
Sexual Competitiveness and Induced Egg Sterility by Aedes aegypti and Aedes albopictus Gamma-Irradiated Males: A Laboratory and Field Study in Mexico

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Strains
2.2. Pupae Irradiation
2.3. Competitiveness Experiments
2.3.1. Laboratory Cages
2.3.2. Field-Cage Test
2.4. Statistical Analysis
3. Results
3.1. Laboratory Study
3.1.1. Insemination of Females
3.1.2. Egg Production
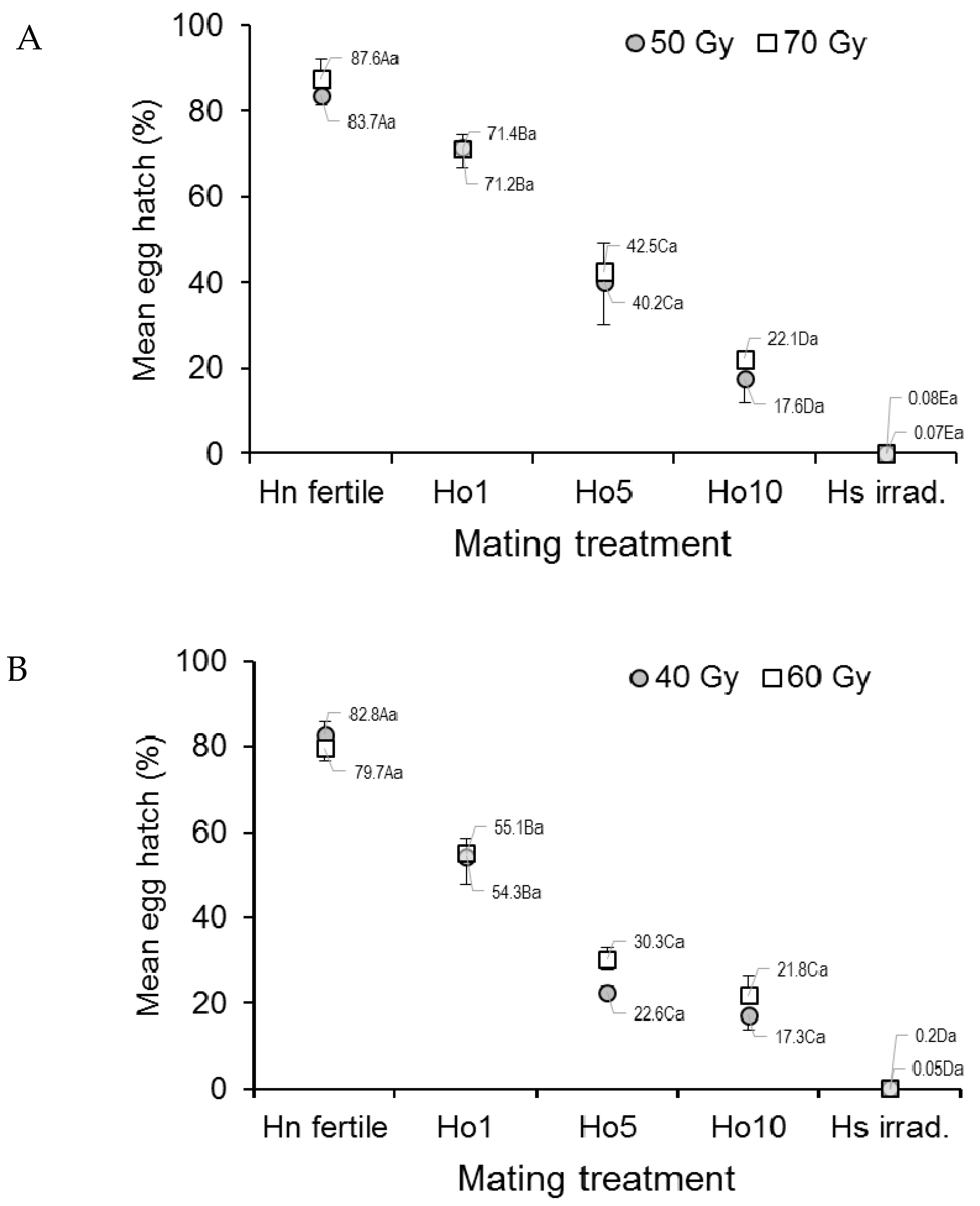
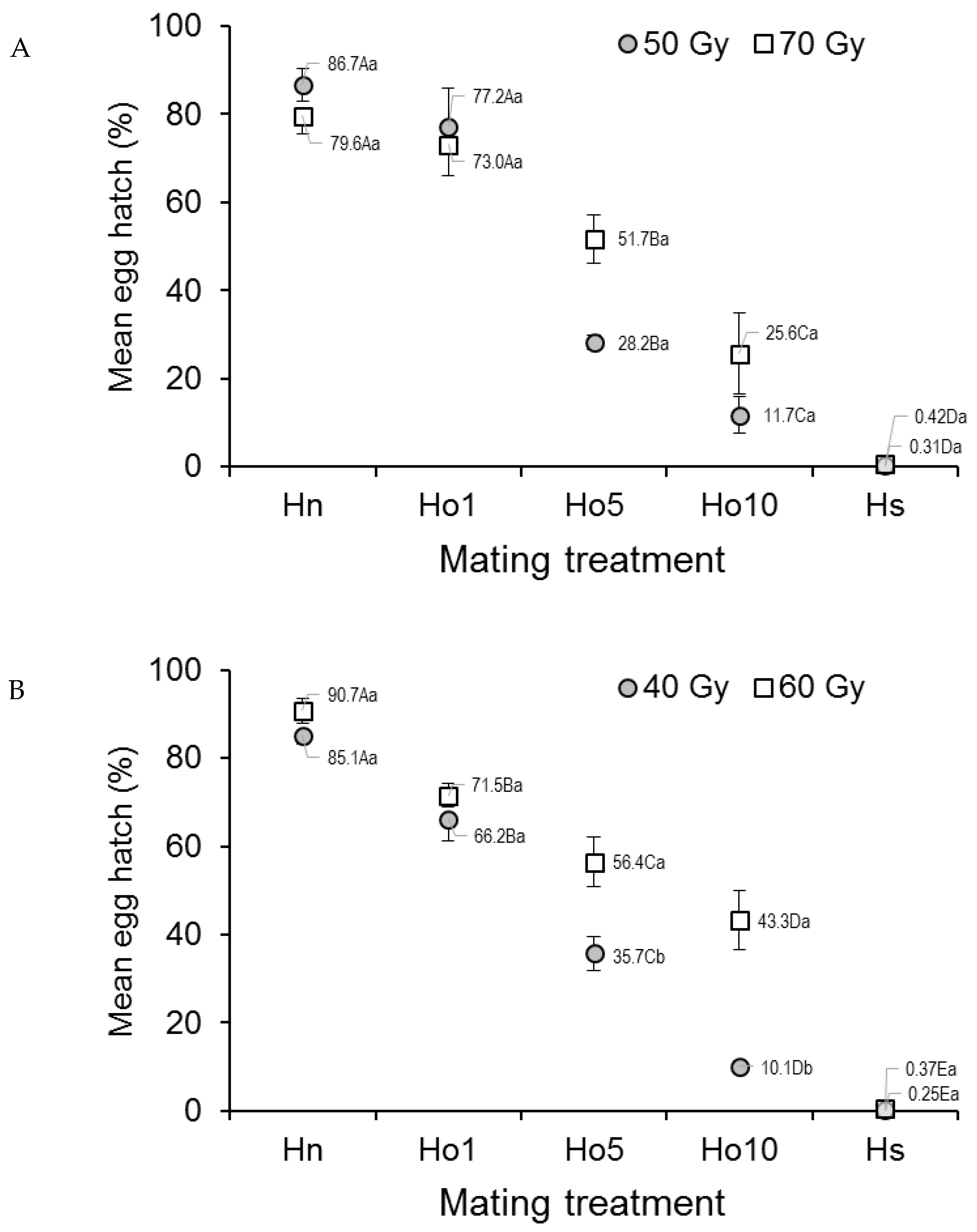
3.1.3. Egg Hatch
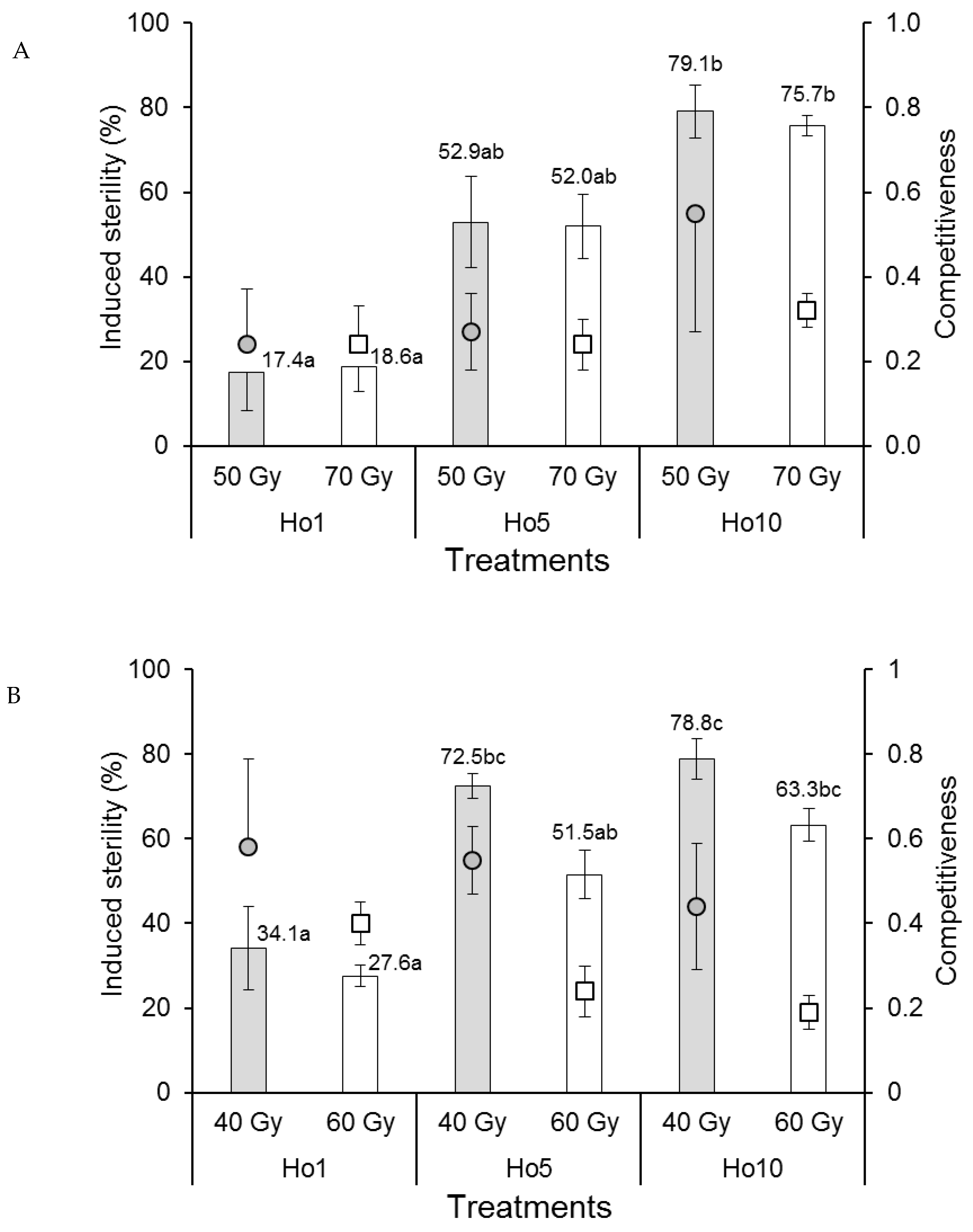
