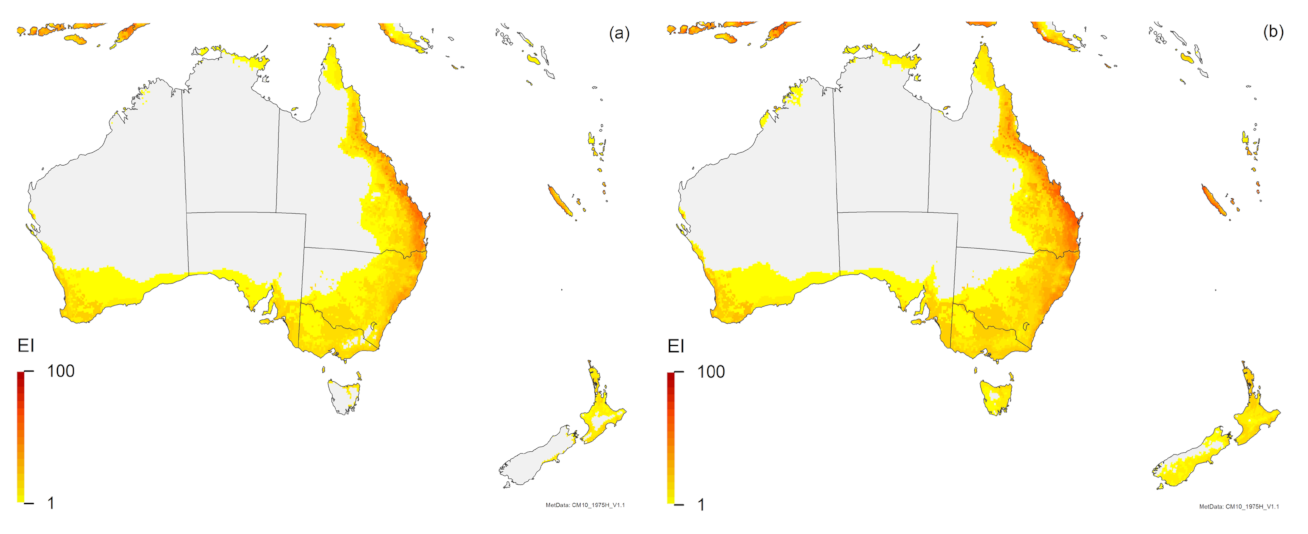
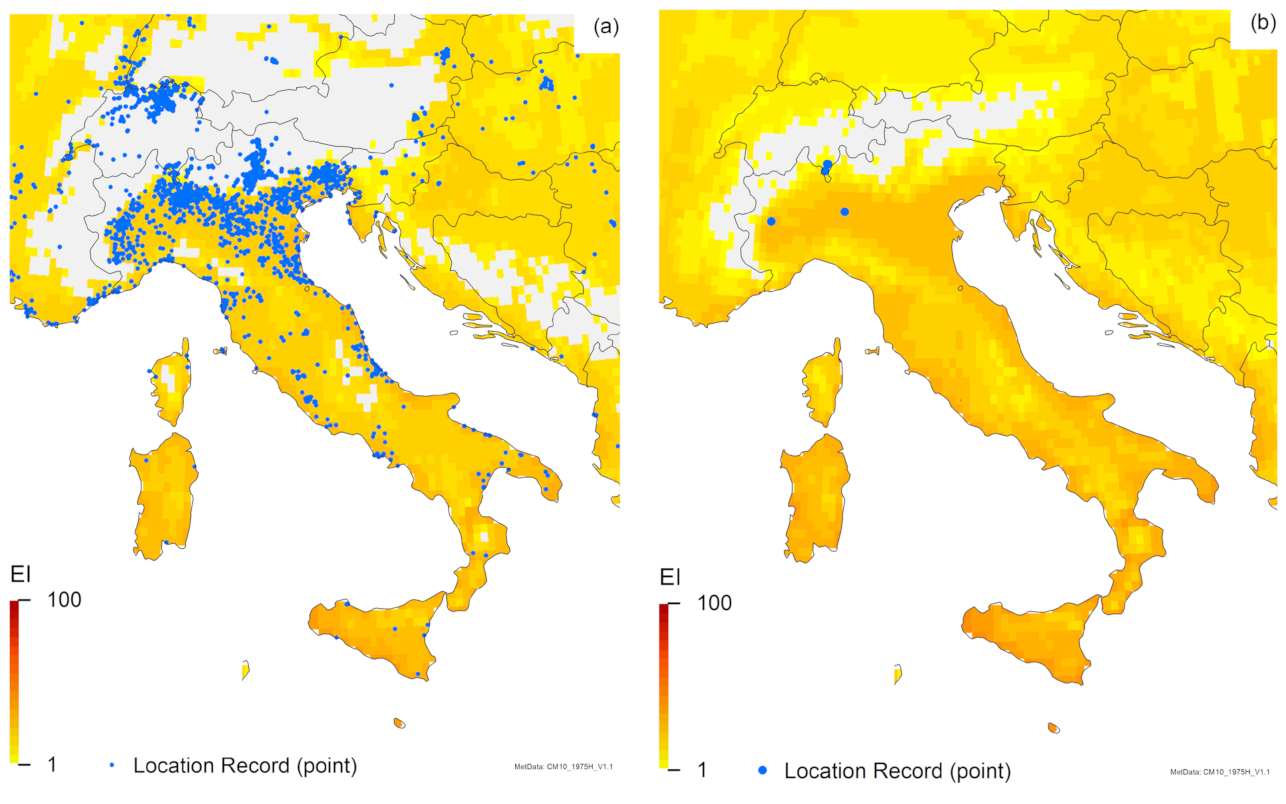
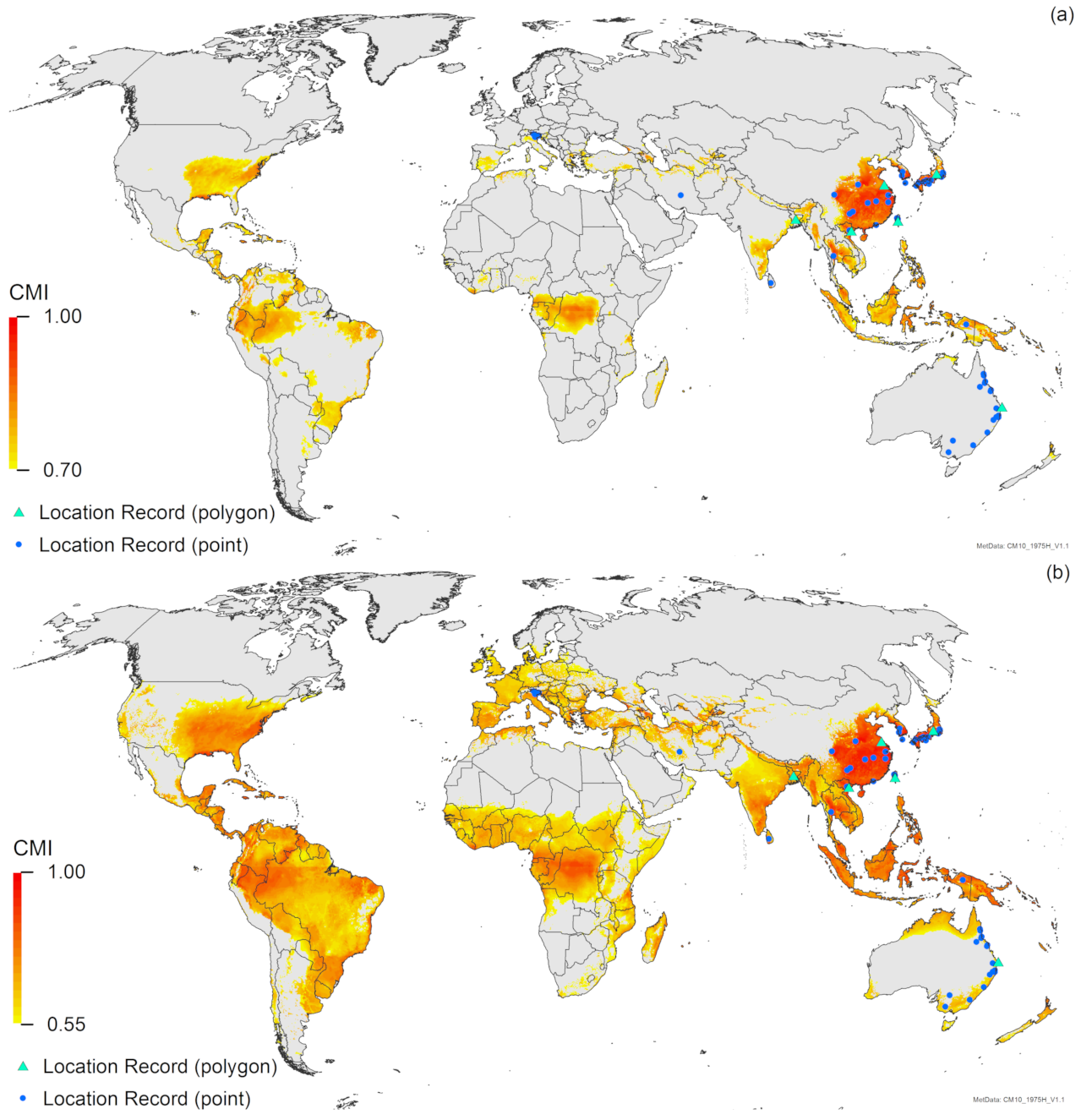
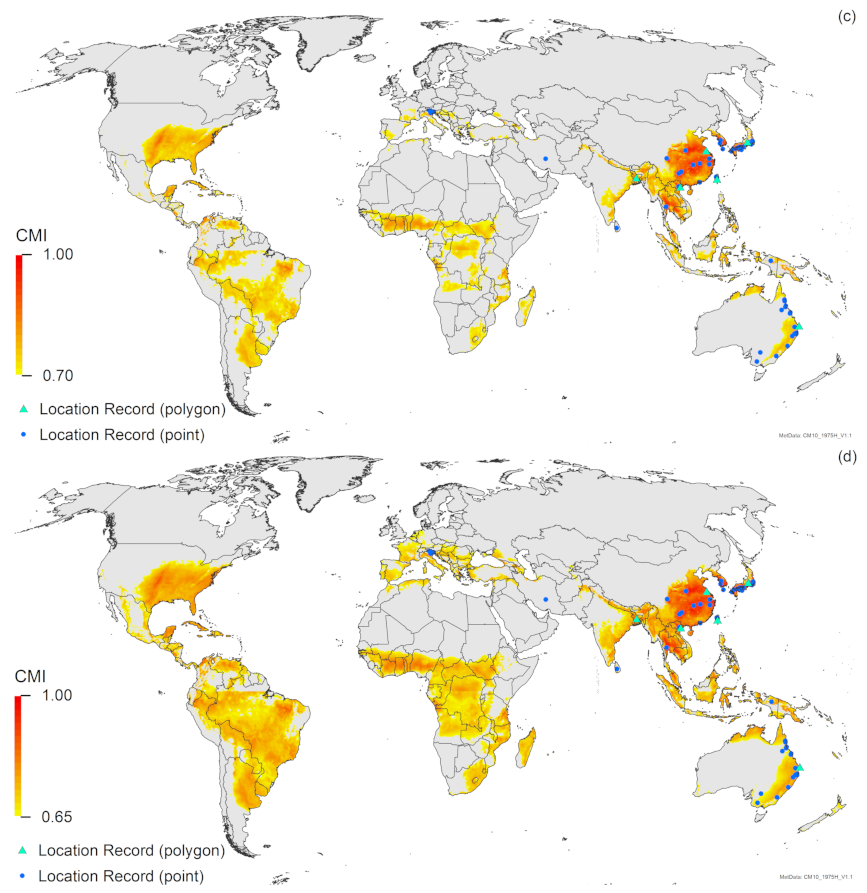
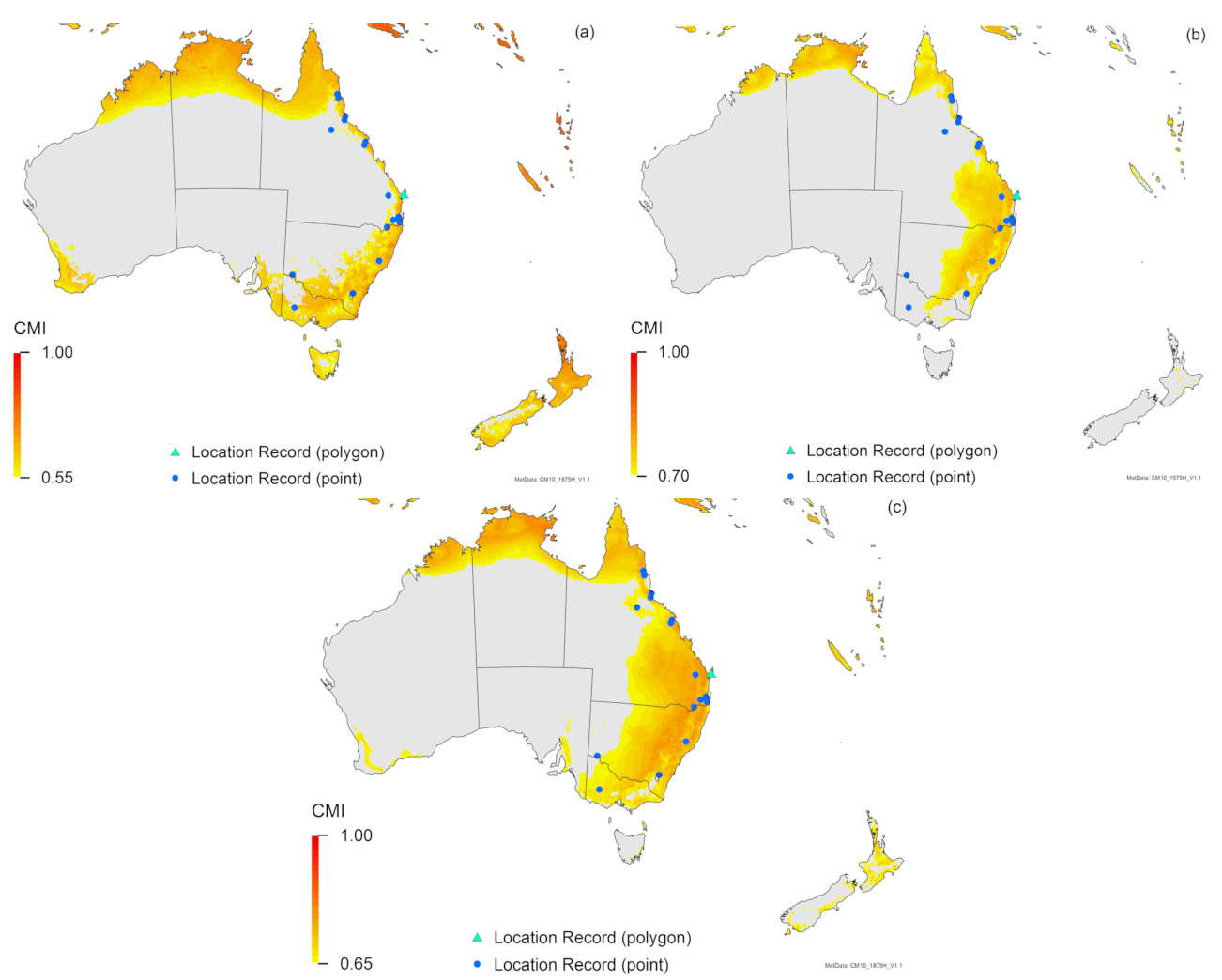
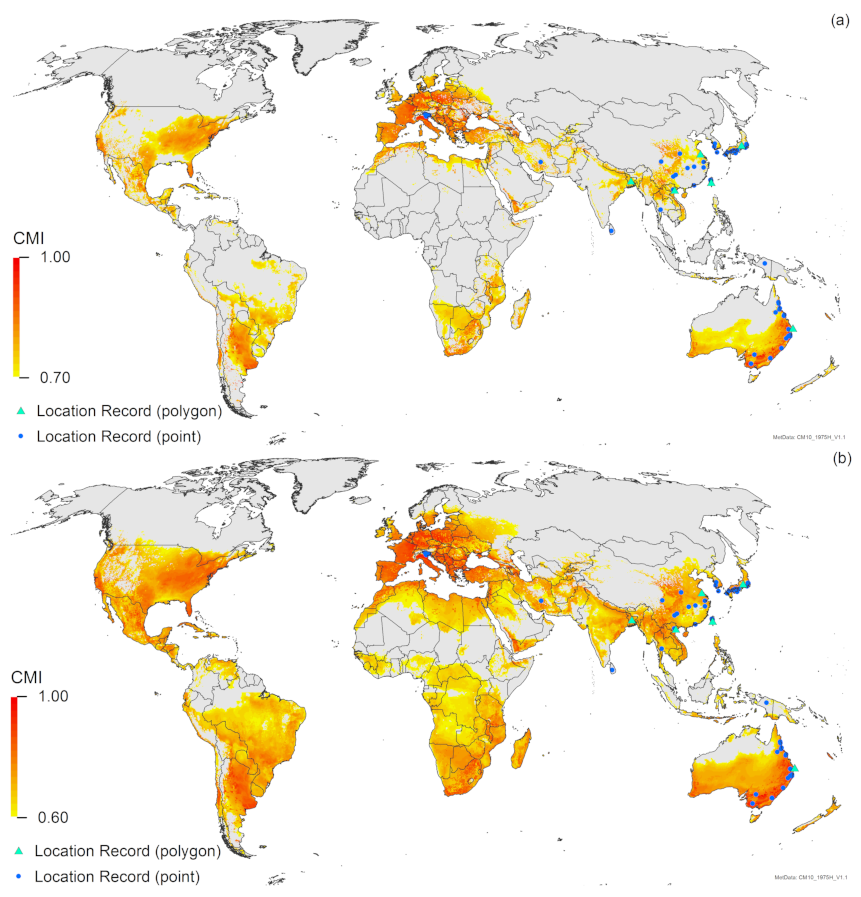
3. Results
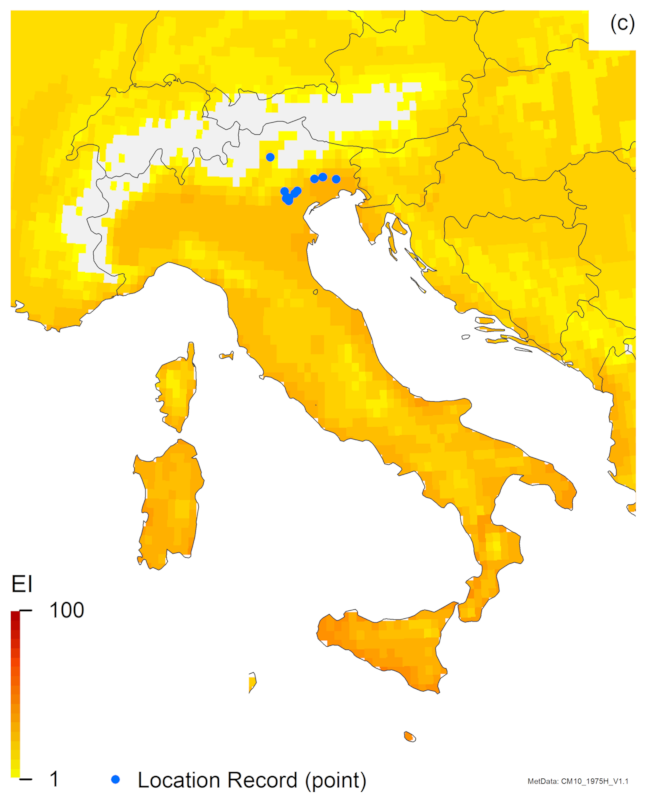
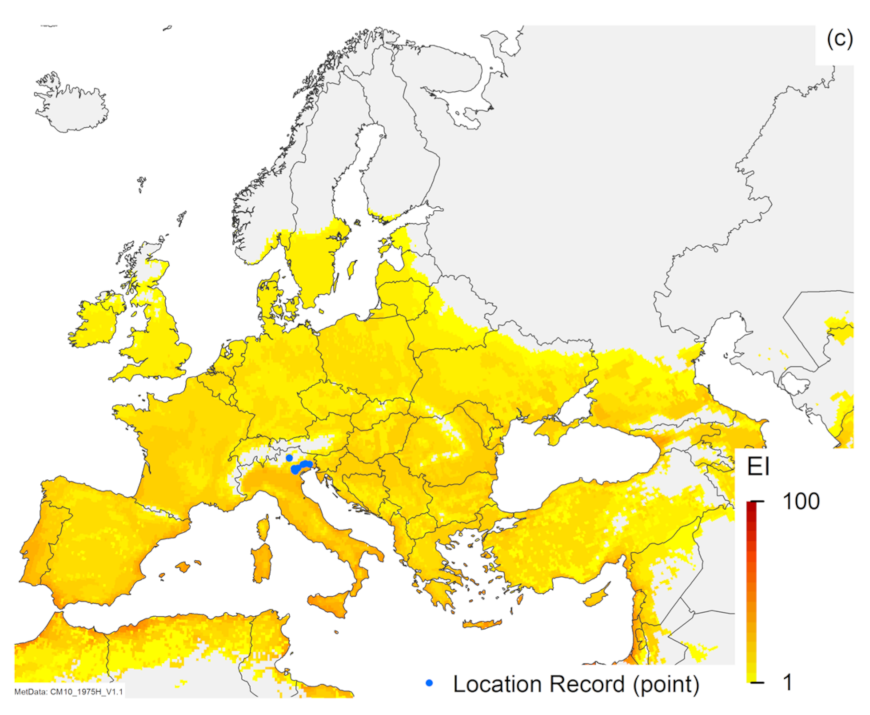
3.1. Regional Match Climates
3.2. Compare Locations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. CLIMEX Match Climates (Regional) for T. mitsukurii
Appendix A.1. Methods


Appendix A.2. Results






Appendix A.3. Discussion
References
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Gariepy, T.; Fraser, H.; Scott-Dupree, C. Brown marmorated stink bug (Hemiptera: Pentatomidae) in Canada: Recent establishment, occurrence, and pest status in southern Ontario. Can. Entomol. 2014, 146, 579–582. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integ. