
Development of Sterile Insect Technique for Control of the European Grapevine Moth, Lobesia botrana, in Urban Areas of Chile

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Development of Mass Rearing Program
2.1. Development of Larval Rearing Diets
2.2. Oviposition Cages
2.3. Larval Rearing
2.4. Moth Collection
2.5. Irradiation
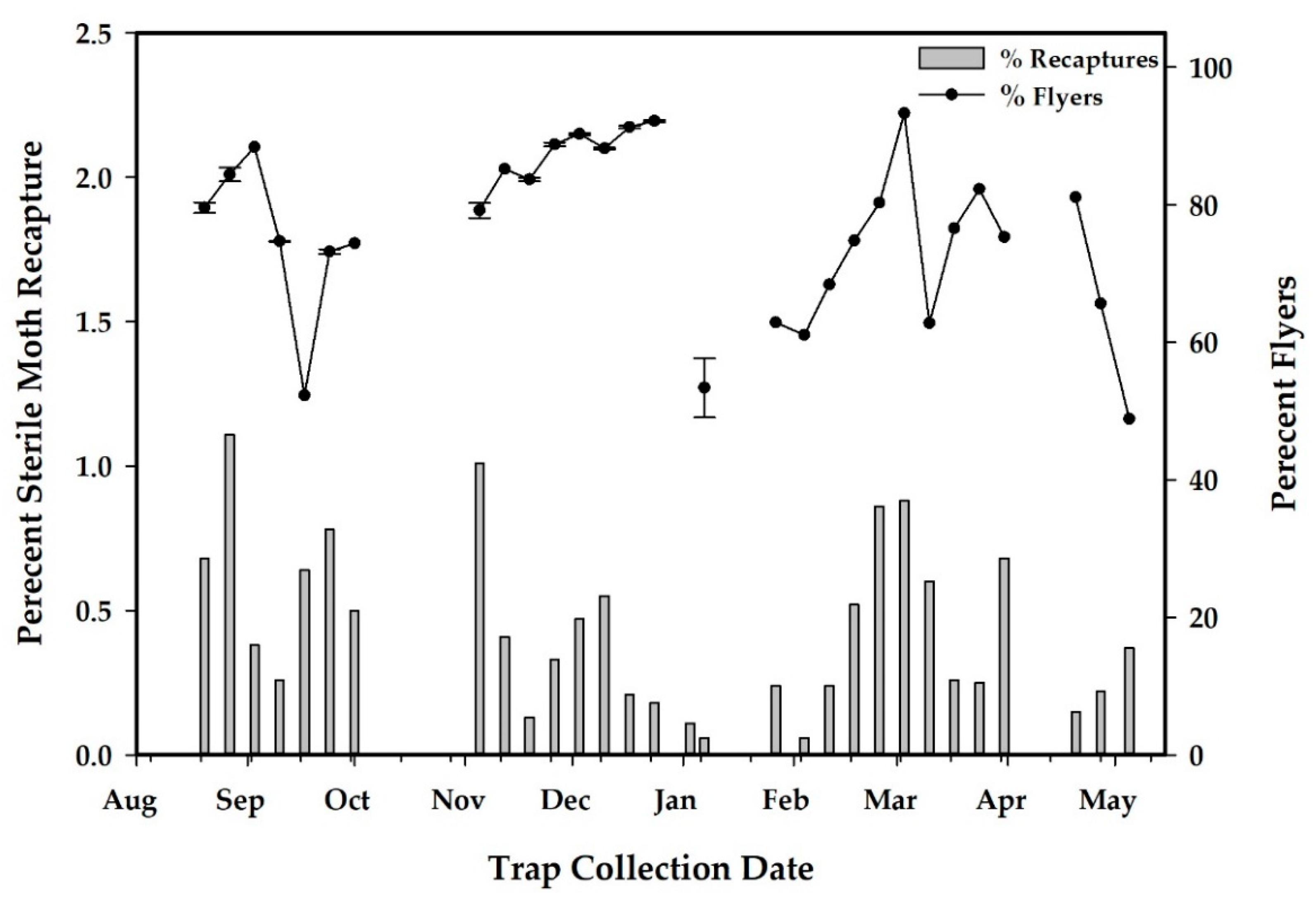
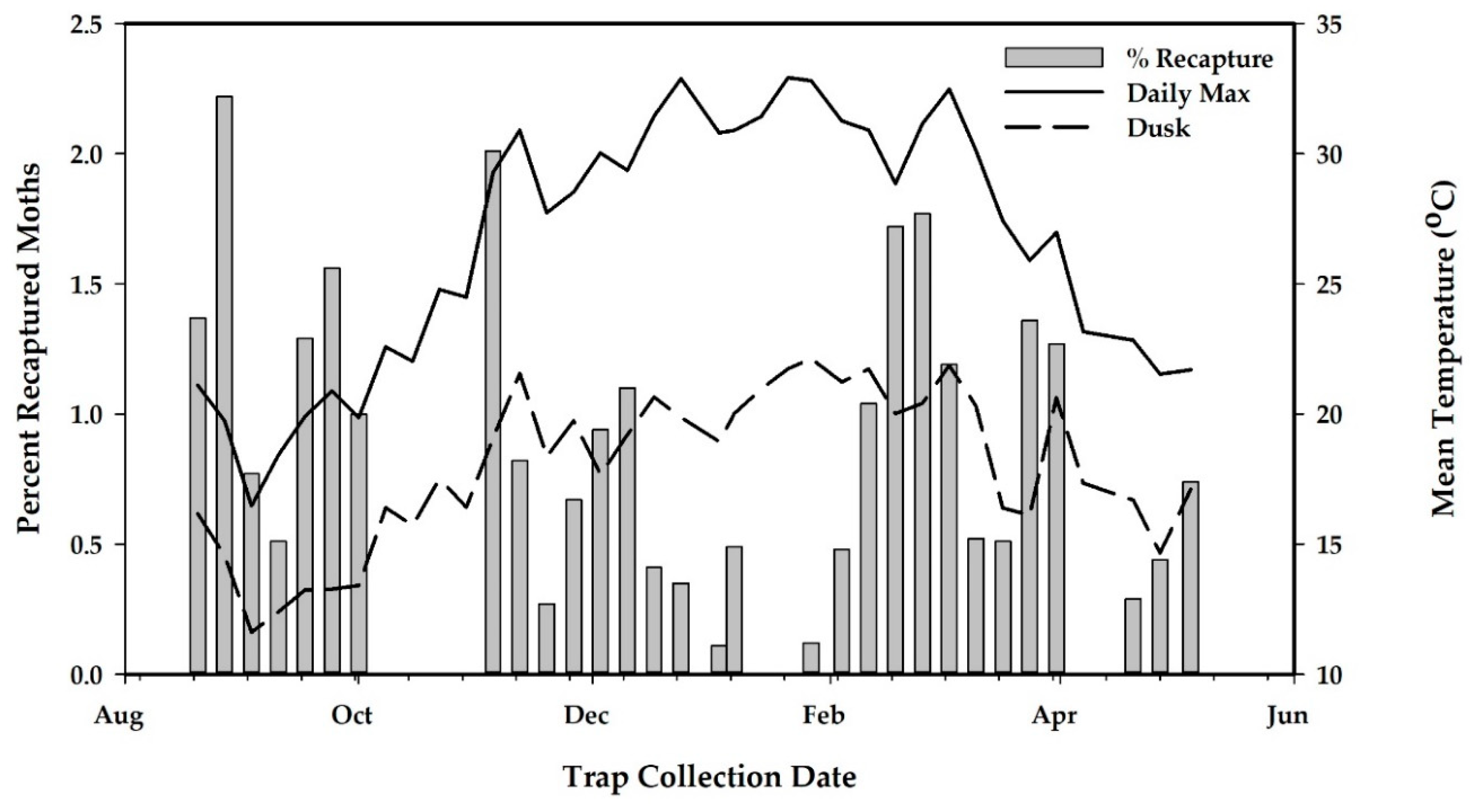
2.6. Quality Control
3. Field Release Studies
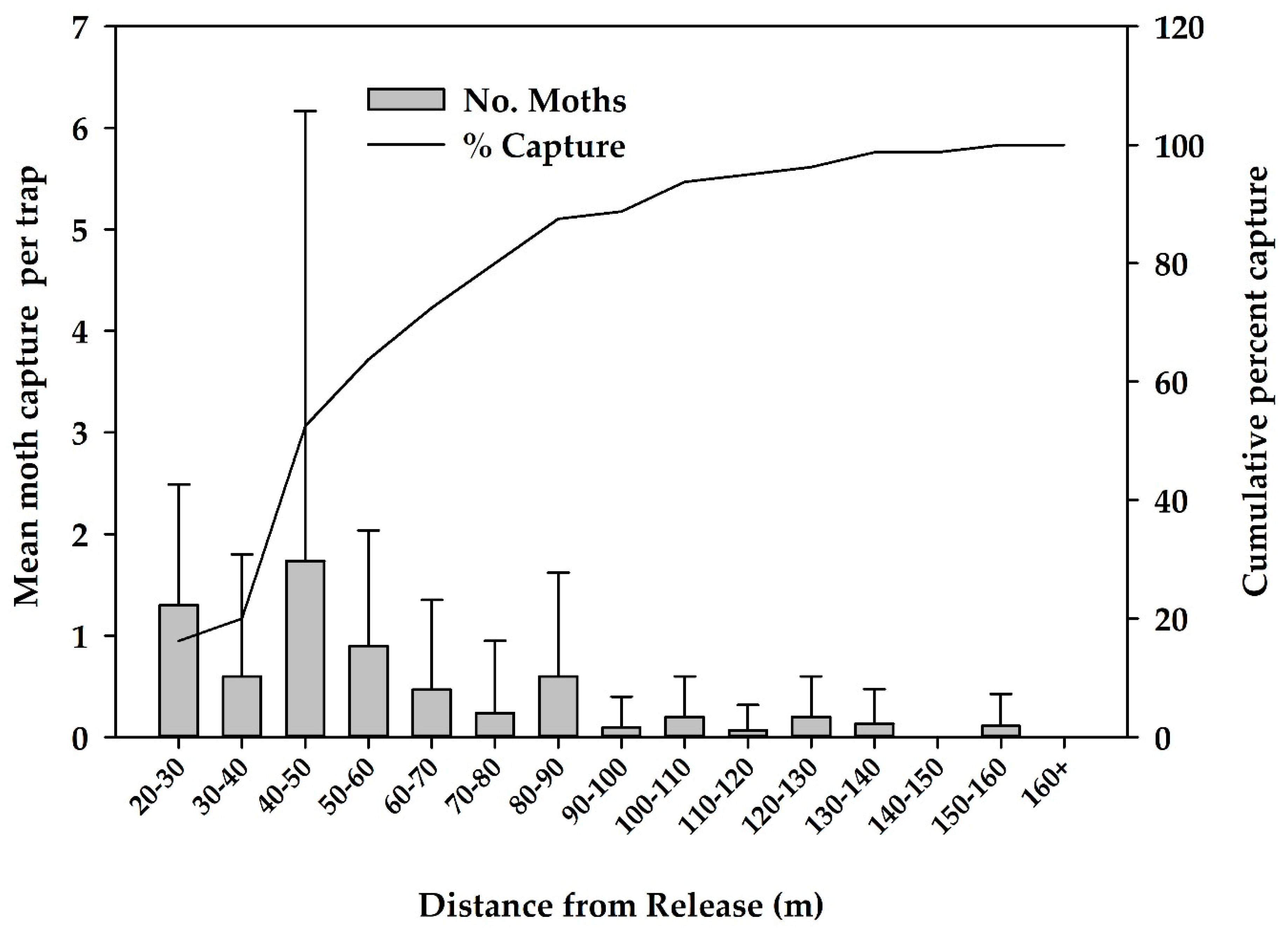
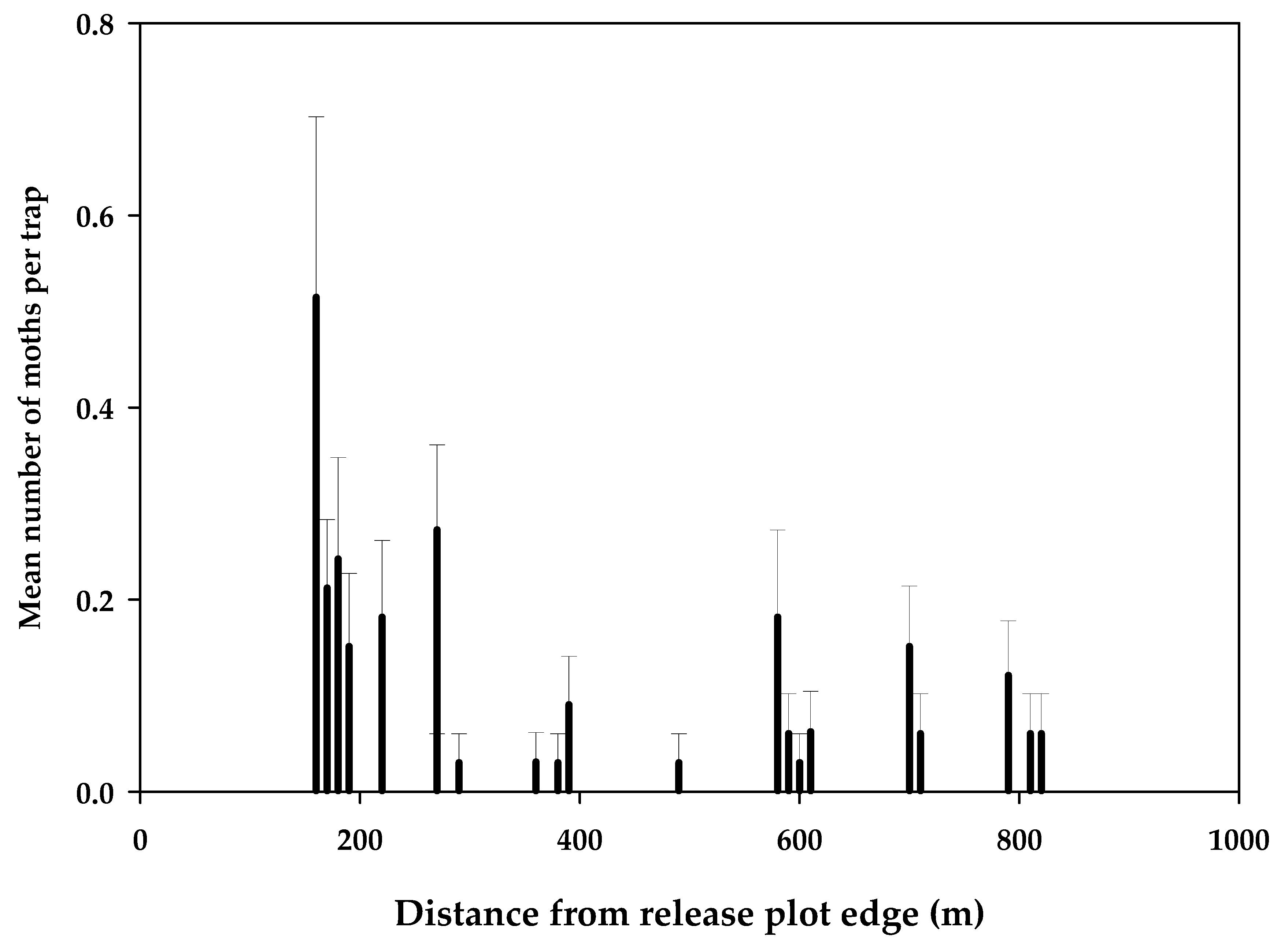
3.1. Dispersal Studies in 2018–2019
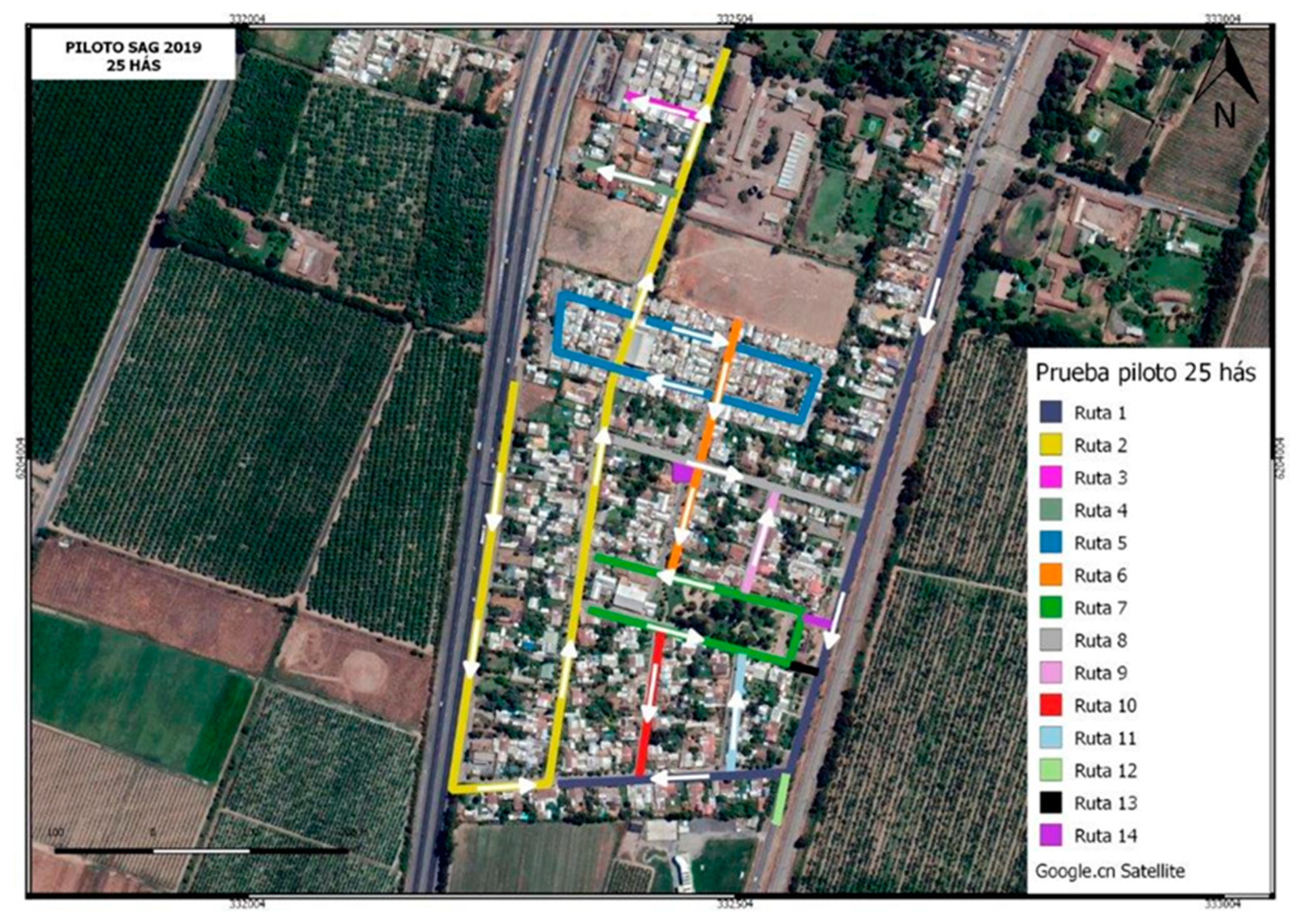
3.2. Pilot Project Field Release Trials in 2019–2020
3.2.1. Sterile Moth Releases in Pilot Project
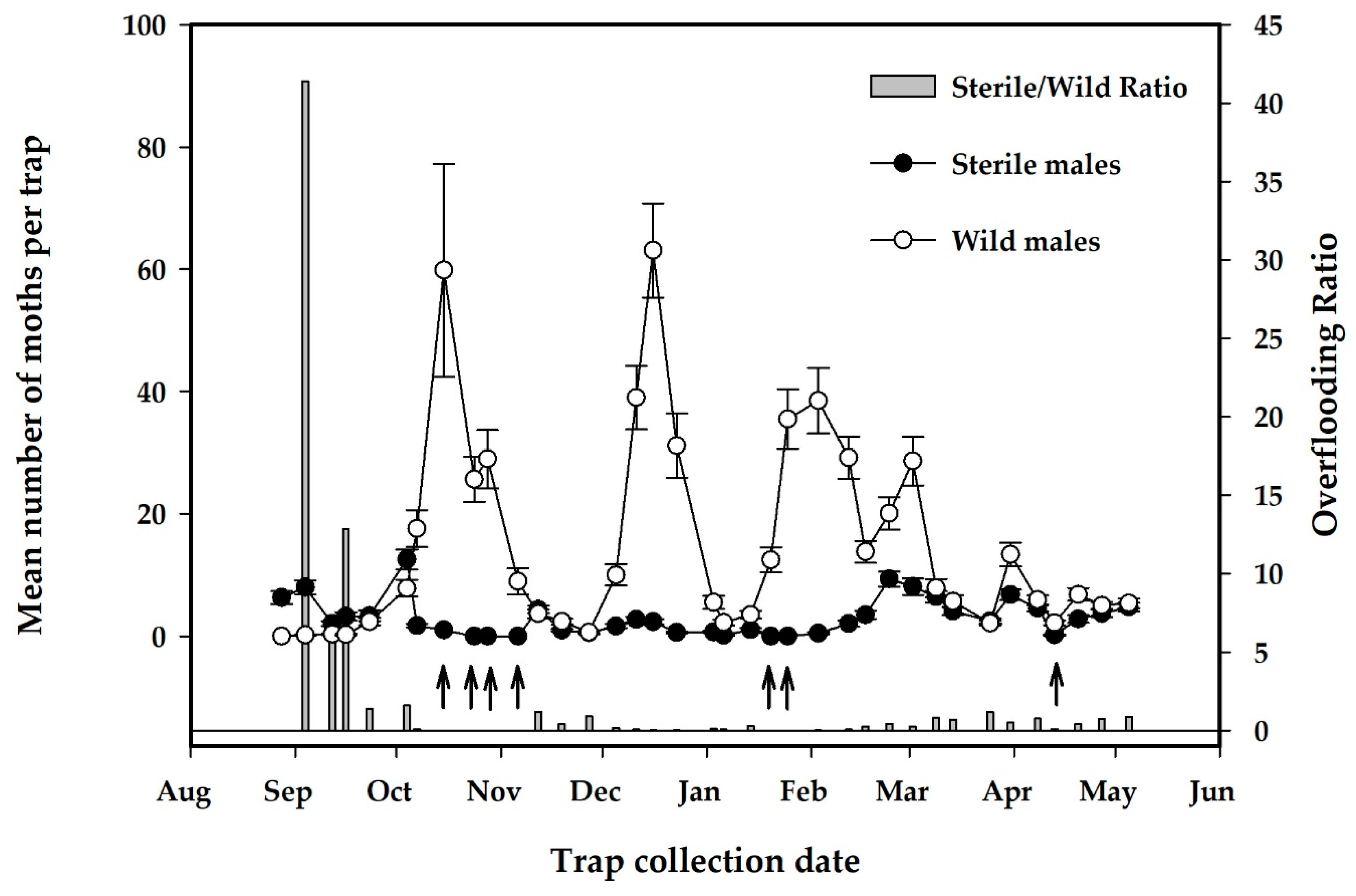
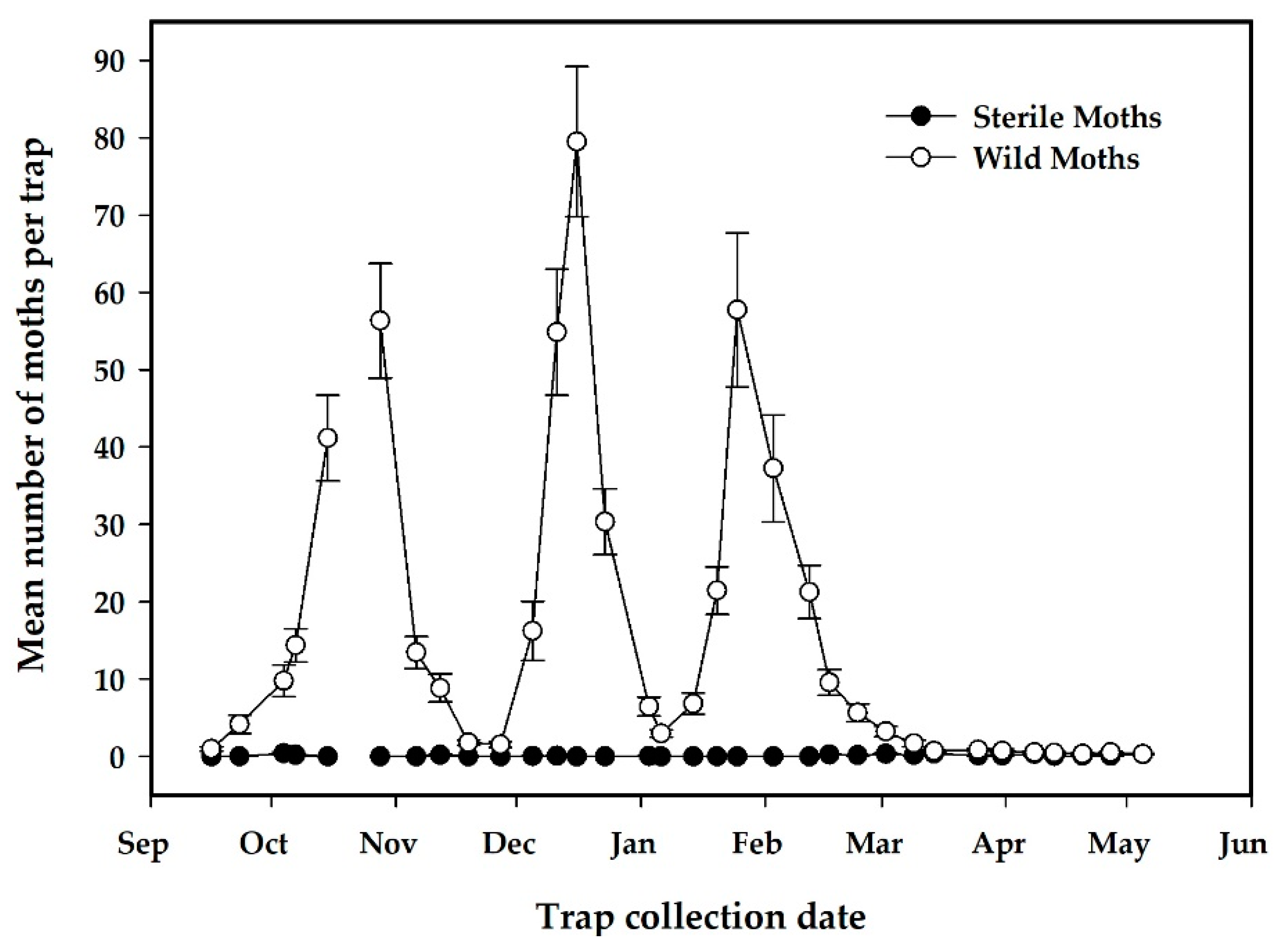
