
Screening of Honey Bee Pathogens in the Czech Republic and Their Prevalence in Various Habitats

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Characterization of Different Types of Habitat
2.3. Sample Preparation and Nucleic Acid Purification
2.4. PCR Conditions
2.5. Statistical Analysis
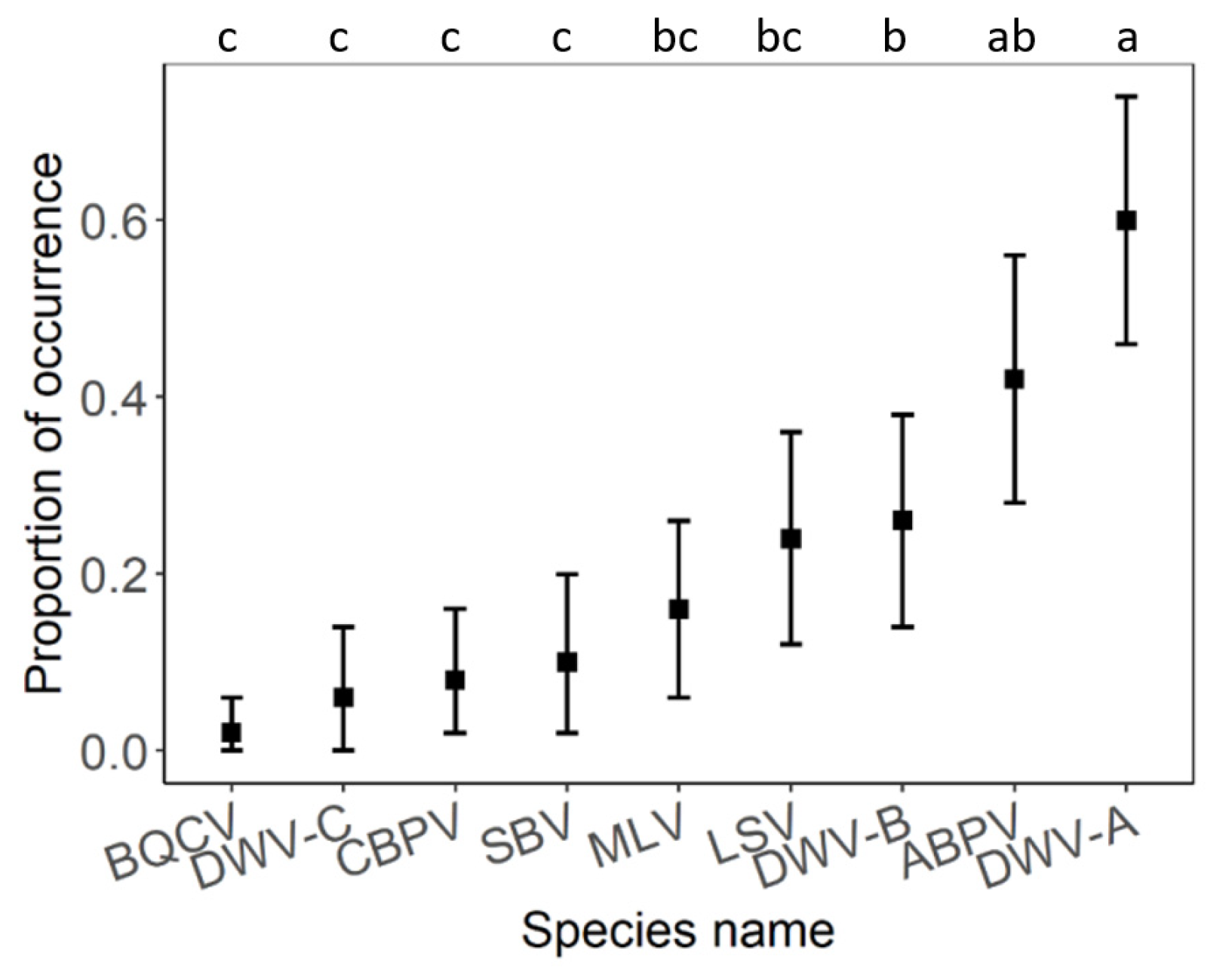
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic Valuation of the Vulnerability of World Agriculture Confronted with Pollinator Decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Stokstad, E. The Case of the Empty Hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Del Nozal, M.J.; Mayo, R.; Bernal, J.L. Honeybee Colony Collapse Due to Nosema ceranae in Professional Apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef]
- Alonso-Salces, R.M.; Cugnata, N.M.; Guaspari, E.; Pellegrini, M.C.; Aubone, I.; De Piano, F.G.; Antunez, K.; Fuselli, S.R. Natural Strategies for the Control of Paenibacillus larvae, the Causative Agent of American Foulbrood in Honey Bees: A Review. Apidologie 2017, 48, 387–400. [Google Scholar] [CrossRef]
- Frazier, M.; Mullin, C.; Frazier, J.; Ashcraft, S. What Have Pesticides Got to Do with It? Am. Bee J. 2008, 148, 521–524. [Google Scholar]
- Mutinelli, F.; Granato, A. La Sindrome Del Collasso Della Colonia (Colony Collapse Disorder) Negli USA. Aggiorn. Sulla Situaz. Attuale. Apoidea 2007, 4, 175–185. [Google Scholar]
- Sharpe, R.J.; Heyden, L.C. Honey Bee Colony Collapse Disorder Is Possibly Caused by a Dietary Pyrethrum Deficiency. Biosci. Hypotheses 2009, 2, 439–440. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet Effects on Honeybee Immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Genersch, E.; Von Der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S. The German Bee Monitoring Project: A Long Term Study to Understand Periodically High Winter Losses of Honey Bee Colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Lanzi, G.; De Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and Biological Characterization of Deformed Wing Virus of Honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordecai, G.J.; Wilfert, L.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Diversity in a Honey Bee Pathogen: First Report of a Third Master Variant of the Deformed Wing Virus Quasispecies. ISME J. 2016, 10, 1264–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Dead or Alive: Deformed Wing Virus and Varroa destructor Reduce the Life Span of Winter Honeybees. Appl. Environ. Microbiol. 2012, 78, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, D.C.; Martin, S.J. Deformed Wing Virus: The Main Suspect in Unexplained Honeybee Deaths Worldwide. Virulence 2012, 3, 589–591. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Brettell, L.E.; Martin, S.J.; Dixon, D.; Jones, I.M.; Schroeder, D.C. Superinfection Exclusion and the Long-Term Survival of Honey Bees in Varroa-Infested Colonies. ISME J. 2016, 10, 1182–1191. [Google Scholar] [CrossRef]
- Natsopoulou, M.E.; McMahon, D.P.; Doublet, V.; Frey, E.; Rosenkranz, P.; Paxton, R.J. The Virulent, Emerging Genotype B of Deformed Wing Virus Is Closely Linked to Overwinter Honeybee Worker Loss. Sci. Rep. 2017, 7, 5242. [Google Scholar] [CrossRef]
