
Expansion of CRISPR Targeting Sites Using an Integrated Gene-Editing System in Apis mellifera


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis of Targeting Sites
2.2. Collection of Honeybee Embryos
2.3. In Vitro Transcription of sgRNA/crRNA and Preparation of CRISPR Protein
2.4. Microinjection and Rearing of Embryos
2.5. Genomic DNA Isolation and Identification of Amyellow-y Mutations
3. Results
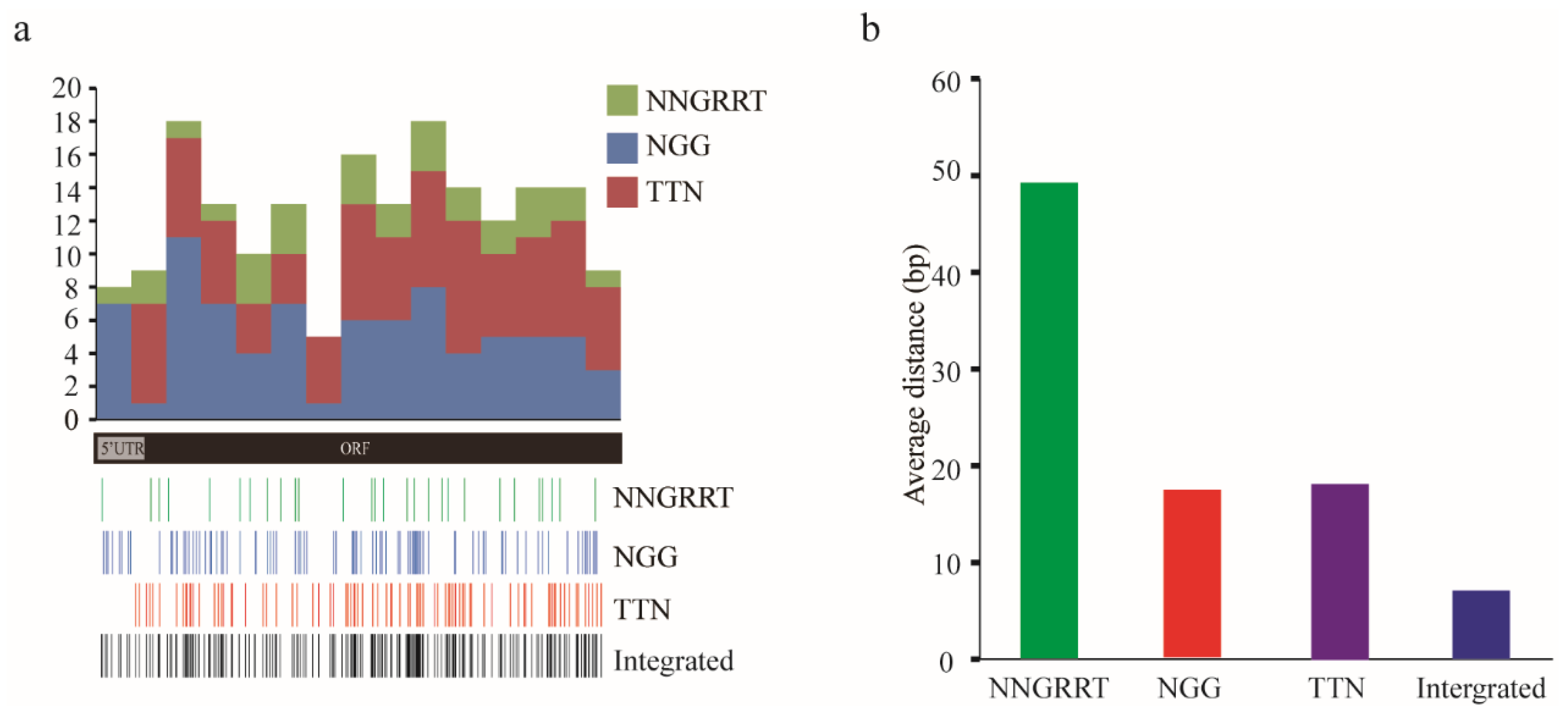
3.1. Comparative Analysis of Targetable Sites on Amyellow-y mRNA
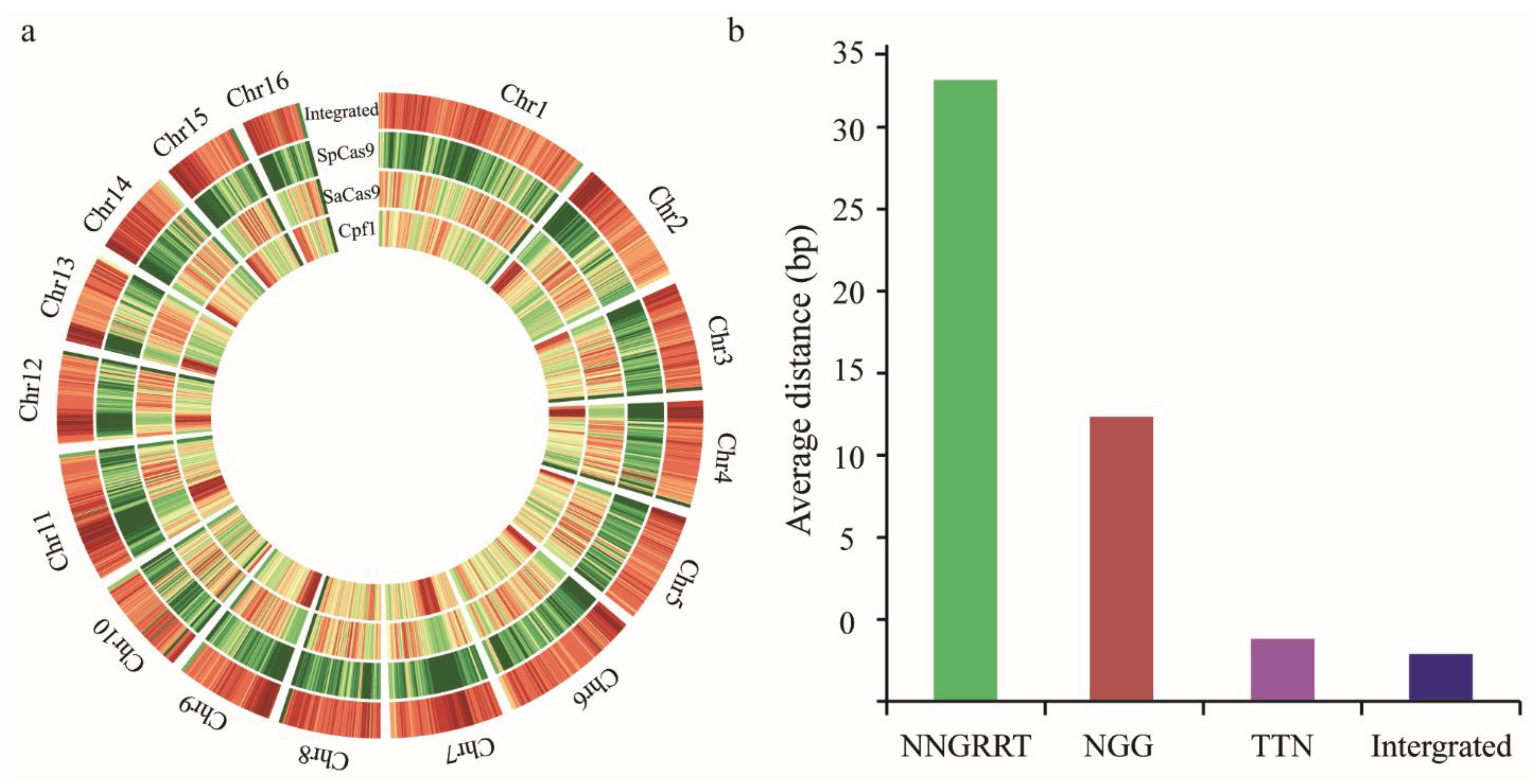
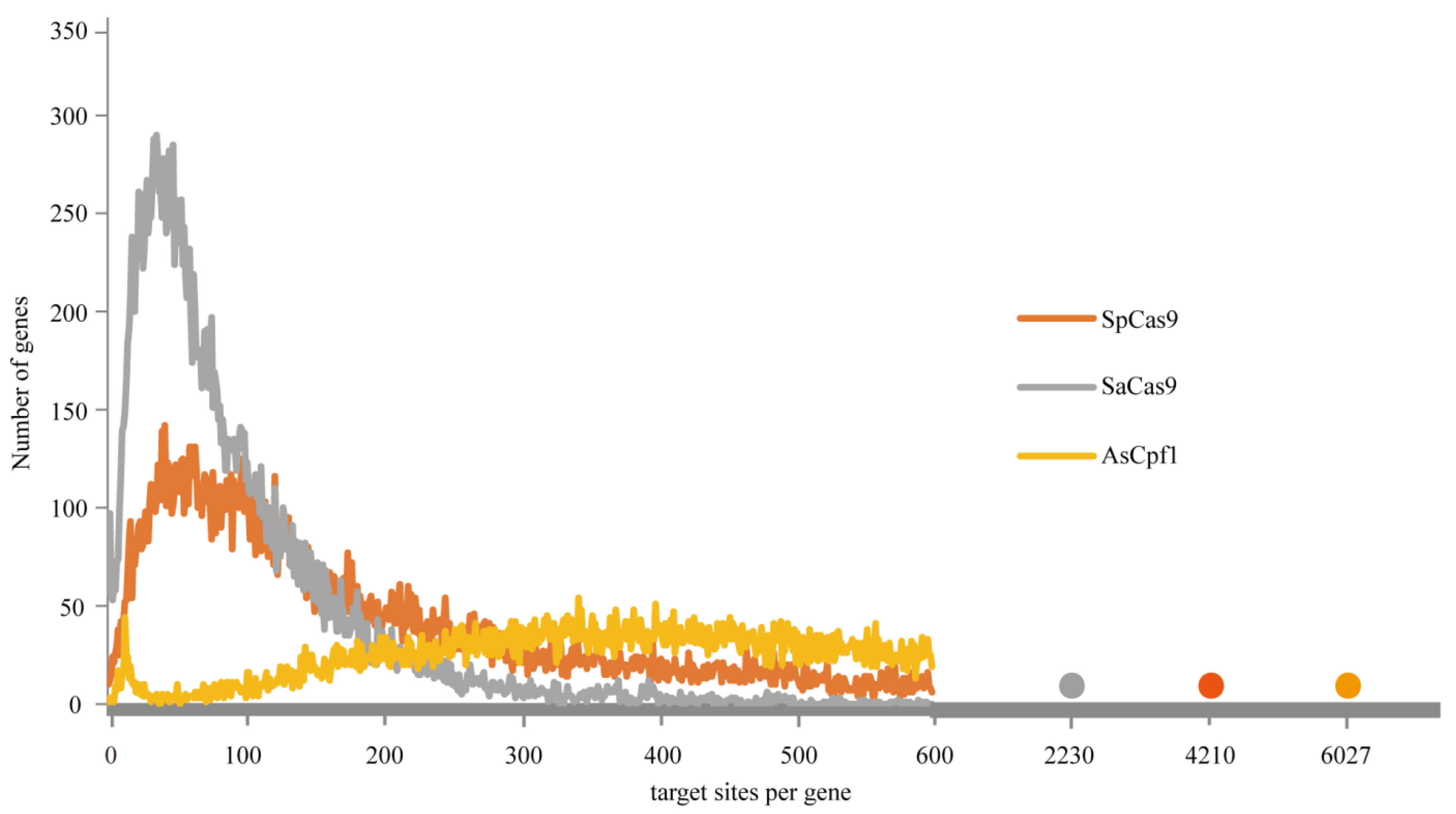
3.2. Bioinformatics Analysis of Targetable Sites on A. mellifera Genome
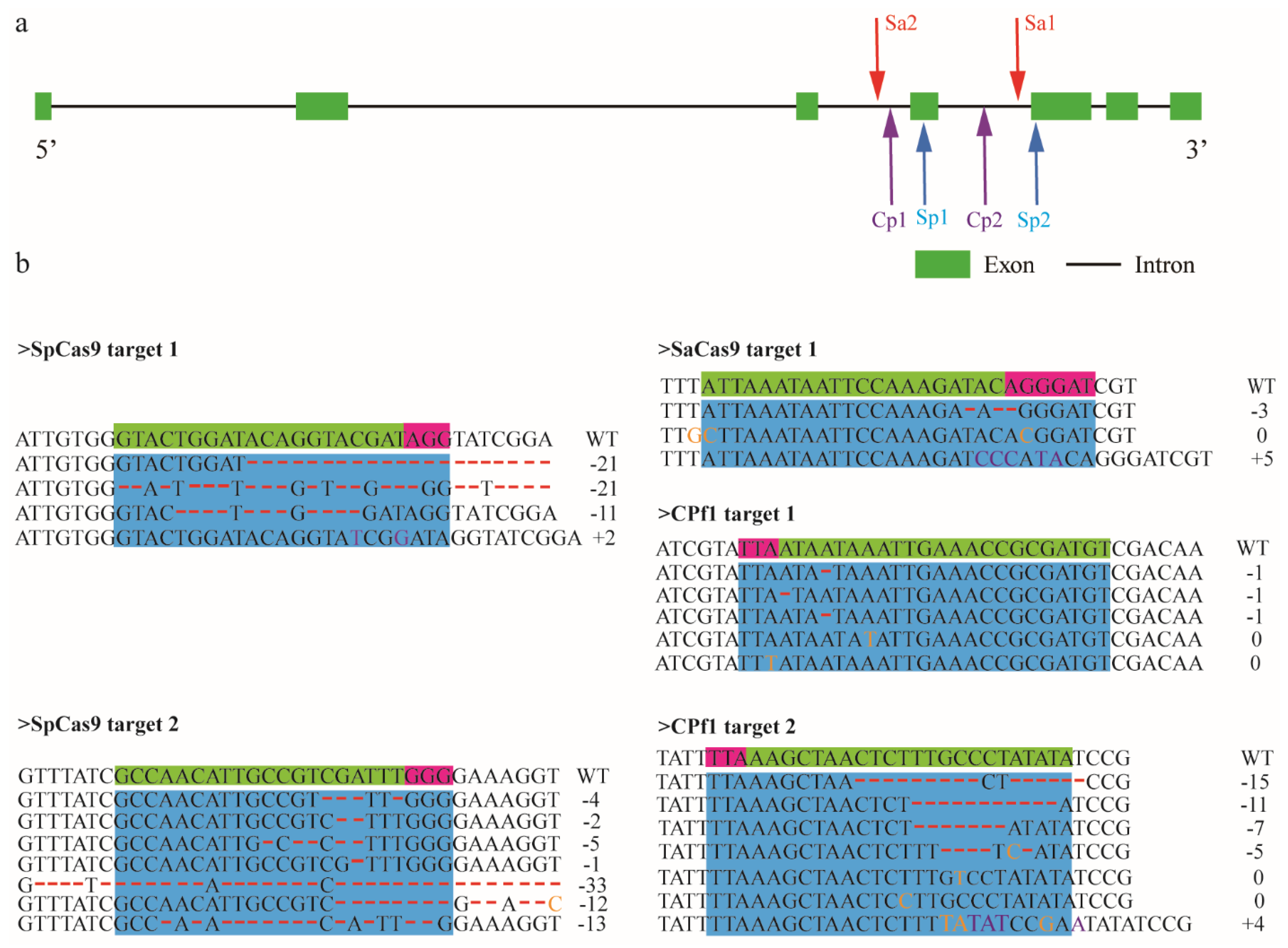
3.3. Genome Editing Mediated by SpCas9, SaCas9 and Cpf1 in A. mellifera
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, D.; Guo, Z.J.; Liu, Y.; Zhang, Y.J. Progress and prospects of CRISPR/Cas Systems in Insects and Other Arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef]
- Kohno, H.; Suenami, S.; Takeuchi, H.; Sasaki, T.; Kubo, T. Production of Knockout Mutants by CRISPR/Cas9 in the European Honeybee, Apis mellifera L. Zool Sci. 2016, 33, 505–512. [Google Scholar] [CrossRef]
- Roth, A.; Vleurinck, C.; Netschitailo, O.; Bauer, V.; Otte, M.; Kaftanoglu, O.; Page, R.E.; Beye, M. A genetic switch for worker nutrition-mediated traits in honeybees. PLoS Biol. 2019, 17, e3000171. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.F.; Zhang, B.; Liao, C.H.; Zeng, Z.J. High-efficiency CRISPR/Cas9-mediated gene editing in honeybee (Apis mellifera) embryos. G3 Genes Genomes Genet. 2019, 9, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Q.; Traniello, I.M.; Rana, S.; Cash-Ahmed, A.C.; Sankey, A.L.; Yang, C.; Robinson, G.E. Neurodevelopmental and transcriptomic effects of CRISPR/Cas9-induced somatic orco mutation in honey bees. J. Neurogenet. 2021, 35, 320–332. [Google Scholar] [CrossRef]
- Değirmenci, L.; Geiger, D.; Rogé Ferreira, F.L.; Keller, A.; Krischke, B.; Beye, M.; Steffan-Dewenter, I.; Scheiner, R. CRISPR/Cas 9-mediated mutations as a new tool for studying taste in honeybees. Chem. Senses 2020, 45, 655–666. [Google Scholar] [CrossRef]
- Nie, H.Y.; Liang, L.Q.; Li, Q.F.; Li, Z.H.Q.; Su, S.K. CRISPR/Cas9 mediated knockout of Amyellow-y gene results in melanization defect of the cuticle in adult Apis mellifera. J. Insect Physiol. 2021, 132, 104264. [Google Scholar] [CrossRef]
- Weinstock, G.M.; Robinson, G.E.; Gibbs, R.A.; Worley, K.C.; Evans, J.D.; Maleszka, R.; Robertson, H.M.; Weaver, D.B.; Beye, M.; Bork, P. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar]
- Chen, L.; Tang, L.Y.; Xiang, H.; Jin, L.J.; Li, Q.Y.; Dong, Y.; Wang, W.; Zhang, G.J. Advances in genome editing technology and its promising application in evolutionary and ecological studies. Gigascience 2014, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.S.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S. In vivo genome editing using Staphylococcus aureus Cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef]