3.2.2. Results of Sterile Moth Releases in Pilot Project
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical Ecology and Management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Èntomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A.; Varela, L.G. Grape Berry Moths in Western European Vineyards and Their Recent Movement into the New World. In Arthropod Management in Vineyards; Springer: Berlin/Heidelberg, Germany, 2012; pp. 339–359. [Google Scholar]
- Anfora, G.; Tasin, M.; De Cristofaro, A.; Ioriatti, C.; Lucchi, A. Synthetic Grape Volatiles Attract Mated Lobesia botrana Females in Laboratory and Field Bioassays. J. Chem. Ecol. 2009, 35, 1054–1062. [Google Scholar] [CrossRef]
- Cooper, M.; Varela, L.; Smith, R.; Whitmer, D.; Simmons, G.; Lucchi, A.; Broadway, R.; Steinhauer, R. Managing newly established pests: Growers, scientists and regulators collaborate on European grapevine moth program. Calif. Agric. 2014, 68, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.P.; Ponti, L.; Cooper, M.L.; Gilioli, G.; Baumgartner, J.; Duso, C. Prospective analysis of the invasive potential of the European grapevine moth Lobesia botrana (Den. & Schiff.) in California. Agric. Èntomol. 2012, 14, 225–238. [Google Scholar] [CrossRef]
- Fermaud, M.; Le Menn, R. Transmission of Botrytis cinerea to grapes by grape berry moth larvae. Phytopathology 1992, 82, 1393–1398. [Google Scholar] [CrossRef]
- FA0-IAEA. The Sterile Insect Technique for Use against the Devastating European Grapevine Moth in Chile. June 2020. Available online: http://www.fao.org/3/ca9542en/ca9542en.pdf (accessed on 30 December 2020).