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Inkley, D.B. Characteristics of home invasion by the brown marmorated stink bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2012, 47, 125–130. [Google Scholar] [CrossRef]
- Arakawa, R.; Namura, Y. Effects of temperature on development of three Trissolcus spp. (Hymenoptera: Scelionidae), egg parasitoids of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Entomol. Sci. 2002, 5, 215–218. [Google Scholar]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Charles, J.; Avila, G.; Hoelmer, K.A.; Hunt, S.; Gardner-Gee, R.; MacDonald, F.; Davis, V. Experimental assessment of the biosafety of Trissolcus japonicus in New Zealand, prior to the anticipated arrival of the invasive pest Halyomorpha halys. Biocontrol 2019, 64, 367–379. [Google Scholar] [CrossRef]
- Horwood, M.; Milnes, J.M.; Cooper, W.R. Brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), detections in Western Sydney, New South Wales, Australia. Austral. Entomol. 2019, 58, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Caron, V.; Yonow, T.; Paull, C.; Talamas, E.J.; Avila, G.; Hoelmer, K. Pre-empting the arrival of the brown marmorated stink bug Halyomorpha halys—Biological control options for Australia. Insects. submitted.
- Yang, Z.-Q.; Yao, Y.-X.; Qiu, L.-F.; Li, Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Hedstrom, C.; Lowenstein, D.; Andrews, H.; Bai, B.; Wiman, N. Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest Sci. 2017, 90, 1169–1179. [Google Scholar] [CrossRef]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.; Shrewsbury, P.M. Field surveys of egg mortality and indigenous egg parasitoids of the brown marmorated stink bug, Halyomorpha halys, in ornamental nurseries in the mid-Atlantic region of the USA. J. Pest Sci. 2017, 90, 1159–1168. [Google Scholar] [CrossRef]