- Ongus, J.R.; Peters, D.; Bonmatin, J.-M.; Bengsch, E.; Vlak, J.M.; van Oers, M.M. Complete Sequence of a Picorna-like Virus of the Genus Iflavirus Replicating in the Mite Varroa destructor. J. Gen. Virol. 2004, 85, 3747–3755. [Google Scholar] [CrossRef]
- McMahon, D.P.; Fürst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.; Paxton, R.J. A Sting in the Spit: Widespread Cross-infection of Multiple RNA Viruses across Wild and Managed Bees. J. Anim. Ecol. 2015, 84, 615–624. [Google Scholar] [CrossRef]
- Tehel, A.; Brown, M.J.; Paxton, R.J. Impact of Managed Honey Bee Viruses on Wild Bees. Curr. Opin. Virol. 2016, 19, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Beims, H.; Bunk, B.; Erler, S.; Mohr, K.I.; Spröer, C.; Pradella, S.; Günther, G.; Rohde, M.; von der Ohe, W.; Steinert, M. Discovery of Paenibacillus larvae ERIC V: Phenotypic and Genomic Comparison to Genotypes ERIC I-IV Reveal Different Inventories of Virulence Factors Which Correlate with Epidemiological Prevalences of American Foulbrood. Int. J. Med Microbiol. 2020, 310, 151394. [Google Scholar] [CrossRef]
- Genersch, E. American Foulbrood in Honeybees and Its Causative Agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef] [PubMed]
- Reybroeck, W. Residues of Antibiotics and Chemotherapeutics in Honey. J. Apic. Res. 2018, 57, 97–112. [Google Scholar] [CrossRef]
- Forsgren, E. European Foulbrood in Honey Bees. J. Invertebr. Pathol. 2010, 103, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Lewkowski, O.; Erler, S. Virulence of Melissococcus plutonius and Secondary Invaders Associated with European Foulbrood Disease of the Honey Bee. MicrobiologyOpen 2019, 8, e00649. [Google Scholar] [CrossRef] [PubMed]
- Erban, T.; Ledvinka, O.; Kamler, M.; Hortova, B.; Nesvorna, M.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. Bacterial Community Associated with Worker Honeybees (Apis mellifera) Affected by European Foulbrood. PeerJ 2017, 5, e3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Hernández, R.; Bartolomé, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; García-Palencia, P.; Meana, A.; Pinto, M.A.; Soroker, V. Nosema ceranae in Apis mellifera: A 12 Years Postdetection Perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [Green Version]
- Spiltoir, C.F. Life Cycle of Ascosphaera apis (Pericystis Apis). Am. J. Bot. 1955, 501–508. [Google Scholar] [CrossRef]
- Mráz, P.; Hýbl, M.; Kopecký, M.; Bohatá, A.; Konopická, J.; Hoštičková, I.; Konvalina, P.; Šipoš, J.; Rost, M.; Čurn, V. The Effect of Artificial Media and Temperature on the Growth and Development of the Honey Bee Brood Pathogen Ascosphaera apis. Biology 2021, 10, 431. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Puchanichanthranon, T.; Wongsiri, S. Inhibitory Effects of Some Medicinal Plant Extracts on the Growth of Ascosphaera apis. In Proceedings of the III WOCMAP Congress on Medicinal and Aromatic Plants-Volume 4: Targeted Screening of Medicinal and Aromatic Plants, Economics 678, Chiang Mai, Thailand, 3–7 February 2003; pp. 183–189. [Google Scholar]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Khan, K.A.; Alqarni, A.S.; Kaur, M.; Al-Waili, N. In Vitro Evaluation of the Effects of Some Plant Essential Oils on Ascosphaera apis, the Causative Agent of Chalkbrood Disease. Saudi J. Biol. Sci. 2017, 24, 1001–1006. [Google Scholar] [CrossRef] [Green Version]