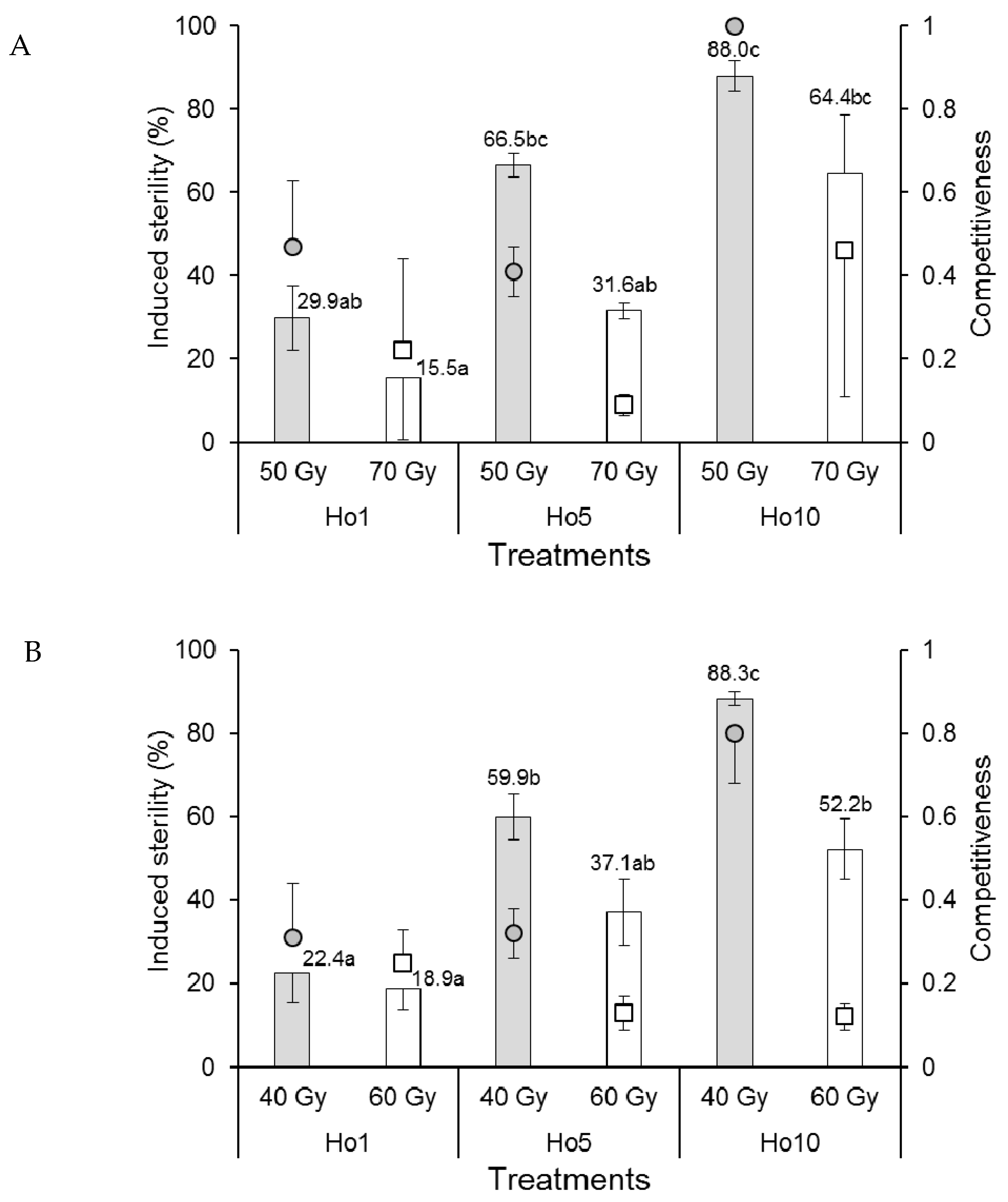
3.1.4. Male Competitiveness and Induced Egg Sterility
3.2. Field-Cage Conditions
3.2.1. Insemination of Females
3.2.2. Egg Production
3.2.3. Egg Hatch
3.2.4. Male Competitiveness and Induced Egg Sterility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Powell, J.R. Mosquito-borne human viral diseases: Why Aedes aegypti? Am. J. Trop. Med. Hyg. 2018, 98, 1563–1565. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Bortel, W.V.