
Skipping the Insect Vector: Plant Stolon Transmission of the Phytopathogen ‘Ca. Phlomobacter fragariae’ from the Arsenophonus Clade of Insect Endosymbionts



{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction
2.3. Quantification of Phlomobacter Titer in Plant Tissues
2.4. Transmission Electron Microscopy (TEM)
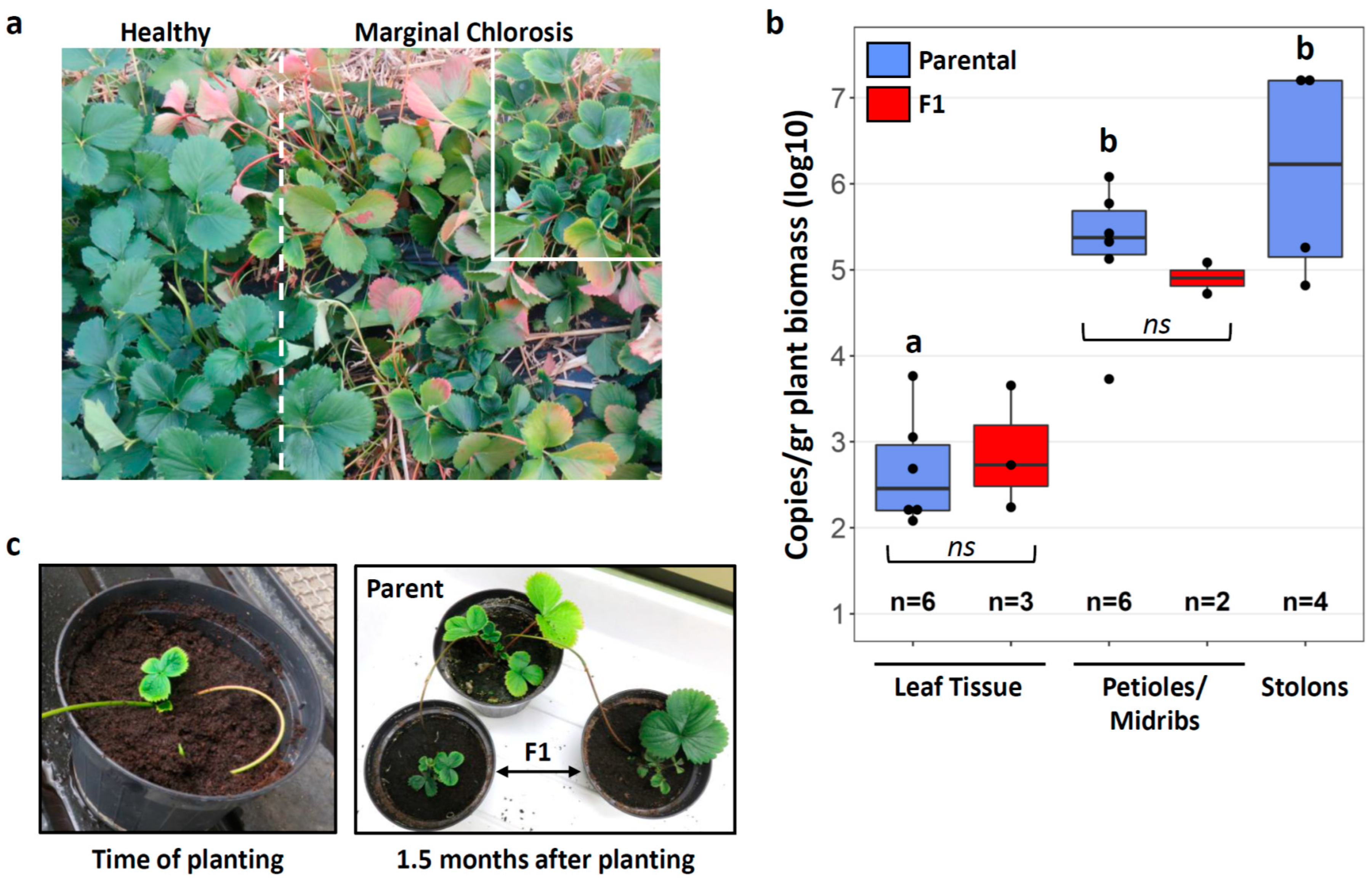
3. Results
3.1. Stolon Transmission of Phlomobacter
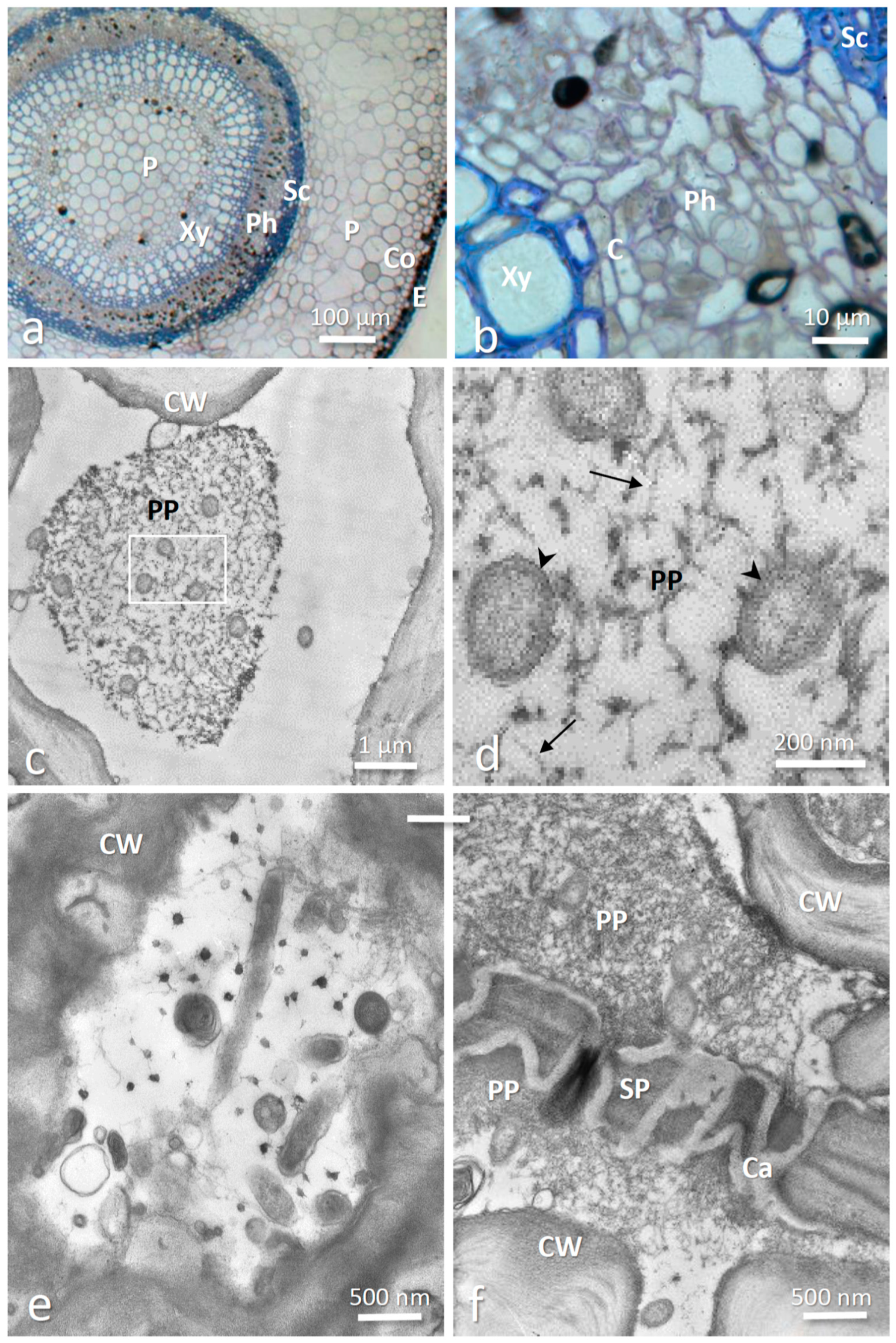
3.2. Intracellular Localization of Phlomobacter in Stolons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.K.; Moran, N.A. Aphid genome expression reveals host-symbiont cooperation in the production of amino acids. Proc. Natl. Acad. Sci. USA 2011, 108, 2849–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, R.; Bennett, G.M.; Cryan, J.R.; Moran, N.A. Evolutionary replacement of obligate symbionts in an ancient and diverse insect lineage. Environ. Microbiol. 2013, 15, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Santos-Garcia, D.; Juravel, K.; Freilich, S.; Zchori-Fein, E.; Latorre, A.; Moya, A.; Morin, S.; Silva, F.J. To B or Not to B: Comparative Genomics Suggests Arsenophonus as a Source of B Vitamins in Whiteflies. Front. Microbiol. 2018, 9, 2254. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [Green Version]
- Jaenike, J.; Unckless, R.; Cockburn, S.N.; Boelio, L.M.; Perlman, S.J. Adaptation via symbiosis: Recent spread of a Drosophila defensive symbiont. Science 2010, 329, 212–215. [Google Scholar] [CrossRef]
- Chrostek, E.; Marialva, M.S.; Esteves, S.S.; Weinert, L.A.; Martinez, J.; Jiggins, F.M.; Teixeira, L. Wolbachia variants induce differential protection to viruses in Drosophila melanogaster: A phenotypic and phylogenomic analysis. PLoS Genet. 2013, 9, e1003896. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Hurst, G.D.; Frost, C.L. Reproductive Parasitism: Maternally Inherited Symbionts in a Biparental World. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, J.; Darby, A.C.; Daniell, T.J.; Godfray, H.C.J.; Douglas, A.E. Linking the bacterial community in pea aphids with host-plant use and natural enemy resistance. Ecol. Entomol. 2004, 29, 60–65. [Google Scholar] [CrossRef]