- Zetsche, B.; Heidenreich, M.; Mohanraju, P.; Fedorova, I.; Kneppers, J.; DeGennaro, E.M.; Winblad, N.; Choudhury, S.R.; Abudayyeh, O.O.; Gootenberg, J.S. Multiplex gene editing by CRISPR–Cpf1 using a single crRNA array. Nat. Biotechnol. 2017, 35, 31–34. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Nishimasu, H.; Shi, X.; Ishiguro, S.; Gao, L.; Hirano, S.; Okazaki, S.; Noda, T.; Abudayyeh, O.O.; Gootenberg, J.S.; Mori, H. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 2018, 361, 1259–1262. [Google Scholar] [CrossRef]
- Hu, J.H.; Miller, S.M.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [Google Scholar] [CrossRef]
- Hirano, S.; Nishimasu, H.; Ishitani, R.; Nureki, O. Structural basis for the altered PAM specificities of engineered CRISPR-Cas9. Mol. Cell 2016, 61, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Kleinstiver, B.P.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Topkar, V.V.; Zheng, Z.; Joung, J.K. Broadening the targeting range of Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition. Nat. Biotechnol. 2015, 33, 1293–1298. [Google Scholar] [CrossRef]
- Wallberg, A.; Bunikis, I.; Pettersson, O.V.; Mosbech, M.-B.; Childers, A.K.; Evans, J.D.; Mikheyev, A.S.; Robertson, H.M.; Robinson, G.E.; Webster, M.T. A hybrid de novo genome assembly of the honeybee, Apis mellifera, with chromosome-length scaffolds. BMC Genom. 2019, 20, 275. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.P.; Vejnar, C.E.; Giraldez, A.J.; Romain, R.; Moreno-Mateos, M.A. Optimized CRISPR-Cpf1 system for genome editing in zebrafish. Methods 2018, 150, 11–18. [Google Scholar] [CrossRef]
- Schulte, C.; Theilenberg, E.; Müller-Borg, M.; Gempe, T.; Beye, M. Highly efficient integration and expression of piggyBac-derived cassettes in the honeybee (Apis mellifera). Proc. Natl. Acad. Sci. USA 2014, 111, 9003–9008. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.L.; Xu, J.; He, L.; Zhang, Y.; Li, K.; Huang, Y.P. CRISPR/Cas9—mediated ebony knockout results in puparium melanism in Spodoptera litura. Insect Sci. 2019, 26, 1011–1019. [Google Scholar] [CrossRef]
- Jiang, W.Y.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef]
- Ma, S.Y.; Chang, J.S.; Wang, X.G.; Liu, Y.Y.; Zhang, J.D.; Lu, W.; Gao, J.; Shi, R.; Zhao, P.; Xia, Q.Y. CRISPR/Cas9 mediated multiplex genome editing and heritable mutagenesis of BmKu70 in Bombyx mori. Sci. Rep. 2014, 4, 4489. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.Y.; Liu, Y.Y.; Liu, Y.; Chang, J.S.; Zhang, T.; Wang, X.G.; Shi, R.; Lu, W.; Xia, X.J.; Zhao, P. An integrated CRISPR Bombyx mori genome editing system with improved efficiency and expanded target sites. Insect Biochem. Mol. Biol. 2017, 83, 13–20. [Google Scholar] [CrossRef]