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Salas, I.; Danis-Lozano, R.; Casas-Martínez, M.; Ulloa, A.; Bond, J.G.; Marina, C.F.; Lopez-Ordóñez, T.; Elizondo-Quiroga, A.; Torres-Monzón, J.A.; Díaz-González, E.E. Historical inability to control Aedes aegypti as a main contributor of fast dispersal of chikungunya outbreaks in Latin America. Antivir. Res. 2015, 124, 30–42. [Google Scholar] [CrossRef] [PubMed]
- CDC. Surveillance and Control of Aedes aegypti and Aedes albopictus in the United States. 2016. Available online: https://www.cdc.gov/chikungunya/pdfs/surveillance-and-control-of-aedes-aegypti-andaedes-albopictus-us.pdf (accessed on 14 August 2020).
- WHO. Global Strategy for Dengue Prevention and Control 2012–2020; World Health Organization: Geneva, Switzerland, 2012; Volume 27, p. 35. Available online: https://apps.who.int/iris/bitstream/handle/10665/75303/9789241504034_eng.pdf (accessed on 11 August 2020).
- Secretaría de Salud. Dirección General de Epidemiología, Panorama Epidemiológico de Dengue 2019. Available online: https://www.gob.mx/cms/uploads/attachment/file/524262/Pano_dengue_52_2019.pdf (accessed on 4 December 2020).