- SAG. Declara Control Obligatorio de la Plaga Polilla del Racimo de la vid (Lobesia botrana) 11 May 2013. Available online: https://normativa.sag.gob.cl/Publico/Normas/DetalleNorma.aspx?id=1055986 (accessed on 16 April 2021).
- SAG. Ministro de Agricultura y Director Nacional del SAG Fiscalizaron Predios en el Marco del Programa de Control por Lobesia botrana 2014. Available online: http://www.sag.gob.cl/noticias/ministro-de-agricultura-y-director-nacional-del-sag-fiscalizaron-predios-en-el-marco-del (accessed on 16 April 2021).
- Addante, R.; Moleas, T. Effectiveness of mating disruption method against Lobesia botrana (Den et Schiff.) (Lepidoptera: Tortricidea) in Apulian vineyards. Acta Hortic. 1995, 422, 247–251. [Google Scholar] [CrossRef]
- USDA-APHIS. Commodity Import Report (CIR) Blueberry (Fruit) from Chile into All Ports. 2021. Available online: https://epermits.aphis.usda.gov/manual/index.cfm?action=cirReportP&PERMITTED_ID=4741 (accessed on 17 March 2021).
- CFIA. Phytosanitary Import Requirements to Prevent the Introduction of Lobesia Botrana, the European Grapevine Moth. 2018. Available online: https://members.wto.org/crnattachments/2017/SPS/CAN/17_5016_00_e.pdf (accessed on 16 April 2021).
- FAO/IAEA. Developing Area-Wide SIT Against the European Grape Vine Moth in Chile. 2018. Available online: http://www-naweb.iaea.org/nafa/news/2018-developing-area-wide-SIT-chile.html (accessed on 11 February 2021).
- SAG. Se Lanza Temporada 2015–2016 Contra Lobesia Botrana en la Región de O’Higgins. 2015. Available online: http://www.sag.gob.cl/noticias/se-lanza-temporada-2015-2016-contra-lobesia-botrana-en-la-region-de-ohiggins (accessed on 16 April 2021).