- Buffington, M.L.; Talamas, E.J.; Hoelmer, K.A. Team Trissolcus: Integrating taxonomy and biological control to combat the brown marmorated stink bug. Am. Entomol. 2018, 64, 224–232. [Google Scholar] [CrossRef]
- Abram, P.K.; Talamas, E.J.; Acheampong, S.; Mason, P.G.; Gariepy, T.D. First detection of the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae), in Canada. J. Hymenopt. Res. 2019, 68, 29. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi Pio, F.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.-C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Milnes, J.M.; Beers, E.H. Trissolcus japonicus (Hymenoptera: Scelionidae) causes low levels of parasitism in three North American pentatomids under field conditions. J. Insect Sci. 2019, 19, 15. [Google Scholar] [CrossRef] [Green Version]
- Lara, J.R.; Pickett, C.H.; Kamiyama, M.T.; Figueroa, S.; Romo, M.; Cabanas, C.; Bazurto, V.; Strode, V.; Briseno, K.; Lewis, M. Physiological host range of Trissolcus japonicus in relation to Halyomorpha halys and other pentatomids from California. Biocontrol 2019, 64, 513–528. [Google Scholar] [CrossRef]
- Zapponi, L.; Bon, M.C.; Fouani, J.M.; Anfora, G.; Schmidt, S.; Falagiarda, M. Assemblage of the egg parasitoids of the invasive stink bug Halyomorpha halys: Insights on plant host associations. Insects 2020, 11, 588. [Google Scholar] [CrossRef]
- Protection Authority. Decision—To Import for Release and/or Release from Containment Any New Organism with Controls under Section 38A of the Hazardous Substances and New Organisms Act 1996. Application APP203336: To Seek Pre-Approval to Release Trissolcus japonicus (the Samurai Wasp) as a Biological Control Agent for Brown Marmorated Stink Bug (Halyomorpha halys) Should It Arrive in New Zealand. 2018. Available online: https://www.epa.govt.nz/assets/FileAPI/hsno-ar/APP203336/0ed5350647/APP203336-Decision.pdf (accessed on 18 November 2020).