- Beltrán-Silva, S.L.; Chacón-Hernández, S.S.; Moreno-Palacios, E.; Pereyra-Molina, J.A. Clinical and differential diagnosis: Dengue, chikungunya and Zika. Rev. Médica Hosp. Gen. México 2018, 81, 146–153. [Google Scholar] [CrossRef]
- Kittayapong, P.; Ninphanomchai, S.; Limohpasmanee, W.; Chansang, C.; Chansang, U.; Mongkalangoon, P. Combined sterile insect technique and incompatible insect technique: The first proof-of-concept to suppress Aedes aegypti vector populations in semi-rural settings in Thailand. PLoS Negl. Trop. Dis. 2019, 13, e0007771. [Google Scholar] [CrossRef] [Green Version]
- Pan American Health Organization (PAHO). Evaluation of Innovative Strategies for Aedes Aegypti Control: Challenges for Their Introduction and Impact Assessment; PAHO: Washington, DC, USA, 2019; p. 62. Available online: https://iris.paho.org/handle/10665.2/51375 (accessed on 8 December 2020).
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Gouagna, L.C.; Damiens, D.; Oliva, C.F.; Boyer, S.; Goff, G.L.; Brengues, C.; Dehecq, J.S.; Raude, J.; Simard, F.; Fontenille, D. Strategic approach, advances, and challenges in the development and application of the SIT for area-wide control of Aedes albopictus mosquitoes in Reunion Island. Insects 2020, 11, 770. [Google Scholar] [CrossRef]
- Bellini, R.; Balestrino, F.; Medici, A.; Gentile, G.; Veronesi, R.; Carrieri, M. Mating competitiveness of Aedes albopictus radio-sterilized males in large enclosures exposed to natural conditions. J. Med. Entomol. 2013, 50, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Klassen, W. Area-Wide integrated pest management and the sterile insect technique. In Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–68. [Google Scholar]
- Huang, Y.S.; Higgs, S.; Vanlandingham, D.L. Biological control strategies for mosquito vectors of arboviruses. Insects 2017, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.M.; McKenzie, L.J. Fluctuating asymmetry as a quality control indicator for insect mass rearing processes. J. Econ. Entomol. 1992, 85, 2045–2050. [Google Scholar] [CrossRef]
- Madakacherry, O.; Lees, R.S.; Gilles, J.R.L. Aedes albopictus (Skuse) males in laboratory and semi-field cages: Release ratios and mating competitiveness. Acta Trop. 2014, 1325, S124–S129. [Google Scholar] [CrossRef]
- Bond, J.G.; Osorio, A.R.; Avila, N.; Gómez-Simuta, Y.; Marina, C.F.; Fernández-Salas, I.; Liedo, P.; Dor, A.; Carvalho, D.O.; Bourtzis, K.; et al. Optimization of irradiation dose to Aedes aegypti and Ae. albopictus in a sterile insect technique program. PLoS ONE 2019, 14, e0212520. [Google Scholar] [CrossRef]
- Parker, A.; Mehta, K. Sterile insect technique: A model for dose optimization for improved sterile insect quality. Fla. Entomol. 2007, 90, 88–95. [Google Scholar] [CrossRef]
- McInnis, D.O.; Lance, D.R.; Jackson, G.C. Behavioral resistance to the sterile insect technique by the Mediterranean fruit fly (Diptera: Tephritidae) in Hawaii. Ann. Entomol. Soc. Am. 1996, 89, 739–744. [Google Scholar] [CrossRef]
- Maïga, H.; Damiens, D.; Niang, A.; Sawadogo, S.P.; Fatherhaman, O.; Lees, R.S.; Roux, O.; Dabiré, R.K.; Quédraogo, G.A.; Tripet, F.; et al. Mating competitiveness of sterile male Anopheles coluzzii in large cages. Malar. J. 2014, 13, 460. [Google Scholar] [CrossRef] [Green Version]
- Damiens, D.; Lebon, C.; Wilkinson, D.A.; Dijoux-Millet, D.; Le Goff, G.; Bheecarry, A.; Gouagna, L.C. Cross-mating compatibility and competitiveness among Aedes albopictus strains from distinct geographic origins—Implications for future application of SIT programs in the South West Indian Ocean Islands. PLoS ONE 2016, 11, e0163788. [Google Scholar] [CrossRef]
- Bond, J.G.; Ramírez-Osorio, A.; Marina, C.F.; Fernández-Salas, I.; Liedo, P.; Dor, A.; Williams, T. Efficiency of two larval diets for mass-rearing of the mosquito Aedes aegypti. PLoS ONE 2017, 12, e0187420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IAEA. Dosimetry for Food Irradiation; Technical Reports Series No. 409; International Atomic Energy Agency: Vienna, Austria, 2002; p. 161. Available online: https://www-pub.iaea.org/MTCD/Publications/PDF/TRS409_scr.pdf (accessed on 8 September 2020).