- Leclercq, S.; Theze, J.; Chebbi, M.A.; Giraud, I.; Moumen, B.; Ernenwein, L.; Greve, P.; Gilbert, C.; Cordaux, R. Birth of a W sex chromosome by horizontal transfer of Wolbachia bacterial symbiont genome. Proc. Natl. Acad. Sci. USA 2016, 113, 15036–15041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, J.P.; Boyd, B.M.; Dale, C. The Life of an Insect Endosymbiont from the Cradle to the Grave. Curr. Biol. 2019, 29, R485–R495. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.P.; Moran, N.A. Extreme genome reduction in symbiotic bacteria. Nat. Rev. Microbiol. 2012, 10, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Orlovskis, Z.; Canale, M.C.; Thole, V.; Pecher, P.; Lopes, J.R.; Hogenhout, S.A. Insect-borne plant pathogenic bacteria: Getting a ride goes beyond physical contact. Curr. Opin. Insect Sci. 2015, 9, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Nadarasah, G.; Stavrinides, J. Insects as alternative hosts for phytopathogenic bacteria. FEMS Microbiol. Rev. 2011, 35, 555–575. [Google Scholar] [CrossRef]
- Chatterjee, S.; Almeida, R.P.; Lindow, S. Living in two worlds: The plant and insect lifestyles of Xylella fastidiosa. Annu. Rev. Phytopathol. 2008, 46, 243–271. [Google Scholar] [CrossRef] [Green Version]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstadter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H.; Skinner, S.W.; Huger, A.M. Male-killing bacteria in a parasitic wasp. Science 1986, 231, 990–992. [Google Scholar] [CrossRef]
- Gherna, R.L.; Werren, J.H.; Weisburg, W.; Cote, R.; Woese, C.R.; Mandelco, L.; Brenner, D.J. Arsenophonus nasoniae gen. nov., sp. nov. the causative agent of the son-killer trait in the parasitic wasp Nasonia vitripennis. Int. J. Syst. Bacteriol. 1991, 41, 563–565. [Google Scholar] [CrossRef]
- Nadal-Jimenez, P.; Griffin, J.S.; Davies, L.; Frost, C.L.; Marcello, M.; Hurst, G.D.D. Genetic manipulation allows in vivo tracking of the life cycle of the son-killer symbiont, Arsenophonus nasoniae, and reveals patterns of host invasion, tropism and pathology. Environ. Microbiol. 2019, 21, 3172–3182. [Google Scholar] [CrossRef] [Green Version]
- Ferree, P.M.; Avery, A.; Azpurua, J.; Wilkes, T.; Werren, J.H. A bacterium targets maternally inherited centrosomes to kill males in Nasonia. Curr. Biol. 2008, 18, 1409–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, O.; Schneppat, U.E.; Berthomieu, A.; Goodman, S.M.; Droz, B.; Paupy, C.; Nkoghe, J.O.; Rahola, N.; Tortosa, P. Origin, acquisition and diversification of heritable bacterial endosymbionts in louse flies and bat flies. Mol. Ecol. 2014, 23, 2105–2117. [Google Scholar] [CrossRef] [PubMed]