- Aronstein, K.A.; Murray, K.D. Chalkbrood Disease in Honey Bees. J. Invertebr. Pathol. 2010, 103, S20–S29. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Bauchan, G.R.; Murphy, C.A.; Ravoet, J.; de Graaf, D.C.; Evans, J.D. Characterization of Two Species of Trypanosomatidae from the Honey Bee Apis mellifera: Crithidia mellificae Langridge and McGhee, and Lotmaria passim n. Gen., n. Sp. J. Eukaryot. Microbiol. 2015, 62, 567–583. [Google Scholar] [CrossRef]
- Langridge, D.; McGhee, R.B. Crithidia mellificae n. Sp. an Acidophilic Trypanosomatid of the Honey Bee Apis mellifera. J. Protozool. 1967, 14, 485–487. [Google Scholar] [CrossRef]
- Vejnovic, B.; Stevanovic, J.; Schwarz, R.S.; Aleksic, N.; Mirilovic, M.; Jovanovic, N.M.; Stanimirovic, Z. Quantitative PCR Assessment of Lotmaria passim in Apis mellifera Colonies Co-Infected Naturally with Nosema ceranae. J. Invertebr. Pathol. 2018, 151, 76–81. [Google Scholar] [CrossRef]
- Ravoet, J.; Maharramov, J.; Meeus, I.; De Smet, L.; Wenseleers, T.; Smagghe, G.; De Graaf, D.C. Comprehensive Bee Pathogen Screening in Belgium Reveals Crithidia mellificae as a New Contributory Factor to Winter Mortality. PLoS ONE 2013, 8, e72443. [Google Scholar]
- Arismendi, N.; Caro, S.; Castro, M.P.; Vargas, M.; Riveros, G.; Venegas, T. Impact of Mixed Infections of Gut Parasites Lotmaria passim and Nosema ceranae on the Lifespan and Immune-Related Biomarkers in Apis mellifera. Insects 2020, 11, 420. [Google Scholar] [CrossRef] [PubMed]
- Castelli, L.; Branchiccela, B.; Invernizzi, C.; Tomasco, I.; Basualdo, M.; Rodriguez, M.; Zunino, P.; Antúnez, K. Detection of Lotmaria passim in Africanized and European Honey Bees from Uruguay, Argentina and Chile. J. Invertebr. Pathol. 2019, 160, 95–97. [Google Scholar] [CrossRef]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Hubert, J.; Bicianova, M.; Ledvinka, O.; Kamler, M.; Lester, P.J.; Nesvorna, M.; Kopecky, J.; Erban, T. Changes in the Bacteriome of Honey Bees Associated with the Parasite Varroa destructor, and Pathogens Nosema and Lotmaria passim. Microb. Ecol. 2017, 73, 685–698. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Hýbl, M.; Mráz, P.; Šipoš, J.; Hoštičková, I.; Bohatá, A.; Čurn, V.; Kopec, T. Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid. Insects 2021, 12, 572. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and Health in Honey Bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Rutter, L.; Carrillo-Tripp, J.; Bonning, B.C.; Cook, D.; Toth, A.L.; Dolezal, A.G. Transcriptomic Responses to Diet Quality and Viral Infection in Apis mellifera. BMC Genom. 2019, 20, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
- Garbuzov, M.; Schürch, R.; Ratnieks, F.L. Eating Locally: Dance Decoding Demonstrates That Urban Honey Bees in Brighton, UK, Forage Mainly in the Surrounding Urban Area. Urban Ecosyst. 2015, 18, 411–418. [Google Scholar] [CrossRef]
