
Pupation Substrate Type and Volume Affect Pupation, Quality Parameters and Production Costs of a Reproductive Colony of Ceratitis capitata (Diptera: Tephritidae) VIENNA 8 Genetic Sexing Strain

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Pupation Substrates
2.3. Experimental Design
2.3.1. Experiment 1
2.3.2. Experiment 2
2.4. Experimental Protocol
2.4.1. Experiment 1
2.4.2. Experiment 2
2.5. Statistical Analyses
2.5.1. Experiment 1
2.5.2. Experiment 2
2.5.3. Estimation of Quantities of Larvae and Costs of Substrates Required to Produce One Million Pupae
3. Results
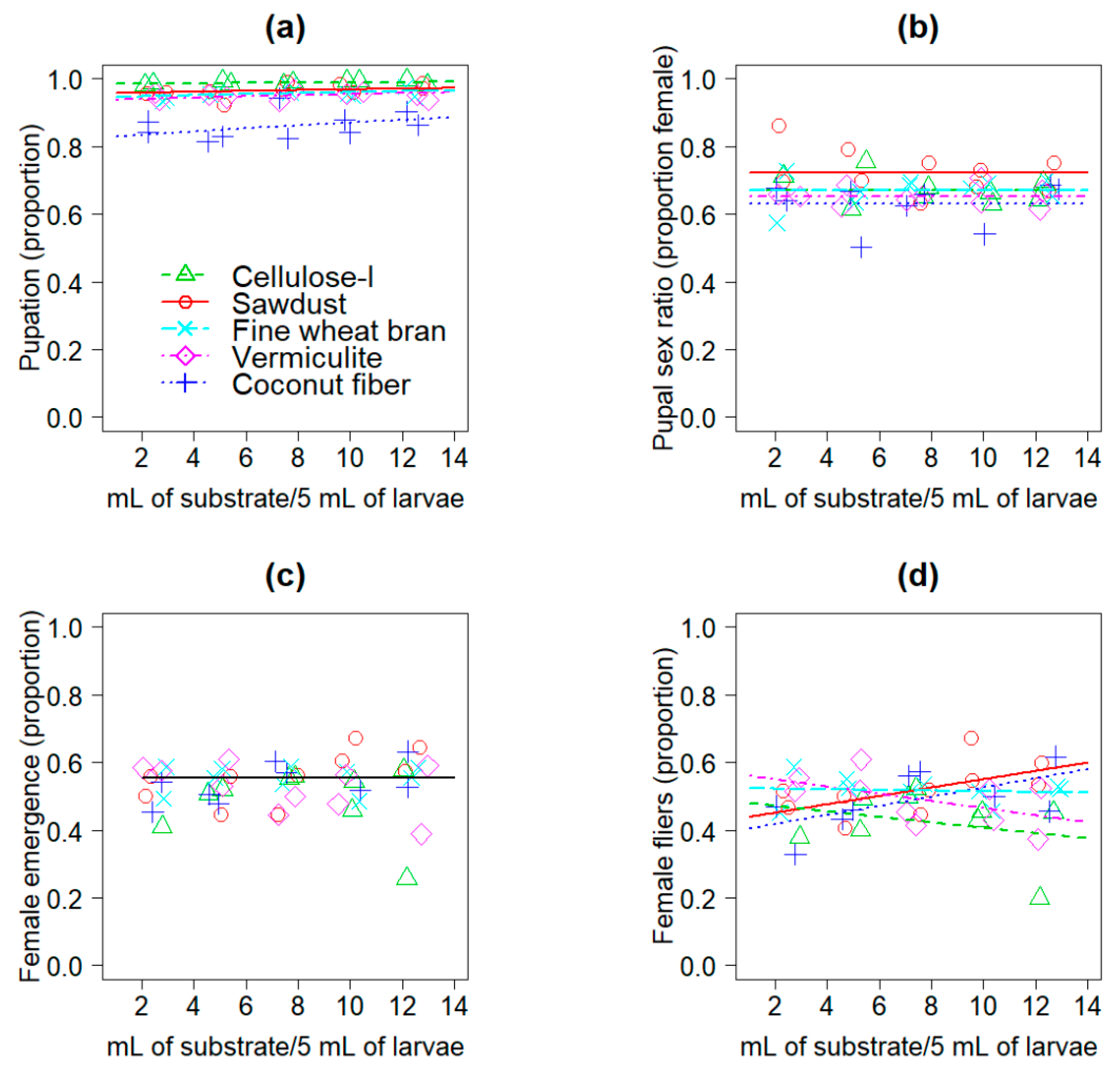
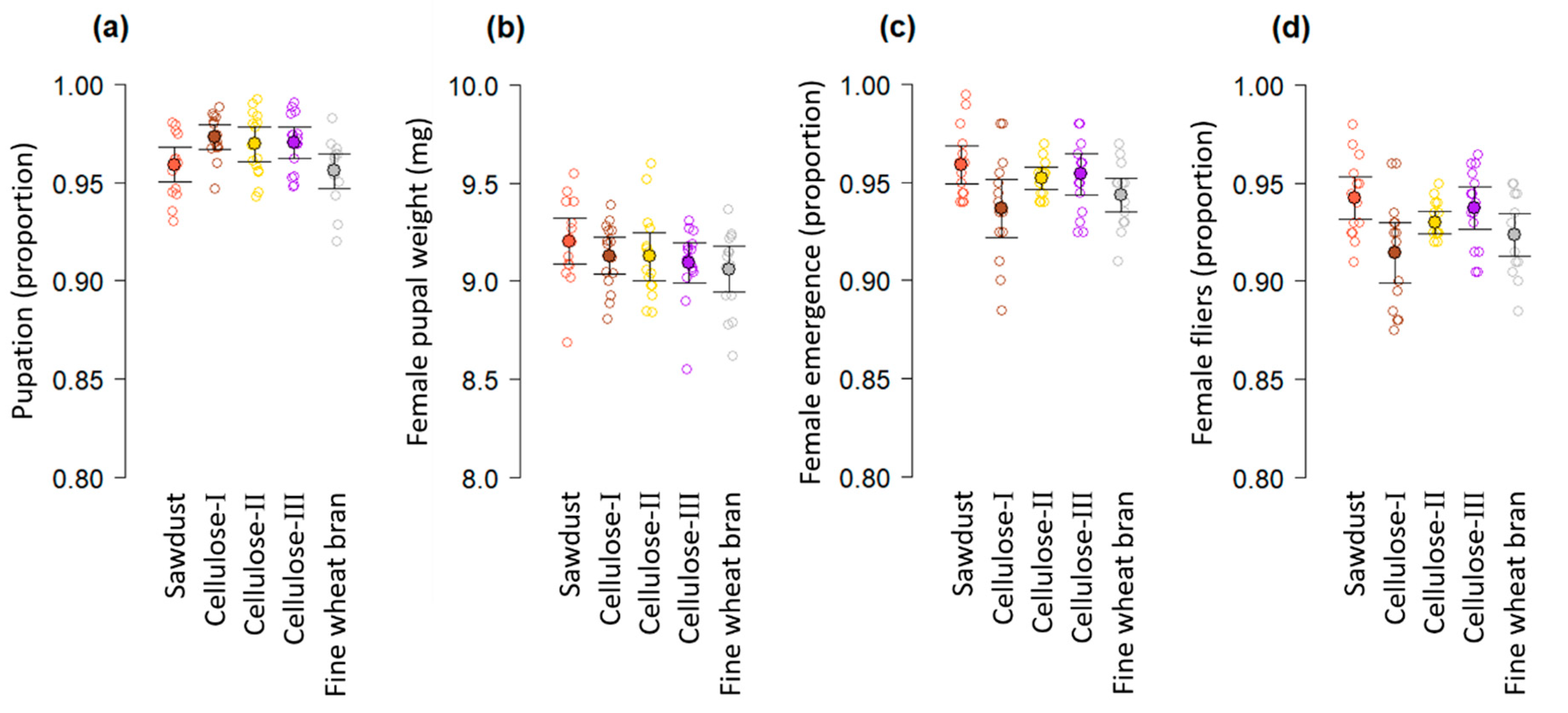
3.1. Experiment 1
3.2. Experiment 2
3.3. Estimation of Quantities of Larvae and Costs of Substrates Required to Produce One Million Pupae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szyniszewska, A.M.; Tatem, A.J. Global assessment of seasonal potential distribution of Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). PLoS ONE 2014, 9, e111582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciarretta, A.; Tabilio, M.R.; Lampazzi, E.; Ceccaroli, C.; Colacci, M.; Trematerra, P. Analysis of the Mediterranean fruit fly [Ceratitis capitata (Wiedemann)] spatio-temporal distribution in relation to sex and female mating status for precision IPM. PLoS ONE 2018, 13, e0195097. [Google Scholar] [CrossRef] [Green Version]
- Liquido, N.J.; Shinoda, L.A.; Cunningham, R.T. Host plants of Mediterranean fruit fly (Diptera: Tephritidae). An annotated world review. Misc. Publ. Entomol. Soc. Am. 1991, 77, 1–52. [Google Scholar]
- Liquido, N.J.; McQuate, G.T.; Suiter, K.A.; Norrbom, A.L.; Yee, W.L.; Chang, C.L. Compendium of Fruit Fly Host Plant Information. In Area-Wide Management of Fruit Fly Pests; Perez-Staples, D., Diaz-Fleischer, F., Montoya, P., Vera, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 363–368. [Google Scholar]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB Int.: Wallingford, Oxon, UK, 1992; 601p. [Google Scholar]
- Hendrichs, J.; Ortiz, G.; Liedo, P.; Schwarz, A. Six years of successful medfly program in Mexico and Guatemala. In Fruit Flies of Economic Importance; Cavalloro, R., Ed.; A. A. Balkema: Rotterdam, The Netherlands, 1983; pp. 353–365. [Google Scholar]