- Novakova, E.; Hypsa, V.; Nguyen, P.; Husnik, F.; Darby, A.C. Genome sequence of Candidatus Arsenophonus lipopteni, the exclusive symbiont of a blood sucking fly Lipoptena cervi (Diptera: Hippoboscidae). Stand. Genom. Sci. 2016, 11, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakova, E.; Husnik, F.; Sochova, E.; Hypsa, V. Arsenophonus and Sodalis Symbionts in Louse Flies: An Analogy to the Wigglesworthia and Sodalis System in Tsetse Flies. Appl Environ. Microbiol. 2015, 81, 6189–6199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosokawa, T.; Nikoh, N.; Koga, R.; Sato, M.; Tanahashi, M.; Meng, X.Y.; Fukatsu, T. Reductive genome evolution, host-symbiont co-speciation and uterine transmission of endosymbiotic bacteria in bat flies. ISME J. 2012, 6, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Morse, S.F.; Bush, S.E.; Patterson, B.D.; Dick, C.W.; Gruwell, M.E.; Dittmar, K. Evolution, multiple acquisition, and localization of endosymbionts in bat flies (Diptera: Hippoboscoidea: Streblidae and Nycteribiidae). Appl Environ. Microbiol. 2013, 79, 2952–2961. [Google Scholar] [CrossRef] [Green Version]
- Jousselin, E.; Coeur d’Acier, A.; Vanlerberghe-Masutti, F.; Duron, O. Evolution and diversity of Arsenophonus endosymbionts in aphids. Mol. Ecol. 2013, 22, 260–270. [Google Scholar] [CrossRef]
- Hall, A.A.; Morrow, J.L.; Fromont, C.; Steinbauer, M.J.; Taylor, G.S.; Johnson, S.N.; Cook, J.M.; Riegler, M. Codivergence of the primary bacterial endosymbiont of psyllids versus host switches and replacement of their secondary bacterial endosymbionts. Environ. Microbiol. 2016, 18, 2591–2603. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Ghanim, M.; Gueguen, G.; Kontsedalov, S.; Vavre, F.; Fleury, F.; Zchori-Fein, E. Inherited intracellular ecosystem: Symbiotic bacteria share bacteriocytes in whiteflies. FASEB J. 2008, 22, 2591–2599. [Google Scholar] [CrossRef] [Green Version]
- Kobialka, M.; Michalik, A.; Swierczewski, D.; Szklarzewicz, T. Complex symbiotic systems of two treehopper species: Centrotus cornutus (Linnaeus, 1758) and Gargara genistae (Fabricius, 1775) (Hemiptera: Cicadomorpha: Membracoidea: Membracidae). Protoplasma 2020, 257, 819–831. [Google Scholar] [CrossRef]
- Kobialka, M.; Michalik, A.; Walczak, M.; Junkiert, L.; Szklarzewicz, T. Sulcia symbiont of the leafhopper Macrosteles laevis (Ribaut, 1927) (Insecta, Hemiptera, Cicadellidae: Deltocephalinae) harbors Arsenophonus bacteria. Protoplasma 2016, 253, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salar, P.; Sémétey, O.; Danet, J.L.; Boudon-Padieu, E.; Foissac, X. “Candidatus Phlomobacter fragariae” and the proteobacterium associated with the low sugar content syndrome of sugar beet are related to bacteria of the arsenophonus clade detected in hemipteran insects. Eur. J. Plant. Pathol. 2010, 126, 123–127. [Google Scholar] [CrossRef]