- Xue, W.H.; Xu, N.; Yuan, X.B.; Chen, H.H.; Zhang, J.L.; Fu, S.J.; Zhang, C.X.; Xu, H.J. CRISPR/Cas9-mediated knockout of two eye pigmentation genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2018, 93, 19–26. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, C.; Han, Y.X.; Chen, Z.L.; Lu, X.C.; Liang, F.; Li, S.; Qin, W.; Lin, S. Expanding CRISPR/Cas9 genome editing capacity in zebrafish using SaCas9. G3 Genes Genomes Genet. 2017, 6, 2517–2521. [Google Scholar] [CrossRef] [Green Version]
- Port, F.; Bullock, S.L. Augmenting CRISPR applications in Drosophila with tRNA-flanked sgRNAs. Nat. Methods 2016, 13, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Jia, G.X.; Choi, J.; Ma, H.M.; Anaya, E.; Ye, C.T.; Shankar, P.; Wu, H.Q. Optimizing sgRNA structure to improve CRISPR-Cas9 knockout efficiency. Genome Biol. 2015, 16, 280. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wang, Z.X.; Xiao, A.; Zhang, Y.T.; Li, W.Y.; Zu, Y.; Yao, S.H.; Lin, S.; Zhang, B. Efficient gene targeting in zebrafish mediated by a zebrafish-codon-optimized cas9 and evaluation of off-targeting effect. J. Genet. Genom. 2014, 41, 43–46. [Google Scholar] [CrossRef]
- Li, M.; Au, L.Y.C.; Douglah, D.; Chong, A.; White, B.J.; Ferree, P.M.; Akbari, O.S. Generation of heritable germline mutations in the jewel wasp Nasonia vitripennis using CRISPR/Cas9. Sci. Rep. 2017, 7, 901. [Google Scholar] [CrossRef] [Green Version]