- Enkerlin, W.R.; Gutiérrez Ruelas, J.M.; Pantaleon, R.; Soto Litera, C.; Villaseñor Cortés, A.; Zavala López, J.L.; Orozco Dávila, D.; Montoya Gerardo, P.; Silva Villarreal, L.; Cotoc Roldán, E.; et al. The Moscamed Regional Programme: Review of a success story of area-wide sterile insect technique application. Entomol. Exp. Appl. 2017, 164, 188–203. [Google Scholar] [CrossRef] [Green Version]
- Bakri, A.; Mehta, K.; Lance, D.R. Sterilizing insects with ionizing radiation. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. [Google Scholar]
- Dowell, R.V.; Worley, J.; Gomes, P.J. Sterile insect supply, emergence, and release. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 441–484. [Google Scholar]
- Enkerlin, W.; Gutiérrez-Ruelas, J.M.; Cortes, A.V.; Roldan, E.C.; Midgarden, D.; Lira, E.; López, J.L.Z.; Hendrichs, J.; Liedo, P.; Arriaga, F.J.T. Area freedom in Mexico from Mediterranean fruit fly (Diptera: Tephritidae): A review of over 30 years of a successful containment program using an integrated area-wide SIT approach. Florida Entomol. 2015, 98, 665–681. [Google Scholar] [CrossRef] [Green Version]
- Caceres, C. Mass rearing of temperature sensitive genetic sexing strains in the Mediterranean fruit fly (Ceratitis capitata). Genetica 2002, 116, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K. Genetic sexing strains of Mediterranean fruit fly (Diptera: Tephritidae): Optimizing high temperature treatment of mass-reared temperature-sensitive lethal strains. J. Econ. Entomol. 1998, 91, 1406–1413. [Google Scholar] [CrossRef]
- Fisher, K. Genetic sexing strains of Mediterranean fruit fly (Diptera: Tephritidae): Quality in mass-reared temperature-sensitive lethal strains treated at high temperatures. J. Econ. Entomol. 2000, 93, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Tween, G. MOSCAMED-Guatemala—An evolution of ideas. In Proceedings of the 6th International Symposium on Fruit Flies of Economic Importance, Stellenbosch, South Africa, 6–10 May 2002; Isteg Scientific Publications: Irene, South Africa, 2004; pp. 119–126. [Google Scholar]