- Bressan, A. Emergence and evolution of Arsenophonus bacteria as insect-vectored plant pathogens. Infect. Genet. Evol. 2014, 22, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Danet, J.L.; Foissac, X.; Zreik, L.; Salar, P.; Verdin, E.; Nourrisseau, J.G.; Garnier, M. “Candidatus Phlomobacter fragariae” Is the Prevalent Agent of Marginal Chlorosis of Strawberry in French Production Fields and Is Transmitted by the Planthopper Cixius wagneri (China). Phytopathology 2003, 93, 644–649. [Google Scholar] [CrossRef] [Green Version]
- Nourrisseau, J.G.; Lansac, M.; Garnier, M. Marginal chlorosis, a new disease of strawberries associated with a bacteriumlike organism. Plant Dis. 1993, 77, 1055–1059. [Google Scholar] [CrossRef]
- Zreik, L.; Bove, J.M.; Garnier, M. Phylogenetic characterization of the bacterium-like organism associated with marginal chlorosis of strawberry and proposition of a Candidatus taxon for the organism, ‘Candidatus Phlomobacter fragariae’. Int. J. Syst. Bacteriol. 1998, 48, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Nao, M.; Usugi, T. Occurrence of strawberry marginal chlorosis caused by “Candidatus Phlomobacter fragariae” in Japan. J. Gen. Plant Pathol. 2006, 72, 374–377. [Google Scholar] [CrossRef]
- Semetey, O.; Gatineau, F.; Bressan, A.; Boudon-Padieu, E. Characterization of a gamma-3 Proteobacteria Responsible for the Syndrome “Basses Richesses” of Sugar Beet Transmitted by Pentastiridius sp. (Hemiptera, Cixiidae). Phytopathology 2007, 97, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Gatineau, F.; Jacob, N.; Vautrin, S.; Larrue, J.; Lherminier, J.; Richard-Molard, M.; Boudon-Padieu, E. Association with the Syndrome “Basses Richesses” of Sugar Beet of a Phytoplasma and a Bacterium-Like Organism Transmitted by a Pentastiridius sp. Phytopathology 2002, 92, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, F.; Babini, A.R.; Lanzoni, C.; Pisi, A.; Credi, R.; Foissac, X.; Salar, P. First report of a γ 3-Proteobacterium associated with diseased strawberries in Italy. Plant Dis. 2007, 91, 1688. [Google Scholar] [CrossRef]
- Salar, P.; Danet, J.L.; Pommier, J.-J.; Foissac, X. The biology of Cixius wagneri, the planthopper vector of ‘Candidatus Phlomobacter fragariae’ in strawberry production tunnels and its consequence for the epidemiology of strawberry marginal chlorosis. Julius Kühn Arch. 2010, 427, 24–26. [Google Scholar]