- Hybl, M.; Mraz, P.; Sipos, J. Diversity of Bees (Apoidea) and Their Pesticide Contamination in Two Different Types of Agricultural Management. In Proceedings of the MendelNet, Brno, Czech Republic, 11–12 November 2020; Volume 27, pp. 216–221. [Google Scholar]
- Ryba, S.; Titera, D.; Schodelbauerova-Traxmandlova, I.; Kindlmann, P. Prevalence of Honeybee Viruses in the Czech Republic and Coinfections with Other Honeybee Disease. Biologia 2012, 67, 590–595. [Google Scholar] [CrossRef]
- Bakonyi, T.; Derakhshifar, I.; Grabensteiner, E.; Nowotny, N. Development and Evaluation of PCR Assays for the Detection of Paenibacillus larvae in Honey Samples: Comparison with Isolation and Biochemical Characterization. Appl. Environ. Microbiol. 2003, 69, 1504–1510. [Google Scholar] [CrossRef] [Green Version]
- Govan, V.; Allsopp, M.; Davison, S. A PCR Detection Method for Rapid Identification of Paenibacillus larvae. Appl. Environ. Microbiol. 1999, 65, 2243–2245. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Bailón, E.; Higes, M.; Martínez-Salvador, A.; Antúnez, K.; Botías, C.; Meana, A.; Prieto, L.; Martín-Hernández, R. The Prevalence of the Honeybee Brood Pathogens A Scosphaera apis, P Aenibacillus larvae and M Elissococcus plutonius in S Panish Apiaries Determined with a New Multiplex PCR Assay. Microb. Biotechnol. 2013, 6, 731–739. [Google Scholar] [CrossRef]
- Stevanovic, J.; Schwarz, R.S.; Vejnovic, B.; Evans, J.D.; Irwin, R.E.; Glavinic, U.; Stanimirovic, Z. Species-Specific Diagnostics of Apis mellifera Trypanosomatids: A Nine-Year Survey (2007–2015) for Trypanosomatids and Microsporidians in Serbian Honey Bees. J. Invertebr. Pathol. 2016, 139, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Arismendi, N.; Bruna, A.; Zapata, N.; Vargas, M. PCR-Specific Detection of Recently Described Lotmaria passim (Trypanosomatidae) in Chilean Apiaries. J. Invertebr. Pathol. 2016, 134, 1–5. [Google Scholar] [CrossRef]
- Riveros, G.; Arismendi, N.; Zapata, N.; Evans, D.; Pérez, I.; Aldea, P.; Vargas, M. Occurrence, Prevalence and Viral Load of Deformed Wing Virus Variants in Apis mellifera Colonies in Chile. J. Apic. Res. 2020, 59, 63–68. [Google Scholar] [CrossRef]
- Sguazza, G.H.; Reynaldi, F.J.; Galosi, C.M.; Pecoraro, M.R. Simultaneous Detection of Bee Viruses by Multiplex PCR. J. Virol. Methods 2013, 194, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and Seasonal Variations of Six Bee Viruses in Apis mellifera L. and Varroa destructor Mite Populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [Green Version]
- Iwanowicz, D.D.; Wu-Smart, J.Y.; Olgun, T.; Smart, A.H.; Otto, C.R.; Lopez, D.; Evans, J.D.; Cornman, R. An Updated Genetic Marker for Detection of Lake Sinai Virus and Metagenetic Applications. PeerJ 2020, 8, e9424. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Ravallec, M.; Tournaire, M.; Cousserans, F.; Bergoin, M.; Dainat, B.; de Miranda, J.R. Viruses Associated with Ovarian Degeneration in Apis mellifera L. Queens. PLoS ONE 2011, 6, e16217. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- ter Braak, C.J.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0. 2012. Available online: https://research.wur.nl/en/publications/canoco-reference-manual-and-users-guide-software-for-ordination-v (accessed on 22 September 2021).