- Vandervoet, T.F.; Bellamy, D.E.; Anderson, D.; MacLellan, R. Trapping for early detection of the brown marmorated stink bug, Halyomorpha halys, in New Zealand. N. Z. Plant Prot. 2019, 72, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.R. The control of Nezara viridula L. with introduced egg parasitoids in Australia. A review of a ‘landmark’ example of classical biological control. Aust. J. Agric. Res. 1990, 41, 1127–1146. [Google Scholar] [CrossRef]
- Johnson, N.F. Revision of Australasian Trissolcus species (Hymenoptera: Scelionidae). Invertebr. Syst. 1991, 5, 211–239. [Google Scholar] [CrossRef]
- Scaccini, D.; Falagiarda, M.; Tortorici, F.; Martinez-Sañudo, I.; Tirello, P.; Reyes-Domínguez, Y.; Gallmetzer, A.; Tavella, L.; Zandigiacomo, P.; Duso, C. An insight into the role of Trissolcus mitsukurii as biological control agent of Halyomorpha halys in northeastern Italy. Insects 2020, 11, 306. [Google Scholar] [CrossRef]
- Andrewartha, H.G.; Birch, L.C. The Ecological Web: More on the Distribution and Abundance of Animals; University of Chicago Press: Chicago, IL, USA, 1984. [Google Scholar]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The geographic range: Size, shape, boundaries, and internal structure. Annu. Rev. Ecol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef] [Green Version]
- Woodward, F.I. Climate and Plant Distribution; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Sutherst, R.W. Pest species distribution modelling: Origins and lessons from history. Biol. Invasions 2014, 16, 239–256. [Google Scholar] [CrossRef]
- Kriticos, D.; De Barro, P.; Yonow, T.; Ota, N.; Sutherst, R. The potential geographical distribution and phenology of Bemisia tabaci Middle East/Asia Minor 1, considering irrigation and glasshouse production. Bull. Entomol. Res. 2020, 110, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zalucki, M.P.; Bao, H.; Chen, H.; Hu, Z.; Zhang, D.; Lin, Q.; Yin, F.; Wang, M.; Feng, X. Population dynamics and “outbreaks” of diamondback moth (Lepidoptera: Plutellidae) in Guangdong province, China: Climate or failure of management? J. Econ. Entomol. 2012, 105, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.T.; Rodriguez, R.; Segura, D.F.; Cladera, J.L.; Sutherst, R.W. Potential geographical distribution of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae), with emphasis on Argentina and Australia. Environ. Entomol. 2002, 31, 1009–1022. [Google Scholar] [CrossRef] [Green Version]
- Guichard, S.; Guis, H.; Tran, A.; Garros, C.; Balenghien, T.; Kriticos, D.J. Worldwide niche and future potential distribution of Culicoides imicola, a major vector of bluetongue and African horse sickness viruses. PLoS ONE 2014, 9, e112491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olfert, O.; Weiss, R. Impact of climate change on potential distributions and relative abundances of Oulema melanopus, Meligethes viridescens and Ceutorhynchus obstrictus in Canada. Agric. Ecosyst. Environ. 2006, 113, 295–301. [Google Scholar] [CrossRef]
- Olfert, O.; Weiss, R.; Cárcamo, H.; Meers, S. The influence of abiotic factors on an invasive pest of pulse crops, Sitona lineatus (L.) (Coleoptera: Curculionidae), in North America. Psyche 2012. [Google Scholar] [CrossRef] [Green Version]
- Stoeckli, S.; Felber, R.; Haye, T. Current distribution and voltinism of the brown marmorated stink bug, Halyomorpha halys, in Switzerland and its response to climate change using a high-resolution CLIMEX model. Int. J. Biometeorol. 2020, 64, 2019–2032. [Google Scholar] [CrossRef]
- Avila, G.A.; Charles, J.G. Modelling the potential geographic distribution of Trissolcus japonicus: A biological control agent of the brown marmorated stink bug, Halyomorpha halys. Biocontrol 2018, 63, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Bomford, M.; Kraus, F.; Barry, S.C.; Lawrence, E. Predicting establishment success for alien reptiles and amphibians: A role for climate matching. Biol. Invasions 2009, 11, 713–724. [Google Scholar] [CrossRef]
- Boag, B.; Evans, K.; Yeates, G.; Johns, P.; Neilson, R. Assessment of the global potential distribution of the predatory land planarian Artioposthia triangulata (Dendy) (Tricladida: Terricola) from ecoclimatic data. N. Z. J. Zool. 1995, 22, 311–318. [Google Scholar] [CrossRef]
- Robertson, M.P.; Kriticos, D.J.; Zachariades, C. Climate matching techniques to narrow the search for biological control agents. Biol. Control 2008, 46, 442–452. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R.W. CLIMEX Version 4: Exploring the Effects of Climate on Plants, Animals and Diseases; CSIRO: Canberra, Australia, 2015. [Google Scholar]
- Phillips, C.B.; Kean, J.M.; Vink, C.J.; Berry, J.A. Utility of the CLIMEX ‘match climates regional’ algorithm for pest risk analysis: An evaluation with non-native ants in New Zealand. Biol. Invasions 2018, 20, 777–791. [Google Scholar] [CrossRef]
- Csurhes, S.; Kriticos, D. Gleditsia triacanthos L. (Caesalpiniaceae), another thorny, exotic fodder tree gone wild. Plant Prot. Q. 1994, 9, 101. [Google Scholar]
- De Villiers, M.; Hattingh, V.; Kriticos, D.J.; Brunel, S.; Vayssières, J.-F.; Sinzogan, A.; Billah, M.; Mohamed, S.; Mwatawala, M.; Abdelgader, H. The potential distribution of Bactrocera dorsalis: Considering phenology and irrigation patterns. Bull. Entomol. Res. 2016, 106, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Moraglio, S.T.; Tortorici, F.; Giromini, D.; Pansa, M.G.; Visentin, S.; Tavella, L. Field collection of egg parasitoids of Pentatomidae and Scutelleridae in Northwest Italy and their efficacy in parasitizing Halyomorpha halys under laboratory conditions. Entomol. Exp. Appl. 2020, 169, 52–63. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J. Pest Sci. 2020, 93, 183–194. [Google Scholar] [CrossRef]
- Haye, T.; Moraglio, S.T.; Stahl, J.; Visentin, S.; Gregorio, T.; Tavella, L. Fundamental host range of Trissolcus japonicus in Europe. J. Pest Sci. 2020, 93, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini-Peverieri, G.; Dieckhoff, C.; Giovannini, L.; Marianelli, L.; Roversi, P.F.; Hoelmer, K. Rearing Trissolcus japonicus and Trissolcus mitsukurii for biological control of Halyomorpha halys. Insects 2020, 11, 787. [Google Scholar] [CrossRef]
- Hymenoptera Online (HOL). USA Ohio State University. 2021. Available online: https://mbd-db.osu.edu/hol/ (accessed on 13 March 2019).