- Bressan, A.; Semetey, O.; Arneodo, J.; Lherminier, J.; Boudon-Padieu, E. Vector transmission of a plant-pathogenic bacterium in the Arsenophonus clade sharing ecological traits with facultative insect endosymbionts. Phytopathology 2009, 99, 1289–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakova, E.; Hypsa, V.; Moran, N.A. Arsenophonus, an emerging clade of intracellular symbionts with a broad host distribution. BMC Microbiol. 2009, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressan, A.; Terlizzi, F.; Credi, R. Independent origins of vectored plant pathogenic bacteria from arthropod-associated Arsenophonus endosymbionts. Microb. Ecol. 2012, 63, 628–638. [Google Scholar] [CrossRef]
- Danet, J.L.; Fimbeau, S.; Pommier, J.-J.; Couture, C.; Foissac, X. Detection of phloem restricted bacteria responsible for strawberry marginal chlorosis (SMC) by real-time PCR in a single assay. Julius Kühn Arch. 2010, 427, 35–38. [Google Scholar]
- Batailler, B.; Lemaitre, T.; Vilaine, F.; Sanchez, C.; Renard, D.; Cayla, T.; Beneteau, J.; Dinant, S. Soluble and filamentous proteins in Arabidopsis sieve elements. Plant Cell Environ. 2012, 35, 1258–1273. [Google Scholar] [CrossRef]
- Faoro, F.; Tornaghi, R. Effects of simultaneous fixation with glutaraldehyde, picric acid and osmium tetroxide on P-proteins in Vitis vinifera. Caryologia 1982, 35, 372–373. [Google Scholar]
- Ernst, A.M.; Jekat, S.B.; Zielonka, S.; Muller, B.; Neumann, U.; Ruping, B.; Twyman, R.M.; Krzyzanek, V.; Prufer, D.; Noll, G.A. Sieve element occlusion (SEO) genes encode structural phloem proteins involved in wound sealing of the phloem. Proc. Natl. Acad. Sci. USA 2012, 109, E1980–E1989. [Google Scholar] [CrossRef] [Green Version]
- Knoblauch, M.; Peters, W.S.; Ehlers, K.; van Bel, A.J. Reversible calcium-regulated stopcocks in legume sieve tubes. Plant Cell 2001, 13, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Dinant, S.; Clark, A.M.; Zhu, Y.; Vilaine, F.; Palauqui, J.C.; Kusiak, C.; Thompson, G.A. Diversity of the superfamily of phloem lectins (phloem protein 2) in angiosperms. Plant Physiol. 2003, 131, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Koh, E.J.; Zhou, L.; Williams, D.S.; Park, J.; Ding, N.; Duan, Y.P.; Kang, B.H. Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with “Candidatus Liberibacter asiaticus”. Protoplasma 2012, 249, 687–697. [Google Scholar] [CrossRef]