- Bassett, A.R.; Tibbit, C.; Ponting, C.P.; Liu, J.L. Highly efficient targeted mutagenesis of Drosophila with the CRISPR/Cas9 system. Cell Rep. 2013, 4, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Ni, X.Y.; Zhou, Z.D.; Huang, J.; Qiao, X.M. Targeted gene disruption by CRISPR/xCas9 system in Drosophila melanogaster. Arch. Insect Biochem. Physiol. 2020, 104, e21662. [Google Scholar] [CrossRef]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- Taning, C.; Eynde, B.V.; Yu, N.; Ma, S.Y.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Kleinstiver, B.P.; Tsai, S.Q.; Prew, M.S.; Nguyen, N.T.; Welch, M.M.; Lopez, J.M.; McCaw, Z.R.; Aryee, M.J.; Joung, J.K. Genome-wide specificities of CRISPR-Cas Cpf1 nucleases in human cells. Nat. Biotechnol. 2016, 34, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, J.; Hur, J.K.; Been, K.W.; Yoon, S.-H.; Kim, J.-S. Genome-wide analysis reveals specificities of Cpf1 endonucleases in human cells. Nat. Biotechnol. 2016, 34, 863–868. [Google Scholar] [CrossRef]
- Müller, M.; Lee, C.M.; Gasiunas, G.; Davis, T.H.; Cradick, T.J.; Siksnys, V.; Bao, G.; Cathomen, T.; Mussolino, C. Streptococcus thermophilus CRISPR-Cas9 systems enable specific editing of the human genome. Mol. Ther. 2016, 24, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.F.; Sander, J.D.; Reyon, D.; Cascio, V.M.; Joung, J.K. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat. Biotechnol. 