- Ramírez y Ramírez, F.; Silva-Villareal, L.; Tejeda, M.; Fuentes, J.; Bello-Rivera, A.; Sánchez-Galicia, F. Funcionamiento, diseño e innovaciones de la nueva planta Moscamed-México. In Book of Abstract, Proceedings of the 9a Reunión del Grupo de Trabajo en Moscas de la Fruta del Hemisferio Occidental, 16–22 October 2016, Buenos Aires, Argentina; Organizing Committee of the 9th Meeting of the Tephritid Workers of the Western Hemisphere: Buenos Aires, Argentina, 2016; pp. 21–26. [Google Scholar]
- Rasgado-Marroquín, M.A.; Velázquez-Dávila, E.; Cruz-De la Cruz, J.A.; Laparra, R.A.; Silva Villareal, L.C.; Tejeda, M.T. A New Diet for a New FacilityDevelopment of a Starter-Finalizer Diet System for Rearing Colonies of the Ceratitis capitata Vienna 8 Strain at a New Facility of Mexico’s Moscamed Program. In Area-Wide Management of Fruit Fly Pests; Perez-staples, D., Dias-Fleischer, F., Montoya, P., Vera, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 217–232. [Google Scholar]
- Yuval, B.; Hendricks, J. Behavior of flies in the genus Ceratitis (Dacinae: Ceratitidini). In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Ratón, FL, USA, 2001; pp. 429–449. [Google Scholar]
- Ozaki, E.T.; Kobayashi, R.M. Effects of pupal handling during laboratory rearing on adult eclosion and flight capability in three tephritid species. J. Econ. Entomol. 1981, 74, 520–525. [Google Scholar] [CrossRef]
- Aceituno-Medina, M.; Rivera-Ciprian, J.P.; Hernández, E. Influence of pupation substrate on mass production and fitness of adult Anastrepha obliqua Macquart (Diptera: Tephritidae) for sterile insect technique application. J. Econ. Entomol. 2017, 110, 2397–2405. [Google Scholar] [CrossRef]
- Langley, P.A.; Maly, H.; Ruhm, F. Application of the sterility principle for the control of the Mediterranean fruit fly (Ceratitis capitata): Pupal metabolism in relation to mass-rearing techniques. Entomol. Exp. Appl. 1972, 15, 23–34. [Google Scholar] [CrossRef]
- Vargas, R.; Chang, H.B.C.; Komura, M.; Kawamoto, D.S. Evaluation of two pupation methods for mass production of Mediterranean fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 1986, 79, 864–867. [Google Scholar] [CrossRef]
- Schwarz, A.J.; Zambada, A.; Orozco, D.H.S.; Zavala, J.L.; Calkins, C.O. Mass production of the Mediterranean fruit fly at Metapa, Mexico. Florida Entomol. 1985, 68, 467–477. [Google Scholar] [CrossRef]