- Simmons, G.S.; Bloem, K.A.; Bloem, S.; Carpenter, J.E.; Suckling, D.M. Impact of Moth Suppression/Eradication Programmes Using the Sterile Insect Technique or Inherited Sterility. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; IAEA. CRC Press: Boca Raton, FL, USA, 2021; pp. 1007–1050. [Google Scholar]
- Dyck, V.A. Rearing Codling Moth for the Sterile Insect Technique; FAO: Rome, Italy, 2010; Volume 199, p. 197. [Google Scholar]
- Miller, E.; Stewart, F.; Anna, L.; John, B. New method of processing diet for mass rearing pink bollworm, P. gossypiella Saunders (Lepidoptera: Gelichiidae). J. Agric. Entomol. 1996, 13, 129–137. [Google Scholar]
- Tzanakakis, Μ.Ε.; Savopoulou-Soultani, Μ.; Oustapassidis, C.S.; Verras, S.; Hatziemmanouel, H. Induction of dormancy in Lobesia botrana by long day and high temperature conditions. Èntomol. Hell. 2017, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Saour, G. Sterile insect technique and F1 sterility in the European grapevine moth, Lobesia botrana. J. Insect Sci. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchi, A.; Sambado, P.; Royo, A.B.J.; Bagnoli, B.; Benelli, G. Lobesia botrana males mainly fly at dusk: Video camera-assisted pheromone traps and implications for mating disruption. J. Pest Sci. 2018, 91, 1327–1334. [Google Scholar] [CrossRef]
- Avidov, Z.; Harpaz, I. Plant Pests of Israel; Israel Universities Press: Jerusalem, Israel, 1969; pp. 380–384. [Google Scholar]
- Sunrise, Sunset, Dawn and Dusk Times around the World. Available online: https://www.gaisma.com/en (accessed on 15 January 2021).
- Steinitz, H.; Sadeh, A.; Kliot, A.; Harari, A. Effects of radiation on inherited sterility in the European grapevine moth (Lobesia botrana). Pest Manag. Sci. 2014, 71, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Marec, F.; Bloem, S.; Carpenter, J. Inherited Sterility in Insects, in Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Hendrichs, D.V.A.J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Marec, F.; Vreysen, M.J.B. Advances and Challenges of Using the Sterile Insect Technique for the Management of Pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloem, S.; Carpenter, J.E. Evaluation of Population Suppression by Irradiated Lepidoptera and Their Progeny. Fla. Èntomol. 2001, 84, 165. [Google Scholar] [CrossRef]
- Barclay, H.J. Modeling incomplete sterility in a sterile release program: Interactions with other factors. Popul. Ecol. 2001, 43, 197–206. [Google Scholar] [CrossRef]
- Kean, J.M.; Wee, S.L.; Stephens, A.E.A.; Suckling, D.M. Modelling the effects of inherited sterility for the application of the sterile insect technique. Agric. Èntomol. 2008, 10, 101–110. [Google Scholar] [CrossRef]
- Saour, G. Flight Ability and Dispersal of European Grapevine Moth Gamma-Irradiated Males (Lepidoptera: Tortricidae). Fla. Èntomol. 2016, 99, 73–78. [Google Scholar] [CrossRef]
- Boersma, N.; Carpenter, J.E. Influence of Holding Temperature and Irradiation on Field Performance of Mass-Reared Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Fla. Èntomol. 2016, 99, 215–221. [Google Scholar] [CrossRef]
- Bloem, S.; Knight, A.L. Assessing the Quality of Mass-Reared Codling Moths (Lepidoptera: Tortricidae) by Using Field Release-Recapture Tests. J. Econ. Èntomol. 1998, 91, 1122–1130. [Google Scholar] [CrossRef]
- Miller, E.; Lowe, A.; Archuleta, S. Evaluation of different release strategies for use in pink bollworm sterile release programs. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–8 January 2000; Dugger, P., Richter, D.A., Eds.; National Cotton Council: Memphis, TN, USA, 2000; pp. 1368–1370. [Google Scholar]
- FAO/IAEA. Guideline for Packing, Shipping, Holding and Release of Sterile Flies in Area-Wide Fruit Fly Control Programmes; Zavala-López, J.L., Enkerlin, W.R., Eds.; FAO: Rome, Italy, 2017; p. 140. [Google Scholar]
- Esch, E.D.; Horner, R.M.; Krompetz, D.C.; Moses-Gonzales, N.; Tesche, M.R.; Suckling, D.M. Operational Parameters for the Aerial Release of Sterile Codling Moths Using an Uncrewed Aircraft System. Insects 2021, 12, 159. [Google Scholar] [CrossRef]
- Hight, S.D.; Carpenter, J.E.; Bloem, S.; Bloem, K.A. Developing a Sterile Insect Release Program for Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae): Effective Overflooding Ratios and Release-Recapture Field Studies. Env. Èntomol. 2005, 34, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Hofmeyr, J.H.; Carpenter, J.E.; Bloem, S. Developing the Sterile Insect Technique for Cryptophlebia leucotreta (Lepidoptera: Tortricidae): Influence of Radiation Dose and Release Ratio on Fruit Damage and Population Growth in Field Cages. J. Econ. Entomol. 2005, 98, 1924–1929. [Google Scholar] [CrossRef] [PubMed]
- Kean, J.M.; Stephens, A.E.A.; Wee, S.L.; Suckling, D.M. Optimizing Strategies for Eradication of Discrete-Generation Lepidopteran Pests Using Inherited Sterility. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 211–220. [Google Scholar]
- Kean, J.M.; Suckling, D.M.; Stringer, L.D.; Woods, B. Modeling the sterile insect technique for suppression of light brown apple moth (Lepidoptera: Tortricidae). J. Econ. Èntomol. 2011, 104, 1462–1475. [Google Scholar] [CrossRef] [PubMed]
- Bloem, S.; Bloem, K.A.; Carpenter, J.E.; Calkins, C.O. Inherited Sterility in Codling Moth (Lepidoptera: Tortricidae): Effect of Substerilizing Doses of Radiation on Insect Fecundity, Fertility, and Control. Ann. Èntomol. Soc. Am. 1999, 92, 222–229. [Google Scholar] [CrossRef]
- Barclay, H.J. Mathematical Models for Using Sterile Insects, in Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 201–244. [Google Scholar]
- Cagnotti, C.L.; Andorno, A.V.; Paladino, L.C.; Botto, E.N.; López, S.N.; Hernández, C.M. Inherited Sterility in Tuta absoluta (Lepidoptera: Gelechiidae): Pest Population Suppression and Potential for Combined Use with a Generalist Predator. Fla. Èntomol. 2016, 99, 87–94. [Google Scholar] [CrossRef]
- Simmons, G.S.; Suckling, D.M.; Carpenter, J.E.; Addison, M.F.; Dyck, V.A.; Vreysen, M.J.B. Improved quality management to enhance the efficacy of the sterile insect technique for lepidopteran pests. J. Appl. Èntomol. 2010, 134, 261–273. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Carpenter, J.E.; Marec, F. Improvement of the sterile insect technique for codling moth, Cydia pomonella (Linnaeus) (Lepidoptera Tortricidae) to facilitate expansion of field application. J. Appl. Èntomol. 2010, 134, 165–181. [Google Scholar] [CrossRef]
- Stringer, L.D.; Sullivan, N.J.; Sullivan, T.E.; Mitchell, V.J.; Manning, L.-A.M.; Mas, F.; Hood-Nowotny, R.C.; Suckling, D.M. Attractiveness and competitiveness of irradiated light brown apple moths. Èntomol. Exp. Et Appl. 2013, 148, 203–212. [Google Scholar] [CrossRef]
- Suckling, D.M.; Stringer, L.D.; Mitchell, V.J.; Sullivan, T.E.S.; Sullivan, N.J.; Simmons, G.S.; Barrington, A.M.; El-Sayed, A.M. Comparative fitness of irradiated light brown apple moths (Lepidoptera: Tortricidae) in a wind tunnel, hedgerow, and vineyard. J. Econ. Èntomol. 2011, 104, 1301–1308. [Google Scholar] [CrossRef] [Green Version]
- Henneberry, T.J.; Keaveny, D.F., III. Suppression of Pink Bollworm by Sterile Moth Releases; ARS-32; United States Department of Agriculture, Agricultural Research Service: Springfield, VA, USA, 1985; pp. 1–74.