- Gómez-Moracho, T.; Buendía-Abad, M.; Benito, M.; García-Palencia, P.; Barrios, L.; Bartolomé, C.; Maside, X.; Meana, A.; Jiménez-Antón, M.D.; Olías-Molero, A.I. Experimental Evidence of Harmful Effects of Crithidia mellificae and Lotmaria passim on Honey Bees. Int. J. Parasitol. 2020, 50, 1117–1124. [Google Scholar] [CrossRef]
- Michalczyk, M.; Bancerz-Kisiel, A.; Sokół, R. Lotmaria passim as Third Parasite Gastrointestinal Tract of Honey Bees Living in Tree Trunk. J. Apic. Sci. 2020, 64, 143–151. [Google Scholar] [CrossRef]
- Cepero, A.; Ravoet, J.; Gómez-Moracho, T.; Bernal, J.L.; Del Nozal, M.J.; Bartolomé, C.; Maside, X.; Meana, A.; González-Porto, A.V.; de Graaf, D.C. Holistic Screening of Collapsing Honey Bee Colonies in Spain: A Case Study. BMC Res. Notes 2014, 7, 649. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; Del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G. How Natural Infection by Nosema ceranae Causes Honeybee Colony Collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.-C.; Linde, A.; Genersch, E. Five-Year Cohort Study of Nosema Spp. in Germany: Does Climate Shape Virulence and Assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevanovic, J.; Stanimirovic, Z.; Genersch, E.; Kovacevic, S.R.; Ljubenkovic, J.; Radakovic, M.; Aleksic, N. Dominance of Nosema ceranae in Honey Bees in the Balkan Countries in the Absence of Symptoms of Colony Collapse Disorder. Apidologie 2011, 42, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; Meana, A.; Bartolomé, C.; Botías, C.; Martín-Hernández, R. Nosema ceranae (Microsporidia), a Controversial 21st Century Honey Bee Pathogen. Environ. Microbiol. Rep. 2013, 5, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae Has Infected Apis mellifera in Europe since at Least 1998 and May Be More Virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Gisder, S.; Schüler, V.; Horchler, L.L.; Groth, D.; Genersch, E. Long-Term Temporal Trends of Nosema Spp. Infection Prevalence in Northeast Germany: Continuous Spread of Nosema ceranae, an Emerging Pathogen of Honey Bees (Apis mellifera), but No General Replacement of Nosema apis. Front. Cell. Infect. Microbiol. 2017, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Shumkova, R.; Georgieva, A.; Radoslavov, G.; Sirakova, D.; Dzhebir, G.; Neov, B.; Bouga, M.; Hristov, P. The First Report of the Prevalence of Nosema ceranae in Bulgaria. PeerJ 2018, 6, e4252. [Google Scholar] [CrossRef] [Green Version]
- Ostroverkhova, N.V.; Konusova, O.L.; Kucher, A.N.; Kireeva, T.N.; Rosseykina, S.A. Prevalence of the Microsporidian Nosema Spp. in Honey Bee Populations (Apis mellifera) in Some Ecological Regions of North Asia. Vet. Sci. 2020, 7, 111. [Google Scholar] [CrossRef]
- Li, Z.; You, X.; Wang, L.; Yan, Z.; Zhou, Z. Spore Morphology and Ultrastructure of an Ascosphaera apis Strain from the Honeybees (Apis mellifera) in Southwest China. Mycologia 2018, 110, 325–338. [Google Scholar] [CrossRef]
- Biová, J.; Bzdil, J.; Dostálková, S.; Petrivalsky, M.; Brus, J.; Carra, E.; Danihlík, J. American Foulbrood in the Czech Republic: ERIC II Genotype of Paenibacillus larvae Is Prevalent. Front. Vet. Sci. 2021, 8, 698976. [Google Scholar] [CrossRef]