- Celedonio, H.; Enkerlin, H.W.; Bruzzone, D. Preliminary studies for the colonization of Anastrepha obliqua and Anastrepha serpentina (Diptera: Tephritidae). In The South American fruit fly, Anastrepha fraterculus (Wied.); Advances in Artificial Rearing, Taxonomic Status, and Biological Studies; Proceedings of a Workshop organized by the Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture and held in Viña del Mar, Chile, 1–2 November 1996; IAEA-TECDOC—1064; International Atomic Agency: Vienna, Austria, 1999; pp. 33–41. [Google Scholar]
- Fay, H.A.C.; Wornoayporn, V. Inert reusable substrates as potential replacements for wheat bran in larval diets for Mediterranean fruit fly, Ceratitis capitata (Wied.) (Diptera: Tephritidae). J. Appl. Entomol. 2002, 126, 92–96. [Google Scholar] [CrossRef]
- Chang, C.L.; Caceres, C.; Jang, E.B. A novel liquid larval diet and its rearing system for Melon Fly, Bactrocera cucurbitae (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2004, 97, 524–528. [Google Scholar] [CrossRef]
- Domínguez, J.; Trinidad, A.; Solís, E.; Hernández, E. Métodos de colonización y cría masiva. In Moscas de la Fruta: Fundamentos y Procedimientos para su Manejo; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: México City, Mexico, 2010; pp. 259–276. [Google Scholar]
- Sookar, P.; Alleck, M.; Ahseek, N.; Permalloo, S.; Bhagwant, S.; Chang, C.L. Artificial rearing of the peach fruit fly Bactrocera zonata (Diptera: Tephritidae). Int. J. Trop. Insect Sci. 2014, 34, S99–S107. [Google Scholar] [CrossRef]
- Rempoulakis, P.; Taret, G.; Haq, I.; Wornayporn, V.; Ahmad, S.; Sto Tomas, U.; Dammalage, T.; Gembinsky, K.; Franz, G.; Cáceres, C.; et al. Evaluation of quality production parameters and mating behavior of novel genetic sexing strains of the Mediterranean fruit fly Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). PLoS ONE 2016, 11, e0157679. [Google Scholar] [CrossRef] [PubMed]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies; Version 7; International Atomic Energy Agency: Vienna, Austria, 2019; pp. 1–148. Available online: http://www-naweb.iaea.org/nafa/ipc/public/ipc-mass-reared-tephritid.html (accessed on 14 February 2021).