- Talamas, E.J.; (Florida Department of Agriculture and Consumer Services, Gainesville, FL, USA). Personal communication, 2020.
- Chen, H.; (State Key Laboratory of Biocontrol, School of Life Sciences/School of Ecology, Sun Yat-sen University, Guangzhou 510275, China). Personal communication, 2019.
- Atlas of Living Australia. Available online: https://bie.ala.org.au/species/urn:lsid:biodiversity.org.au:afd.taxon:592af144-bea0-4612-a3a3-dbdcf71ea4f6 (accessed on 10 March 2021).
- Mohammadpour, M.; Ziaaddini, M.; Jalali, M.A.; Hashemirad, H.; Mohammadi-Khoramabadi, A. Egg parasitoids of the pistachio green stink bug, Brachynema germari (Hemiptera: Pentatomidae) in Kerman province, Iran. Zool. Ecol. 2016, 26, 28–34. [Google Scholar] [CrossRef]
- Davis, C.; Krauss, N. Recent Introductions for Biological Control in Hawaii-XII. Proc. Hawaii Entomol. Soc. 1967, 19, 375–380. [Google Scholar]
- Gariepy, T.D.; (Agriculture and Agri-Food Canada, London, ON, Canada). Personal communication, 2019.
- Avila, G.A.; (The New Zealand Institute for Plant & Food Research, Auckland, New Zealand). Personal communication, 2019.
- Global Biodiversity Information Facility. GBIF.org, GBIF Occurrence. Available online: https://doi.org/10.15468/dl.w2sym7 (accessed on 24 August 2020).
- EPPO Global Database. Halyomorpha halys. Available online: https://gd.eppo.int/taxon/HALYHA/distribution (accessed on 16 October 2020).
- Avila, G.A.; (The New Zealand Institute for Plant & Food Research, Auckland, New Zealand). Personal communication, 2020.
- Boberg, J.; (Swedish University of Agricultural Sciences, Uppsala, Sweden). Personal communication, 2020.
- Stevens, L.; (Department for Environment, Food and Rural Affairs, York, UK). Personal communication, 2020.
- Gariepy, T.D.; (Agriculture and Agri-Food Canada, London, ON, Canada). Personal communication, 2020.
- Abram, P.K.; Hueppelsheuser, T.; Acheampong, S.; Clarke, P.; Douglas, H.; Gariepy, T.D. Evidence of established brown marmorated stink bug populations in British Columbia, Canada. J. Entomol. Soc. B.C. 2017, 114, 83–86. [Google Scholar]
- Chouinard, G.; Larose, M.; Légaré, J.-P.; Bourgeois, G.; Racette, G.; Barrette, M. Interceptions and captures of Halyomorpha halys (Hemiptera: Pentatomidae) in Quebec from 2008 to 2018. Phytoprotection 2018, 98, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Cianferoni, F.; Graziani, F.; Dioli, P.; Ceccolini, F. Review of the occurrence of Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae) in Italy, with an update of its European and world distribution. Biologia 2018, 73, 599–607. [Google Scholar] [CrossRef]
- Konjević, A. First records of the brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Hemiptera: Pentatomidae) in Republic of North Macedonia. Acta. Zool. Bulg. 2020, 72, 687–690. [Google Scholar]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Roigé, M.; Phillips, C.B. Validation and uncertainty analysis of the match climates regional algorithm (CLIMEX) for Pest risk analysis. Ecol. Inform. 2020, 101196. [Google Scholar] [CrossRef]
- Kriticos, D.J. Regional climate-matching to estimate current and future sources of biosecurity threats. Biol. Invasions 2012, 14, 1533–1544. [Google Scholar] [CrossRef]
- Arakawa, R.; Miura, M.; Fujita, M. Effects of host species on the body size, fecundity, and longevity of Trissolcus mitsukurii (Hymenoptera: Scelionidae), a solitary egg parasitoid of stink bugs. Appl. Entomol. Zool. 2004, 39, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Kriticos, D.; Sutherst, R.; Brown, J.; Adkins, S.; Maywald, G. Climate change and biotic invasions: A case history of a tropical woody vine. Biol. Invasions 2003, 5, 147–165. [Google Scholar] [CrossRef]
- Kobayashi, T.; Cosenza, G.W. Integrated control of soybean stink bugs in the Cerrados. JARQ 1987, 20, 229–236. [Google Scholar]
- Yonow, T.; Kriticos, D.J.; Ota, N. The potential distribution of cassava mealybug (Phenacoccus manihoti), a threat to food security for the poor. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Yonow, T.; Kriticos, D.J.; Ota, N.; Van Den Berg, J.; Hutchison, W.D. The potential global distribution of Chilo partellus, including consideration of irrigation and cropping patterns. J. Pest Sci. 2017, 90, 459–477. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.; Henrich, V.; Frenken, K.; Burke, J. Global Map of Irrigation Areas version 5; Rheinische Friedrich-Wilhelms-University: Bonn, Germany; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Yonow, T.; Ramirez-Villegas, J.; Abadie, C.; Darnell, R.E.; Ota, N.; Kriticos, D.J. Black Sigatoka in bananas: Ecoclimatic suitability and disease pressure assessments. PLoS ONE 2019, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R. Assessing the distribution of exotic egg parasitoids of Halyomorpha halys in Europe with a large-scale monitoring program. Insects 2021, 12, 316. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Kriticos, D.J.; Leriche, A. The effects of climate data precision on fitting and projecting species niche models. Ecography 2010, 33, 115–127. [Google Scholar] [CrossRef]