- Simmons, G.S.; McKemey, A.R.; Morrison, N.I.; O’Connell, S.; Tabashnik, B.E.; Claus, J.; Fu, G.; Tang, G.; Sledge, M.; Walker, A.S.; et al. Field Performance of a Genetically Engineered Strain of Pink Bollworm. PLoS ONE 2011, 6, e024110. [Google Scholar] [CrossRef]
- Bloem, S.; Carpenter, J.E.; Bloem, K.A.; Tomlin, L.; Taggart, S. Effect of Rearing Strategy and Gamma Radiation on Field Competitiveness of Mass-Reared Codling Moths (Lepidoptera: Tortricidae). J. Econ. Entomol. 2004, 97, 1891–1898. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Harari, A.; Seth, R.K.; Wee, S.L.; Conlong, D.E.; Suckling, D.M.; Woods, B.; Lebdi-Grissa, K.; Simmons, G.; Carpenter, J.E. Stable Isotope Markers Differentiate between Mass-Reared and Wild Lepidoptera in Sterile Insect Technique Programs. Fla. Èntomol. 2016, 99, 166–176. [Google Scholar] [CrossRef]
- Suckling, D.M.; Conlong, D.E.; Carpenter, J.E.; Bloem, K.A.; Rendon, P.; Vreysen, M.J.B. Global range expansion of pest Lepidoptera requires socially acceptable solutions. Biol. Invasions 2017, 19, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.B. Monitoring Sterile and Wild Insects in Area-Wide Integrated Pest Management Programmes, in Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Hendrichs, D.V.A.J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 485–528. [Google Scholar]
- Simmons, G.S.; Varela, L.; Daugherty, M.; Cooper, M.; Lance, D.; Mastro, V.; Carde, R.T.; Lucchi, A.; Ioriatti, C.; Bagnoli, B.; et al. Area-wide eradication of the invasive European grapevine moth, Lobesia botrana in California, USA. In Area-Wide Integrated Pest Management. Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 581–596. [Google Scholar]
- Staten, R.T.; Walters, M.L. Technology Used by Field Managers for Pink Bollworm Eradication with Its Successful Outcome in the United States and Mexico, in Area-Wide Integrated Pest Management. Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 51–92. [Google Scholar]
- SAG. SAG y INIA Desarrollan Sistema de Monitoreo Para el Combate de la Lobesia Botrana. 2015. Available online: http://www.sag.cl/noticias/sag-e-inia-desarrollan-sistema-de-monitoreo-para-el-combate-de-la-lobesia-botrana (accessed on 5 March 2021).
- Henderson, P.A.; Southwood, T.R.E. Ecological Methods; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Epopa, P.S.; Millogo, A.A.; Collins, C.M.; North, A.; Tripet, F.; Benedict, M.Q.; Diabate, A. The use of sequential mark-release-recapture experiments to estimate population size, survival and dispersal of male mosquitoes of the Anopheles gambiae complex in Bana, a west African humid savannah village. Parasites Vectors 2017, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Iyaloo, D.; Facknath, S.; Bheecarry, A. A Mark-Release-Recapture Experiment with Radio-Sterilised Aedes albopictus Males as Part of a Sterile Insect Technique Programme against the Vector Mosquito in Panchvati, Mauritius. Afr. Èntomol. 2020, 28, 187–191. [Google Scholar] [CrossRef]
- Bouyer, J.; Balestrino, F.; Culbert, N.; Yamada, H.; Argilés, R.; FAO/IAEA. Guidelines for Mark-Release-Recapture procedures of Aedes mosquitoes; Food and Agriculture Organization of the United Nations/International Atomic Energy Agency: Vienna, Austria, 2020; p. 22. [Google Scholar]
- Vreysen, M.J.B.; Klassen, W.; Carpenter, J.E. Overview of Technological Advances Toward Greater Efficiency and Efficacy in Sterile Insect-Inherited Sterility Programs against Moth Pests. Fla. Èntomol. 2016, 99, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Klassen, W.; Vreysen, M.J.B. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Hendrichs, J.; Enkerlin, W.R.; Pereira, R. Invasive insect pests: Challenges and the role of the sterile insect technique in their prevention, containment, and eradication. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Dyck, V.A.; Graham, S.H.; Bloem, K.A. Implementation of the Sterile Insect Release Programme to Eradicate the Codling Moth, Cydia Pomonella (L) (Lepidoptera: Olethreutidae); International Atomic Energy Agency (IAEA), IAEA: British Columbia, BC, Canada, 1993. [Google Scholar]
- Suckling, D.M.; Barrington, A.M.; Chhagan, A.; Stephens, A.E.A.; Burnip, G.M.; Charles, J.G.; Wee, S.L. Eradication of the Australian Painted Apple Moth Teia anartoides in New Zealand: Trapping, Inherited Sterility, and Male Competitiveness. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 603–615. [Google Scholar]
- Horner, R.; Paterson, G.; Walker, J.T.; Perry, G.L.; Jaksons, R.; Suckling, D.M. Will Peri-Urban Cydia pomonella (Lepidoptera: Tortricidae) Challenge Local Eradication? Insects 2020, 11, 207. [Google Scholar] [CrossRef] [Green Version]
- Kovaleski, A.; Mumford, J. Pulling out the evil by the root: The codling moth Cydia pomonella eradication programme in Brazil. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 581–590. [Google Scholar]