- Kim, J.S.; Sagaram, U.S.; Burns, J.K.; Li, J.L.; Wang, N. Response of sweet orange (Citrus sinensis) to ‘Candidatus Liberibacter asiaticus’ infection: Microscopy and microarray analyses. Phytopathology 2009, 99, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musetti, R.; Buxa, S.V.; De Marco, F.; Loschi, A.; Polizzotto, R.; Kogel, K.H.; van Bel, A.J. Phytoplasma-triggered Ca(2+) influx is involved in sieve-tube blockage. Mol. Plant Microbe Interact. 2013, 26, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musetti, R.; Paolacci, A.; Ciaffi, M.; Tanzarella, O.A.; Polizzotto, R.; Tubaro, F.; Mizzau, M.; Ermacora, P.; Badiani, M.; Osler, R. Phloem cytochemical modification and gene expression following the recovery of apple plants from apple proliferation disease. Phytopathology 2010, 100, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faoro, F.; Tornaghi, R. Cytochemical identification of callose deposits around plasmodesmata of virus-infected phloem cells. Ultramicroscopy 1983, 12, 162–163. [Google Scholar] [CrossRef]
- Faoro, F.; Tornaghi, R.; Belli, G. Localization of closteroviruses on grapevine thin sections and their identification by immunogold labelling. J. Phytopathol. 1991, 133, 297–306. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Watson, D.C.; Gordon, T.R. Transmission of Fusarium oxysporum f. sp. fragariae through stolons in strawberry plants. Plant Dis. 2019, 103, 1249–1251. [Google Scholar] [CrossRef]
- Gordon, T.R.; Kirkpatrick, S.C.; Shaw, D.V.; Larson, K.D. Differential infection of mother and runner plant generations by Verticillium dahliae in a high elevation strawberry nursery. HortScience 2002, 37, 927–931. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dittmer, J.; Lusseau, T.; Foissac, X.; Faoro, F. Skipping the Insect Vector: Plant Stolon Transmission of the Phytopathogen ‘Ca. Phlomobacter fragariae’ from the Arsenophonus Clade of Insect Endosymbionts. Insects 2021, 12, 93. https://doi.org/10.3390/insects12020093
Dittmer J, Lusseau T, Foissac X, Faoro F. Skipping the Insect Vector: Plant Stolon Transmission of the Phytopathogen ‘Ca. Phlomobacter fragariae’ from the Arsenophonus Clade of Insect Endosymbionts. Insects. 2021; 12(2):93. https://doi.org/10.3390/insects12020093
Chicago/Turabian StyleDittmer, Jessica, Thierry Lusseau, Xavier Foissac, and Franco Faoro. 2021. "Skipping the Insect Vector: Plant Stolon Transmission of the Phytopathogen ‘Ca. Phlomobacter fragariae’ from the Arsenophonus Clade of Insect Endosymbionts" Insects 12, no. 2: 93. https://doi.org/10.3390/insects12020093