- Vargas, R.I.; Chang, H.B.; Komura, M.; Kawamoto, D. Mortality, stadial duration, and weight loss in three species of mass-reared fruit fly pupae (Diptera: Tephritidae) held with and without vermiculite at selected relative humidities. J. Econ. Entomol. 1987, 80, 972–974. [Google Scholar] [CrossRef]
- NOM [Norma Oficial Mexicana]. NOM-F-317-S-1978. Determinación de pH en Alimentos. Secretaría de Patrimonio y Fomento Industrial.- Direcc. Gral. de Normas.- Depto. de Normalización Nacional.- Oficina de Alimentos, Bebidas y Productos Farmacéuticos. Diario Oficial de la Federación, 3 May 1978. [Google Scholar]
- NMX [Norma Mexicana]. NOM-F-083-1986. Determinación de humedad en productos alimenticios. Dirección General de Normas. Diario Oficial de la Federación, 14 July 1986. [Google Scholar]
- NOM [Norma Oficial Mexicana]. NOM-F-083-1986. Secretaria del Trabajo y Previsión Social. Agentes extinguidores-Polvo químico seco tipo ABC a base de fosfato mono amónico. Secretaría del Trabajo y Previsión Social. Diario Oficial de la Federación, 17 April 2002. [Google Scholar]
- SENASICA. Manual de Pruebas de Control de Calidad de Moscas del Mediterráneo Estériles del CEMM; Programa Moscamed: México City, Mexico; Available online: http://publico.senasica.gob.mx/?doc=24292. (accessed on 23 February 2021).
- Buckley, Y.M. Generalized linear models. In Ecological Statistics: Contemporary Theory and Application; Fox, G.A., Negrete-Yankelevich, S., Sosa, V.J., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 131–147. [Google Scholar]
- Crawley, M.J. The R Book, 2nd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2012; pp. 1–1076. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Warton, D.I.; Hui, F.K. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestel, D.; Tolmasky, D.; Rabossi, A.; Quesada-Allué, L.A. Lipid, carbohydrates and protein patterns during metamorphosis of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2003, 96, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Nestel, D.; Papadopoulos, N.T.; Pascacio-Villafán, C.; Righini, N.; Altuzar-Molina, A.R.; Aluja, M. Resource allocation and compensation during development in holometabolous insects. J. Insect Physiol. 2016, 95, 78–88. [Google Scholar] [CrossRef]
- Merkey, A.B.; Wong, C.K.; Hoshizaki, D.K.; Gibbs, A.G. Energetics of metamorphosis in Drosophila melanogaster. J. Insect Physiol. 2011, 57, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Hooper, G.H.S. Effect of pupation environment on the quality of pupae and adults of the Mediterranean fruit fly. Entomol. Exp. Appl. 1987, 44, 155–159. [Google Scholar] [CrossRef]
- Cavalloro, R.; Delrio, G. Soil factors influencing the pupation of Ceratitis capitata Wiedemann. Boll. Lab. Entomol. Agric. Filippo Silvestri 1975, 32, 190–195. [Google Scholar]
- Eskafi, F.M.; Fernandez, A. Larval-pupal mortality of Mediterranean fruit fly (Diptera: Tephritidae) from interaction of soil, moisture, and temperature. Environ. Entomol. 1990, 19, 1666–1670. [Google Scholar] [CrossRef]
- Jackson, C.G.; Long, J.P.; Klungness, L.M. Depth of pupation in four species of fruit flies (Diptera: Tephritidae) in sand with and without moisture. J. Econ. Entomol. 1998, 91, 138–142. [Google Scholar] [CrossRef]
- Hulthen, A.D.; Clarke, A.R. The influence of soil type and moisture on pupal survival of Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). Aust. J. Entomol. 2006, 45, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Quesada-Moraga, E.; Valverde-García, P.; Garrido-Jurado, I. The effect of temperature and soil moisture on the development of the preimaginal mediterranean fruit fly (Diptera: Tephritidae). Environ. Entomol. 2012, 41, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.C. Ecology of insect rearing systems: A mini-review of insect rearing papers from 1906–2017. Adv. Entomol. 2018, 6, 86–115. [Google Scholar] [CrossRef] [Green Version]