- Kiritani, K. The impact of global warming and land-use change on the pest status of rice and fruit bugs (Heteroptera) in Japan. Glob. Change Biol. 2007, 13, 1586–1595. [Google Scholar] [CrossRef]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Wiman, N.G.; Walton, V.M.; Shearer, P.W.; Rondon, S.I.; Lee, J.C. Factors affecting flight capacity of brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). J. Pest Sci. 2015, 88, 37–47. [Google Scholar] [CrossRef]
- Lee, D.-H.; Leskey, T. Flight behavior of foraging and overwintering brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Bull. Entomol. Res. 2015, 105, 566–573. [Google Scholar] [CrossRef] [Green Version]
- James, D.G. Energy reserves, reproductive status and population biology of overwintering Biprorulus bibax (Hemiptera, Pentatomidae) in southern New South Wales citrus groves. Aust. J. Zool. 1990, 38, 415–422. [Google Scholar] [CrossRef]
- Van der Ploeg, R.R.; Böhm, W.; Kirkham, M.B. On the origin of the theory of mineral nutrition of plants and the law of the minimum. Soil Sci. Soc. Am. J. 1999, 63, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Shelford, V.E. The Ecology of North America; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | T. mitsukurii | T. japonicus | H. halys |
|---|---|---|---|---|
| Moisture | ||||
| SM0 | lower soil moisture threshold | 0.1 | 0.1 | 0.1 |
| SM1 | lower optimum soil moisture | 0.5 | 0.4 | 0.5 |
| SM2 | upper optimum soil moisture | 1.6 | 1.2 | 1 |
| SM3 | upper soil moisture threshold | 1.8 | 1.6 | 1.5 |
| Temperature | ||||
| DV0 | lower temperature threshold | 12 °C | 12 °C | 12 °C |
| DV1 | lower optimum temperature | 25 °C | 27 °C | 27 °C |
| DV2 | upper optimum temperature | 30 °C | 30 °C | 30 °C |
| DV3 | upper temperature threshold | 32 °C | 34 °C | 33 °C |
| Diapause | ||||
| DPD0 | diapause induction daylength | 12 h light | ||
| DPT0 | diapause induction temperature | 5 °C | ||
| DPT1 | diapause termination temperature | 5 °C | ||
| DPD | diapause development days | 0 | ||
| DPSW | diapause summer (1) or winter (0) | 0 | ||
| Cold Stress | ||||
| TTCS | cold stress temperature threshold | −7 °C | −18.3 °C | −18 °C |
| THCS | temperature threshold stress accumulation rate | −0.01 week−1 | −0.0015 week−1 | −0.01 week−1 |
| DTCS | degree-day cold stress threshold | |||
| DHCS | degree-day cold stress accumulation rate | |||
| Heat Stress | ||||
| TTHS | heat stress temperature threshold | 34 °C | 33 °C | |
| THHS | temperature threshold stress accumulation rate | 0.055 week−1 | 0.01 week−1 | |
| DTHS | degree-day heat stress threshold | 4 | ||
| DHHS | degree-day heat stress accumulation rate | 0.0008 week−1 | ||
| Dry Stress | ||||
| SMDS | soil moisture dry stress threshold | 0.1 | 0.1 | 0.1 |
| HDS | stress accumulation rate | −0.01 week−1 | −0.01 week−1 | −0.01 week−1 |
| Wet Stress | ||||
| SMWS | soil moisture wet stress threshold | 1.8 | 1.6 | 1.5 |
| HWS | stress accumulation rate | 0.01 week−1 | 0.0065 week−1 | 0.002 week−1 |
| Hot Wet Stress | ||||
| TTHW | hot wet stress threshold temperature | 27 °C | 28 °C | |
| MTHW | hot wet stress threshold soil moisture | 1.4 | 1.5 | |
| PHW | hot wet stress accumulation rate | 0.0024 week−1 | 0.007 week−1 | |
| Threshold Heat Sum | ||||
| PDD | number of degree-days above DV0 needed to complete one generation | 185 °C-days | 175 °C-days | 595 °C-days |
| Irrigation Scenario | 2.5 mm week−1 as top up | all models run with this irrigation scenario in this paper | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yonow, T.; Kriticos, D.J.; Ota, N.; Avila, G.A.; Hoelmer, K.A.; Chen, H.; Caron, V. Modelling the Potential Geographic Distribution of Two Trissolcus Species for the Brown Marmorated Stink Bug, Halyomorpha halys. Insects 2021, 12, 491. https://doi.org/10.3390/insects12060491
Yonow T, Kriticos DJ, Ota N, Avila GA, Hoelmer KA, Chen H, Caron V. Modelling the Potential Geographic Distribution of Two Trissolcus Species for the Brown Marmorated Stink Bug, Halyomorpha halys. Insects. 2021; 12(6):491. https://doi.org/10.3390/insects12060491
Chicago/Turabian StyleYonow, Tania, Darren J. Kriticos, Noboru Ota, Gonzalo A. Avila, Kim A. Hoelmer, Huayan Chen, and Valerie Caron. 2021. "Modelling the Potential Geographic Distribution of Two Trissolcus Species for the Brown Marmorated Stink Bug, Halyomorpha halys" Insects 12, no. 6: 491. https://doi.org/10.3390/insects12060491