- IAEA. Gafchromic® Dosimetry System for SIT—Standard Operating Procedure. 2004. Available online: http://www-naweb.iaea.org/nafa/ipc/public/ipc-gafchromicdosimetry-sterile-insect-technique.html (accessed on 12 August 2020).
- Fried, M. Determination of sterile-insect competitiveness. J. Econ. Entomol. 1971, 64, 869–872. [Google Scholar] [CrossRef]
- Jamovi. Jamovi Statistical Software v.1.2.27.0. 2020. Available online: https://www.jamovi.org (accessed on 15 September 2020).
- Yamada, H.; Vreysen, M.J.B.; Gilles, J.R.L.; Munhenga, G.; Damiens, D.D. The effects of genetic manipulation, dieldrin treatment and irradiation on the mating competitiveness of male Anopheles arabiensis in field cages. Malar. J. 2014, 13, 318. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.F.; Jacquet, M.; Gilles, J.; Lemperiere, G.; Maquart, P.O.; Quilici, S.; Schooneman, F.; Vreysen, M.J.B.; Boyer, S. The sterile insect technique for controlling populations of Ae. albopictus (Diptera: Culicidae) on Reunion Island: Mating vigour of sterilized males. PLoS ONE 2012, 7, e49414. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Lees, R.S.; Xi, Z.; Gilles, J.R.L.; Bourtzis, K. Combining the sterile insect technique with Wolbachia-based approaches: II—A safe approach to Aedes albopictus population suppression programmes, designed to minimize the consequences of inadvertent female release. PLoS ONE 2015, 10, e0135194. [Google Scholar] [CrossRef]
- Munhenga, G.; Brooke, B.D.; Gilles, J.R.L.; Slabbert, K.; Kemp, A.; Dandalo, L.C.; Wood, O.R.; Lobb, L.N.; Govender, D.; Renke, M.; et al. Mating competitiveness of sterile genetic sexing strain males (GAMA) under laboratory and semi-field conditions: Steps towards the use of the Sterile Insect Technique to control the major malaria vector Anopheles arabiensis in South Africa. Parasites Vectors 2016, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, D.O.; Torres-Monzon, J.A.; Koskinioti, P.; Wijegunawardana, N.D.A.D.; Liang, X.; Pillwax, G.; Xi, Z.; Bourtzis, K. Aedes aegypti lines for combined sterile insect technique and incompatible insect technique applications: The importance of host genomic background. Entomol. Exp. Appl. 2020, 168, 560–572. [Google Scholar] [CrossRef]
- Damiens, D.; Soliban, S.M.; Balestrino, F.; Alsir, R.; Vreysen, M.J.B. Different blood and sugar feeding regimes affect the productivity of Anopheles arabiensis colonies (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 336–343. [Google Scholar] [CrossRef]
- Soma, D.D.; Maïga, H.; Mamai, W.; Bimbile-Somda, N.S.; Venter, N.; Ali, A.B.; Yamada, H.; Diabaté, A.; Fournet, F.; Ouédraogo, G.A.; et al. Does mosquito mass-rearing produce an inferior mosquito? Malar. J. 2017, 16, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestrino, F.; Medici, A.; Candini, G.; Carrieri, M.; Maccagnani, B.; Calvitti, M.; Maini, S.; Bellini, R. γ Ray dosimetry and mating capacity studies in laboratory on Aedes albopictus males. J. Med. Entomol. 2010, 47, 581–591. [Google Scholar] [CrossRef]
- Du, W.; Hu, C.; Yu, C.; Tong, J.; Qui, J.; Zhang, S.; Liu, Y. Comparison between pupal and adult X-ray radiation, designed for the sterile insect technique for Aedes albopictus control. Acta Trop. 2019, 199, 105110. [Google Scholar] [CrossRef]
- Bouyer, J.; Vreysen, M.J.B. Yes, irradiated sterile male mosquitoes can be sexually competitive! Trends Parasitol. 2020, 36, 877–880. [Google Scholar] [CrossRef]
- Benedict, M.Q.; Robinson, A.S. The first releases of transgenic mosquitoes: An argument for the sterile insect technique. Trends Parasitol. 2003, 19, 349–355. [Google Scholar] [CrossRef]
- Helinski, M.E.H.; Parker, A.G.; Knols, B.G.J. Radiation-induced sterility for pupal and adult stages of the malaria mosquito Anopheles arabiensis. Malar. J. 2006, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Ernawan, B.; Tambunan, U.S.F.; Sugoro, I.; Sasmita, H. Effects of gamma irradiation dose-rate on sterile male Aedes aegypti. In Proceedings of the International Biology Conference 2016, Azores, Portugal, 18–22 July 2016; Available online: https://aip.scitation.org/doi/pdf/10.1063/1.4985401 (accessed on 12 August 2020).