- Erban, T.; Ledvinka, O.; Kamler, M.; Hortova, B.; Nesvorna, M.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. European Foulbrood in Czechia after 40 Years: Application of next-Generation Sequencing to Analyze Melissococcus plutonius Transmission and Influence on the Bacteriome of Apis mellifera. PeerJ Prepr. 2017, 4, e2618v1. [Google Scholar]
- Budge, G.E.; Barrett, B.; Jones, B.; Pietravalle, S.; Marris, G.; Chantawannakul, P.; Thwaites, R.; Hall, J.; Cuthbertson, A.G.; Brown, M.A. The Occurrence of Melissococcus plutonius in Healthy Colonies of Apis mellifera and the Efficacy of European Foulbrood Control Measures. J. Invertebr. Pathol. 2010, 105, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.; Franco, S. Investigation of an Atypical Case of European Foulbrood in France. Vet. Rec. Case Rep. 2021, 9, e45. [Google Scholar] [CrossRef]
- Grangier, V.; Belloy, L.; Charrière, J.-D.; Doherr, M.G.; Fritsche, A.; Waldvogel, A.S. Real-Time PCR as a Decision Aid in the Control of European Foulbrood. J. Apic. Res. 2015, 54, 366–372. [Google Scholar] [CrossRef]
- Kevill, J.L.; de Souza, F.S.; Sharples, C.; Oliver, R.; Schroeder, D.C.; Martin, S.J. DWV-A Lethal to Honey Bees (Apis mellifera): A Colony Level Survey of DWV Variants (A, B, and C) in England, Wales, and 32 States across the US. Viruses 2019, 11, 426. [Google Scholar] [CrossRef] [Green Version]
- Olgun, T.; Everhart, S.E.; Anderson, T.; Wu-Smart, J. Comparative Analysis of Viruses in Four Bee Species Collected from Agricultural, Urban, and Natural Landscapes. PLoS ONE 2020, 15, e0234431. [Google Scholar]
- McMahon, D.P.; Natsopoulou, M.E.; Doublet, V.; Fürst, M.; Weging, S.; Brown, M.J.; Gogol-Döring, A.; Paxton, R.J. Elevated Virulence of an Emerging Viral Genotype as a Driver of Honeybee Loss. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160811. [Google Scholar] [CrossRef]
- Tehel, A.; Vu, Q.; Bigot, D.; Gogol-Döring, A.; Koch, P.; Jenkins, C.; Doublet, V.; Theodorou, P.; Paxton, R. The Two Prevalent Genotypes of an Emerging Infectious Disease, Deformed Wing Virus, Cause Equally Low Pupal Mortality and Equally High Wing Deformities in Host Honey Bees. Viruses 2019, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Gisder, S.; Möckel, N.; Eisenhardt, D.; Genersch, E. In Vivo Evolution of Viral Virulence: Switching of Deformed Wing Virus between Hosts Results in Virulence Changes and Sequence Shifts. Environ. Microbiol. 2018, 20, 4612–4628. [Google Scholar] [CrossRef] [Green Version]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; Evans, J.D. Recent Spread of Varroa destructor Virus-1, a Honey Bee Pathogen, in the United States. Sci. Rep. 2017, 7, 17447. [Google Scholar] [CrossRef] [Green Version]