- Pascacio-Villafán, C.; Birke, A.; Williams, T.; Aluja, M. Modeling the cost-effectiveness of insect rearing on artificial diets: A test with a tephritid fly used in the sterile insect technique. PLoS ONE 2017, 12, e0173205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascacio-Villafán, C.; Guillén, L.; Aluja, M. Agar and carrageenan as cost-effective gelling agents in yeast-reduced artificial diets for mass-rearing fruit flies and their parasitoids. Insects 2020, 11, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Pupation Substrate | Cost per kg (US $) 1 | Density (g/cm3) 2 | Moisture (%) 2 | pH 2 | Moisture Retention Capacity (%) 2 | Granularity (Size Range) | Origin |
|---|---|---|---|---|---|---|---|
| Coconut fiber 3 | 0.36 | 0.112 (0.007) | 13.0 (0.05) | 4.5 (0.01) | 804.6 (4.3) | 149–1410 µm | Coconut shell |
| Fine wheat bran 4 | 0.49 | 0.413 (0.003) | 11.2 (0.6) | 6.3 (0.03) | 456.5 (11.5) | 149–1000 µm | Wheat husk |
| Vermiculite 5 | 0.94 | 0.254 (0.001) | 6.8 (0.4) | 5.8 (0.09) | 378.7 (12.6) | 149–1410 µm | Mineral |
| Sawdust 6 | 0.99 | 0.191 (0.018) | 10.1 (0.2) | 4.3 (0.05) | 422.1 (3.7) | 149–1410 µm | Wood |
| Cellulose III 7 | 2.00 | 0.206 (0.014) | 8.8 (0.1) | 4.6 (0.14) | 560.2 (2.2) | 74–149 µm | Wood dust |
| Cellulose II 8 | 2.10 | 0.266 (0.029) | 8.2 (0.1) | 4.4 (0.12) | 500.4 (5.9) | 74–149 µm | Wood dust |
| Cellulose I 9 | 2.10 | 0.080 (0.0004) | 5.0 (0.2) | 7.6 (0.22) | 832.9 (4.2) | 32–200 µm | Recycled paper |
| Pupation Substrate | Substrate Volume (mL of Substrate/5 mL of Larvae) 1 | Predicted Pupation (Proportion) 2 | No. of Larvae Required to Produce 1 Million Pupae 3 | Substrate Volume Required to Produce 1 Million Pupae (mL) 4 | Weight of Substrate Volume Needed to Produce 1 Million Pupae (kg) 5 | Substrate Cost for 1 Million Pupae (US $) 6 |
|---|---|---|---|---|---|---|
| Cellulose I | 2.5 | 0.9860 | 1,014,238 | 9486 | 0.76 | 1.59 |
| Coconut fiber | 12.5 | 0.8804 | 1,135,809 | 53,115 | 5.95 | 2.14 |
| Sawdust | 5.0 | 0.9623 | 1,039,204 | 19,439 | 3.71 | 3.67 |
| Cellulose III | 2.5 | 0.9860 | 1,014,238 | 9486 | 1.95 | 3.91 |
| Fine wheat bran | 5.0 | 0.9526 | 1,049,723 | 19,636 | 8.11 | 3.97 |
| Vermiculite | 5.0 | 0.9460 | 1,057,033 | 19,772 | 5.02 | 4.72 |
| Cellulose II | 2.5 | 0.9860 | 1,014,238 | 9486 | 2.52 | 5.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascacio-Villafán, C.; Quintero-Fong, L.; Guillén, L.; Rivera-Ciprian, J.P.; Aguilar, R.; Aluja, M. Pupation Substrate Type and Volume Affect Pupation, Quality Parameters and Production Costs of a Reproductive Colony of Ceratitis capitata (Diptera: Tephritidae) VIENNA 8 Genetic Sexing Strain. Insects 2021, 12, 337. https://doi.org/10.3390/insects12040337
Pascacio-Villafán C, Quintero-Fong L, Guillén L, Rivera-Ciprian JP, Aguilar R, Aluja M. Pupation Substrate Type and Volume Affect Pupation, Quality Parameters and Production Costs of a Reproductive Colony of Ceratitis capitata (Diptera: Tephritidae) VIENNA 8 Genetic Sexing Strain. Insects. 2021; 12(4):337. https://doi.org/10.3390/insects12040337
Chicago/Turabian StylePascacio-Villafán, Carlos, Luis Quintero-Fong, Larissa Guillén, José Pedro Rivera-Ciprian, Reynaldo Aguilar, and Martín Aluja. 2021. "Pupation Substrate Type and Volume Affect Pupation, Quality Parameters and Production Costs of a Reproductive Colony of Ceratitis capitata (Diptera: Tephritidae) VIENNA 8 Genetic Sexing Strain" Insects 12, no. 4: 337. https://doi.org/10.3390/insects12040337