- Andrearsen, M.H.; Curtis, C.F. Optimal life stages for radiation sterilization of Anopheles for sterile insect release. Med. Vet. Entomol. 2005, 19, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lees, R.S.; Xi, Z.; Bourtzis, K.; Gilles, J.R.L. Combining the sterile insect technique with the incompatible insect technique: III- Robust mating competitiveness of irradiated triple Wolbachia-Infected Aedes albopictus male under semi-field conditions. PLoS ONE 2016, 11, e0151864. [Google Scholar] [CrossRef] [PubMed]
- Helinski, M.E.H.; Parker, A.G.; Knols, B.G.J. Radiation biology of mosquitoes. Malar. J. 2009, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Calkins, C.O.; Parker, A.G. Sterile insect quality. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 269–296. [Google Scholar]
- Bellini, R.; Calvitti, M.; Medici, A.; Carrieri, M.; Celli, G.; Maini, S. Use of the sterile insect technique against Aedes albopictus in Italy: First results of a pilot trial. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 505–515. [Google Scholar]
- Harrington, L.C.; Ponlawat, A.; Edman, J.D.; Scott, T.W.; Vermeylen, F. Influence of container size, location, and time of day on oviposition patterns of the dengue vector, Aedes aegypti, in Thailand. Vector-Borne Zoonotic Dis. 2008, 8, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meza-Hernandez, J.S.; Díaz-Fleischer, F. Comparison of sexual compatibility between laboratory and wild Mexican fruit flies under laboratory and field conditions. J. Econ. Entomol. 2006, 99, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Facchinelli, L.; Valerio, L.; Ramsey, J.M.; Gould, F.; Walsh, R.K.; Bond, G.; Robert, M.A.; Lloid, A.L.; James, A.A.; Alphey, L.; et al. Field cage studies and progressive evaluation of genetically-engineered mosquitoes. PLoS Negl. Trop. Dis. 2013, 7, e2001. [Google Scholar] [CrossRef]
- Helinski, M.E.H.; Knols, B.G.J. Mating competitiveness of male Anopheles arabiensis mosquitoes irradiated with a partially or fully sterilizing dose in small and large laboratory cages. J. Med. Entomol. 2008, 45, 698–705. [Google Scholar] [CrossRef]
- Marina, C.F.; Bond, J.G.; Hernández-Arriaga, K.; Valle, J.; Ulloa, A.; Fernández-Salas, I.; Carvalho, D.O.; Bourtzis, K.; Dor, A.; Williams, T.; et al. Population dynamics of Aedes aegypti and Aedes albopictus in two rural villages in southern Mexico: Baseline data for an evaluation of the sterile insect technique. Insects 2021, 12, 58. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species, Mating Treatment | Dose (Gy) | Mean Female Insemination ± SE (%) 1 | Mean Egg Production/female ± SE 1 |
|---|---|---|---|
| A: Ae. aegypti | |||
| Hn (fertile) | − | 83 ± 3 a | 34.4 ± 3.8 ab |
| Ho1 | 50 | 77 ± 13 ab | 33.5 ± 2.9 ab |
| Ho5 | 50 | 80 ± 6 ab | 33.4 ± 1.9 ab |
| Ho10 | 50 | 90 ± 6 a | 40.4 ± 2.3 a |
| Hs (irrad.) | 50 | 63 ± 7 b | 24.0 ± 3.6 b |