- Schartel, T.E.; Bayles, B.R.; Cooper, M.L.; Simmons, G.S.; Thomas, S.M.; Varela, L.G.; Daugherty, M.P. Reconstructing the European Grapevine Moth (Lepidoptera: Tortricidae), Invasion in California: Insights From a Successful Eradication. Ann. Èntomol. Soc. Am. 2019, 112, 107–117. [Google Scholar] [CrossRef]
- Tobin, P.C.; Kean, J.M.; Suckling, D.M.; McCullough, D.G.; Herms, D.A.; Stringer, L.D. Determinants of successful arthropod eradication programs. Biol. Invasions 2014, 16, 401–414. [Google Scholar] [CrossRef]
- Zalom, F.; Grieshop, J.; Lelea, M.A.; Sedell, J.K. Community Perceptions of Emergency Responses to Invasive Species in California: Case Studies of the Light Brown Apple Moth and the European Grapevine Moth, Report Submitted to USDA in Fulfillment of Cooperative Agreement #10–8100-1531-CA; University of California: Davis, CA, USA, 2013. [Google Scholar]
- Lance, D.R.; Leonard, D.S.; Mastro, V.C.; Walters, M.L. Mating Disruption as a Suppression Tactic in Programs Targeting Regulated Lepidopteran Pests in US. J. Chem. Ecol. 2016, 42, 590–605. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Quantity | Percent |
|---|---|---|
| Tap water | 28,600 | 65.0 |
| Agar agar | 572 | 1.3 |
| Wheat bran | 2640 | 6.0 |
| Corn oil | 44 | 0.1 |
| Toasted soya flour | 3036 | 6.9 |
| Sugar | 792 | 1.8 |
| Wheat germ | 2255 | 5.1 |
| Nipagin | 88 | 0.2 |
| Potassium sorbate | 88 | 0.2 |
| Choline chloride | 88 | 0.2 |
| Formaldehyde (10% solution in water) | 220 | 0.5 |
| Acetic acid (25% solution in water) | 748 | 1.7 |
| Vitamínico, formula B 1 | 176 | 0.4 |
| Sawdust | 4664 | 10.6 |
| Release No. | No. of Moths Released | No. Recaptured | % Recaptured | Mean (SD) Dispersal Distance (m) | Maximum Dispersal Distance (m) |
|---|---|---|---|---|---|
| 1 | 2500 | 41 | 3.3% | 50.5 (18) | 102 |
| 2 | 5000 | 3 | 0.1% | 36 (15.6) | 58 |
| 3 | 5000 | 5 | 0.2% | 86.4 (31.2) | 133 |
| 4 | 5000 | 6 | 0.2% | 71.2 (44.2) | 155 |
| 5 | 5000 | 25 | 1.0% | 66.5 (30) | 133 |
| Total | 22,500 | 80 | 0.7% | 58.8 (28.2) | 155 |
| Day | Mean % Surviving (SD) |
|---|---|
| 1 | 90.0 (7.9) |
| 2 | 76.8 (18.1) |
| 3 | 61.5 (23.3) |
| 4 | 34.4 (23.8) |
| 5 | 19.9 (1.5) |
| 6 | NA |
| 7 | 18.2 (20.9) |
| 8 | 6.0 (7.8) |
| 9 | 1.0 (2.0) |
| 10 | 0 |
| Statistic | Number | Range | Comments |
|---|---|---|---|
| Release period | 20 August 2019 to 5 May 2020 | N/A | Two per week shipments by overnight air cargo from Arica |
| Total No. of shipments | 57 | N/A | 17 missed releases: 1 missed release/week for 19 September 2019, 3 October 2019, 12 November 2019, and 12 December 2019; 7 complete missed weeks for 8 October to 29 October 2019, 11 January to 18 January 2020, and 7 April 2020 |
| Shipping Temperatures | 1.85–7.8 °C (average low, high) | −0.59–9.86 °C | Data available for 11 shipments |
| No. moths released/week, mean (SE) | 20,334 (2705) | 7107–60,430 | |
| No. moths released/ha, mean (SE) | 813 (108) | 284–2417 | |
| Total No. moths released | 752,353 | N/A | |
| Flyers/shipment (%), mean (SE) | 80.9 (2.8)% | 21.9–93.5% | Field laboratory flight testing at ambient outdoor temperatures |
| Statistic | Number | Range | Comments |
|---|---|---|---|
| Total No. of sterile moths captured | 3460 | N/A | |
| Total No. of sterile moths captured in release plot | 3374 | N/A | 97.5% of total recaptures |
| Total No. of sterile moths captured in no-release plot | 86 | N/A | 2.5% of total recaptures |
| Recapture rate of male moths (%), mean (SE) | 0.9 (0.1)% | 0.1–2.2% | Calculated from previous week estimate of male moth release numbers |
| Overflooding ratio, mean (SE) | 2.1 (1.2) | 0–41.4 | No. sterile/No. wild moths captured on traps |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simmons, G.S.; Salazar Sepulveda, M.C.; Fuentes Barrios, E.A.; Idalsoaga Villegas, M.; Medina Jimenez, R.E.; Garrido Jerez, A.R.; Henderson, R.; Donoso Riffo, H. Development of Sterile Insect Technique for Control of the European Grapevine Moth, Lobesia botrana, in Urban Areas of Chile. Insects 2021, 12, 378. https://doi.org/10.3390/insects12050378
Simmons GS, Salazar Sepulveda MC, Fuentes Barrios EA, Idalsoaga Villegas M, Medina Jimenez RE, Garrido Jerez AR, Henderson R, Donoso Riffo H. Development of Sterile Insect Technique for Control of the European Grapevine Moth, Lobesia botrana, in Urban Areas of Chile. Insects. 2021; 12(5):378. https://doi.org/10.3390/insects12050378
Chicago/Turabian StyleSimmons, Gregory. S., Melissa Cristal Salazar Sepulveda, Edith Alejandra Fuentes Barrios, Marcela Idalsoaga Villegas, Raul Enrique Medina Jimenez, Alvaro Rodrigo Garrido Jerez, Ruth Henderson, and Hernán Donoso Riffo. 2021. "Development of Sterile Insect Technique for Control of the European Grapevine Moth, Lobesia botrana, in Urban Areas of Chile" Insects 12, no. 5: 378. https://doi.org/10.3390/insects12050378