- Manley, R.; Temperton, B.; Doyle, T.; Gates, D.; Hedges, S.; Boots, M.; Wilfert, L. Knock-on Community Impacts of a Novel Vector: Spillover of Emerging DWV-B from Varroa-infested Honeybees to Wild Bumblebees. Ecol. Lett. 2019, 22, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Kevill, J.L.; Highfield, A.; Mordecai, G.J.; Martin, S.J.; Schroeder, D.C. ABC Assay: Method Development and Application to Quantify the Role of Three DWV Master Variants in Overwinter Colony Losses of European Honey Bees. Viruses 2017, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, H.; Imdorf, A.; Haueter, M.; Radloff, S.; Neumann, P. Virus Infections and Winter Losses of Honey Bee Colonies (Apis mellifera). J. Apic. Res. 2010, 49, 60–65. [Google Scholar] [CrossRef]
- Nguyen, B.K.; Ribière, M.; vanEngelsdorp, D.; Snoeck, C.; Saegerman, C.; Kalkstein, A.L.; Schurr, F.; Brostaux, Y.; Faucon, J.-P.; Haubruge, E. Effects of Honey Bee Virus Prevalence, Varroa destructor Load and Queen Condition on Honey Bee Colony Survival over the Winter in Belgium. J. Apic. Res. 2011, 50, 195–202. [Google Scholar] [CrossRef]
- Faurot-Daniels, C.; Glenny, W.; Daughenbaugh, K.F.; McMenamin, A.J.; Burkle, L.A.; Flenniken, M.L. Longitudinal Monitoring of Honey Bee Colonies Reveals Dynamic Nature of Virus Abundance and Indicates a Negative Impact of Lake Sinai Virus 2 on Colony Health. PLoS ONE 2020, 15, e0237544. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Cornman, R.S.; Evans, J.D.; Semberg, E.; Haddad, N.; Neumann, P.; Gauthier, L. Genome Characterization, Prevalence and Distribution of a Macula-like Virus from Apis mellifera and Varroa destructor. Viruses 2015, 7, 3586–3602. [Google Scholar] [CrossRef] [Green Version]
- McMenamin, A.J.; Flenniken, M.L. Recently Identified Bee Viruses and Their Impact on Bee Pollinators. Curr. Opin. Insect Sci. 2018, 26, 120–129. [Google Scholar] [CrossRef]
- Abou Kubaa, R.; Molinatto, G.; Khaled, B.; Daher-Hjaij, N.; Heinoun, K.; Saponari, M. First Detection of Black Queen Cell Virus, Varroa destructor Macula-like Virus, Apis mellifera Filamentous Virus and Nosema ceranae in Syrian Honey Bees Apis mellifera Syriaca. Bull. Insectol. 2018, 71, 217–224. [Google Scholar]
- von Büren, R.S.; Oehen, B.; Kuhn, N.J.; Erler, S. High-Resolution Maps of Swiss Apiaries and Their Applicability to Study Spatial Distribution of Bacterial Honey Bee Brood Diseases. PeerJ 2019, 7, e6393. [Google Scholar] [CrossRef]
- Brodschneider, R.; Brus, J.; Danihlík, J. Comparison of Apiculture and Winter Mortality of Honey Bee Colonies (Apis mellifera) in Austria and Czechia. Agric. Ecosyst. Environ. 2019, 274, 24–32. [Google Scholar] [CrossRef]
- Taric, E.; Glavinic, U.; Vejnovic, B.; Stanojkovic, A.; Aleksic, N.; Dimitrijevic, V.; Stanimirovic, Z. Oxidative Stress, Endoparasite Prevalence and Social Immunity in Bee Colonies Kept Traditionally vs. Those Kept for Commercial Purposes. Insects 2020, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Tehel, A.; Streicher, T.; Tragust, S.; Paxton, R.J. Experimental Infection of Bumblebees with Honeybee-Associated Viruses: No Direct Fitness Costs but Potential Future Threats to Novel Wild Bee Hosts. R. Soc. Open Sci. 2020, 7, 200480. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Li-Byarlay, H.; Huang, M.H.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Migratory Management and Environmental Conditions Affect Lifespan and Oxidative Stress in Honey Bees. Sci. Rep. 2016, 6, 32023. [Google Scholar] [CrossRef] [PubMed]