| Hn (fertile) | - | 83 ± 3 a | 34.8 ± 0.9 ab |
| Ho1 | 70 | 67 ± 9 ab | 34.0 ± 0.9 ab |
| Ho5 | 70 | 63 ± 12 ab | 31.0 ± 1.6 ab |
| Ho10 | 70 | 70 ± 12 ab | 38.2 ± 3.0 a |
| Hs (irrad.) | 70 | 50 ± 6 b | 31.3 ± 0.8 b |
| B: Ae. albopictus | |||
| Hn (fertile) | − | 77 ± 3 a | 44.4 ± 2.7 a |
| Ho1 | 40 | 57 ± 3 b | 34.3 ± 8.1 ab |
| Ho5 | 40 | 63 ± 9 ab | 33.9 ± 9.7 ab |
| Ho10 | 40 | 73 ± 3 a | 41.3 ± 5.9 a |
| Hs (irrad.) | 40 | 50 ± 6 b | 23.9 ± 1.6 b |
| Hn (fertile) | − | 73 ± 3 a | 34.2 ± 3.9 ab |
| Ho1 | 60 | 53 ± 3 b | 26.1 ± 3.5 ab |
| Ho5 | 60 | 60 ± 6 ab | 28.5 ± 6.4 ab |
| Ho10 | 60 | 70 ± 6 a | 42.8 ± 12.1 a |
| Hs (irrad.) | 60 | 43 ± 3 b | 20.0 ± 0.3 b |
| Species, Mating Treatment | Dose (Gy) | Mean Female Insemination ± SE (%) 1 | Mean Egg Production/Female ± SE 1 |
|---|---|---|---|
| A: Ae. aegypti | |||
| Hn (fertile) | − | 83 ± 3 ab | 21.9 ± 1.1 a |
| Ho1 | 50 | 70 ± 6 ab | 13.5 ± 3.0 ab |
| Ho5 | 50 | 73 ± 7 ab | 18.0 ± 1.5 ab |
| Ho10 | 50 | 87 ± 3 a | 22.1 ± 3.4 a |
| Hs (irrad.) | 50 | 60 ± 6 b | 15.2 ± 1.7 b |
| Hn (fertile) | − | 73 ± 12 ab | 19.2 ± 0.6 a |
| Ho1 | 70 | 63 ± 3 ab | 17.5 ± 0.8 ab |
| Ho5 | 70 | 67 ± 9 ab | 16.2 ± 0.4 ab |
| Ho10 | 70 | 77 ± 9 a | 16.5 ± 1.0 a |
| Hs (irrad.) | 70 | 53 ± 12 b | 11.6 ± 1.8 b |
| B: Ae. albopictus | |||
| Hn (fertile) | − | 60 ± 6 a | 30.4 ± 2.5 a |
| Ho1 | 40 | 47 ± 3 ab | 22.4 ± 1.3 b |
| Ho5 | 40 | 53 ± 3 ab | 22.0 ± 0.9 ab |
| Ho10 | 40 | 67 ± 92 a | 24.8 ± 3.9 ab |
| Hs (irrad.) | 40 | 37 ± 3 b | 15.2 ± 4.0 c |
| Hn (fertile) | − | 70 ± 12 a | 24.1 ± 0.5 a |
| Ho1 | 60 | 43 ± 9 ab | 17.6 ± 0.8 b |
| Ho5 | 60 | 47 ± 12 ab | 19.2 ± 0.7 ab |
| Ho10 | 60 | 77 ± 9 a | 23.9 ± 0.7 ab |
| Hs (irrad.) | 60 | 40 ± 6 b | 8.5 ± 2.7 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bond, J.G.; Aguirre-Ibáñez, S.; Osorio, A.R.; Marina, C.F.; Gómez-Simuta, Y.; Tamayo-Escobar, R.; Dor, A.; Liedo, P.; Carvalho, D.O.; Williams, T. Sexual Competitiveness and Induced Egg Sterility by Aedes aegypti and Aedes albopictus Gamma-Irradiated Males: A Laboratory and Field Study in Mexico. Insects 2021, 12, 145. https://doi.org/10.3390/insects12020145
Bond JG, Aguirre-Ibáñez S, Osorio AR, Marina CF, Gómez-Simuta Y, Tamayo-Escobar R, Dor A, Liedo P, Carvalho DO, Williams T. Sexual Competitiveness and Induced Egg Sterility by Aedes aegypti and Aedes albopictus Gamma-Irradiated Males: A Laboratory and Field Study in Mexico. Insects. 2021; 12(2):145. https://doi.org/10.3390/insects12020145
Chicago/Turabian StyleBond, J. Guillermo, Santiago Aguirre-Ibáñez, Adriana R. Osorio, Carlos F. Marina, Yeudiel Gómez-Simuta, Rodolfo Tamayo-Escobar, Ariane Dor, Pablo Liedo, Danilo O. Carvalho, and Trevor Williams. 2021. "Sexual Competitiveness and Induced Egg Sterility by Aedes aegypti and Aedes albopictus Gamma-Irradiated Males: A Laboratory and Field Study in Mexico" Insects 12, no. 2: 145. https://doi.org/10.3390/insects12020145