2014, 32, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.J.; Yang, Z.H.; Mao, D.C.; Chang, Z.; Qiao, H.H.; Wang, X.; Sun, J.; Hu, Q.; Cui, Y.; Liu, L.P. Performance of the Cas9 Nickase System in Drosophila melanogaster. G3 Genes Genomes Genet. 2014, 4, 1955–1962. [Google Scholar] [CrossRef] [Green Version]
- Kleinstiver, B.P.; Pattanayak, V.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Zheng, Z.; Joung, J.K. High-fidelity CRISPR–Cas9 nucleases with no detectable genome-wide off-target effects. Nature 2016, 529, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Dagdas, Y.S.; Kleinstiver, B.P.; Welch, M.M.; Sousa, A.A.; Harrington, L.B.; Sternberg, S.H.; Joung, J.K.; Yildiz, A.; Doudna, J.A. Enhanced proofreading governs CRISPR–Cas9 targeting accuracy. Nature 2017, 550, 407–410. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, L.; Li, Z.; Li, Q.; Wang, X.; Su, S.; Nie, H. Expansion of CRISPR Targeting Sites Using an Integrated Gene-Editing System in Apis mellifera. Insects 2021, 12, 954. https://doi.org/10.3390/insects12100954
Liang L, Li Z, Li Q, Wang X, Su S, Nie H. Expansion of CRISPR Targeting Sites Using an Integrated Gene-Editing System in Apis mellifera. Insects. 2021; 12(10):954. https://doi.org/10.3390/insects12100954
Chicago/Turabian StyleLiang, Liqiang, Zhenghanqing Li, Qiufang Li, Xiuxiu Wang, Songkun Su, and Hongyi Nie. 2021. "Expansion of CRISPR Targeting Sites Using an Integrated Gene-Editing System in Apis mellifera" Insects 12, no. 10: 954. https://doi.org/10.3390/insects12100954