- Graystock, P.; Ng, W.H.; Parks, K.; Tripodi, A.D.; Muñiz, P.A.; Fersch, A.A.; Myers, C.R.; McFrederick, Q.S.; McArt, S.H. Dominant Bee Species and Floral Abundance Drive Parasite Temporal Dynamics in Plant-Pollinator Communities. Nat. Ecol. Evol. 2020, 4, 1358–1367. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences 5′-3′ | TA [°C] | Reference |
|---|---|---|---|
| Nosema apis | F: GGGGGCATGTCTTTGACGTACTATGTA R: GGGGGGCGTTTAAAATGTGAAACAACTATG | 62 | [48] |
| Nosema ceranae | F: CGGCGACGATGTGATATGAAAATATTAA R: CCCGGTCATTCTCAAACAAAAAACCG | 62 | [48] |
| Paenibacillus larvae | F: GCTCTGTTGCCAAGGAAGAA R: AGGCGGAATGCTTACTGTGT | 55 | [49] |
| Melissococcus plutonius | F: GAAGAGGAGTTAAAAGGCGC R: TTATCTCTAAGGCGTTCAAAGG | 55 | [50] |
| Ascosphaera apis | F: TGTGTCTGTGCGGCTAGGTG R: GCTAGCCAGGGGGGAACTAA | 60 | [51] |
| Crithidia mellificae | F: AGTTTGAGCTGTTGGATTTGTT R: AACCTATTACAGGCACAGTTGC | 56 | [52] |
| Lotmaria passim | F: TGACTTGAATTAGCAAGCATGGGATAACA R: CCTTTAGGCTACCGTTTCGGCTTTTGTTGGT | 60 | [53] |
| DWV-A | F: CGTCGGCCTATCAAAG R: CTTTTCTAATTCAACTTCACC | 60 | [54] |
| DWV-B | F: GCCCTGTTCAAGAACATG R: CTTTTCTAATTCAACTTCACC | 60 | [54] |
| DWV-C | F: TACTAGTGCTGGTTTTCCTTT R: ATAAGTTGCGTGGTTGAC | 60 | [54] |
| BQCV | F: GGACGAAAGGAAGCCTAAAC R: ACTAGGAAGAGACTTGCACC | 48 | [48] |
| CBPV | F: AACCTGCCTCAACACAGGCAAC R: ACATCTCTTCTTCGGTGTCAGCC | 60 | [55] |
| ABPV | F: TGAGAACACCTGTAATGTGG R: ACCAGAGGGTTGACTGTGTG | 48 | [56] |
| SBV | F: GGATGAAAGGAAATTACCAG R: CCACTAGGTGATCCACACT | 48 | [56] |
| LSV | F: CKTGCGGNCCTCATTTCTTCATGTC R: CATGAATCCAAKGTCAAAGGTRTCGT | 60 | [57] |
| MLV | F: ATCCCTTTTCAGTTCGCT R: AGAAGAGACTTCAAGGAC | 60 | [58] |
| Df. | AIC | LRT | Pr (Chi) | |
|---|---|---|---|---|
| Dependent variable: species occurrence | ||||
| Full model | 243.68 | |||
| Eukaryote | 4 | 332.25 | 96.570 | <0.0001 |
| Habitat | 2 | 246.76 | 7.081 | 0.02899 |
| Full model | 398.26 | |||
| Virus | 9 | 494.96 | 114.695 | <0.0001 |
| Habitat | 2 | 396.69 | 2.423 | 0.2977 |
| Dependent variable: number of eukaryotic species | ||||
| Full model | 156.78 | |||
| Habitat | 2 | 157.23 | 4.453 | 0.107 |
| Dependent variable: number of virus types | ||||
| Full model | 181.88 | |||
| Habitat | 2 | 180.52 | 2.642 | 0.267 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mráz, P.; Hýbl, M.; Kopecký, M.; Bohatá, A.; Hoštičková, I.; Šipoš, J.; Vočadlová, K.; Čurn, V. Screening of Honey Bee Pathogens in the Czech Republic and Their Prevalence in Various Habitats. Insects 2021, 12, 1051. https://doi.org/10.3390/insects12121051
Mráz P, Hýbl M, Kopecký M, Bohatá A, Hoštičková I, Šipoš J, Vočadlová K, Čurn V. Screening of Honey Bee Pathogens in the Czech Republic and Their Prevalence in Various Habitats. Insects. 2021; 12(12):1051. https://doi.org/10.3390/insects12121051
Chicago/Turabian StyleMráz, Petr, Marian Hýbl, Marek Kopecký, Andrea Bohatá, Irena Hoštičková, Jan Šipoš, Kateřina Vočadlová, and Vladislav Čurn. 2021. "Screening of Honey Bee Pathogens in the Czech Republic and Their Prevalence in Various Habitats" Insects 12, no. 12: 1051. https://doi.org/10.3390/insects12121051