
A Link between Atmospheric Pressure and Fertility of Drosophila Laboratory Strains




{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Melanogaster Strains and Rearing
2.2. Fertility Analysis
2.3. Meteorological Data Analysis
2.4. Statistical Analysis
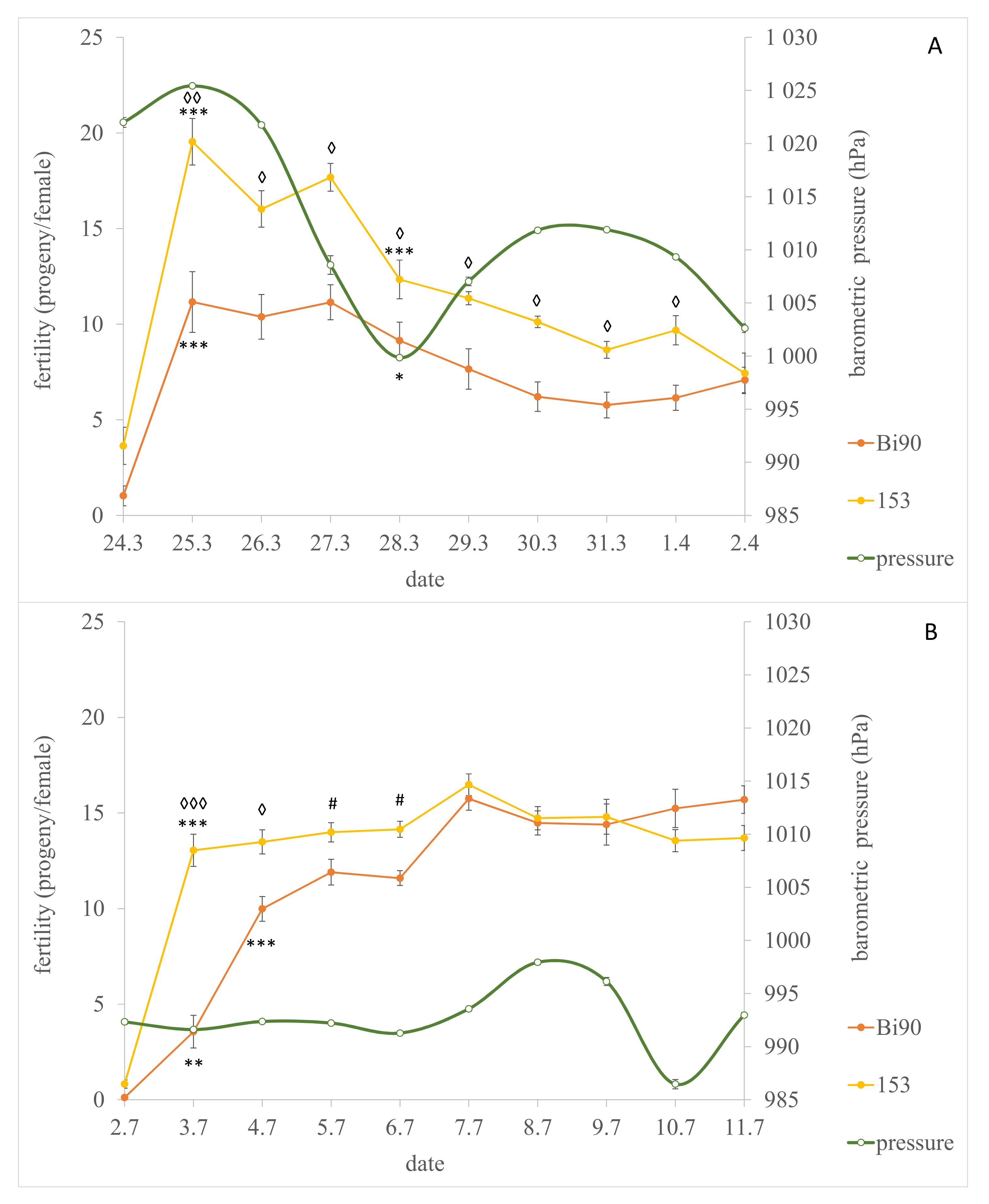
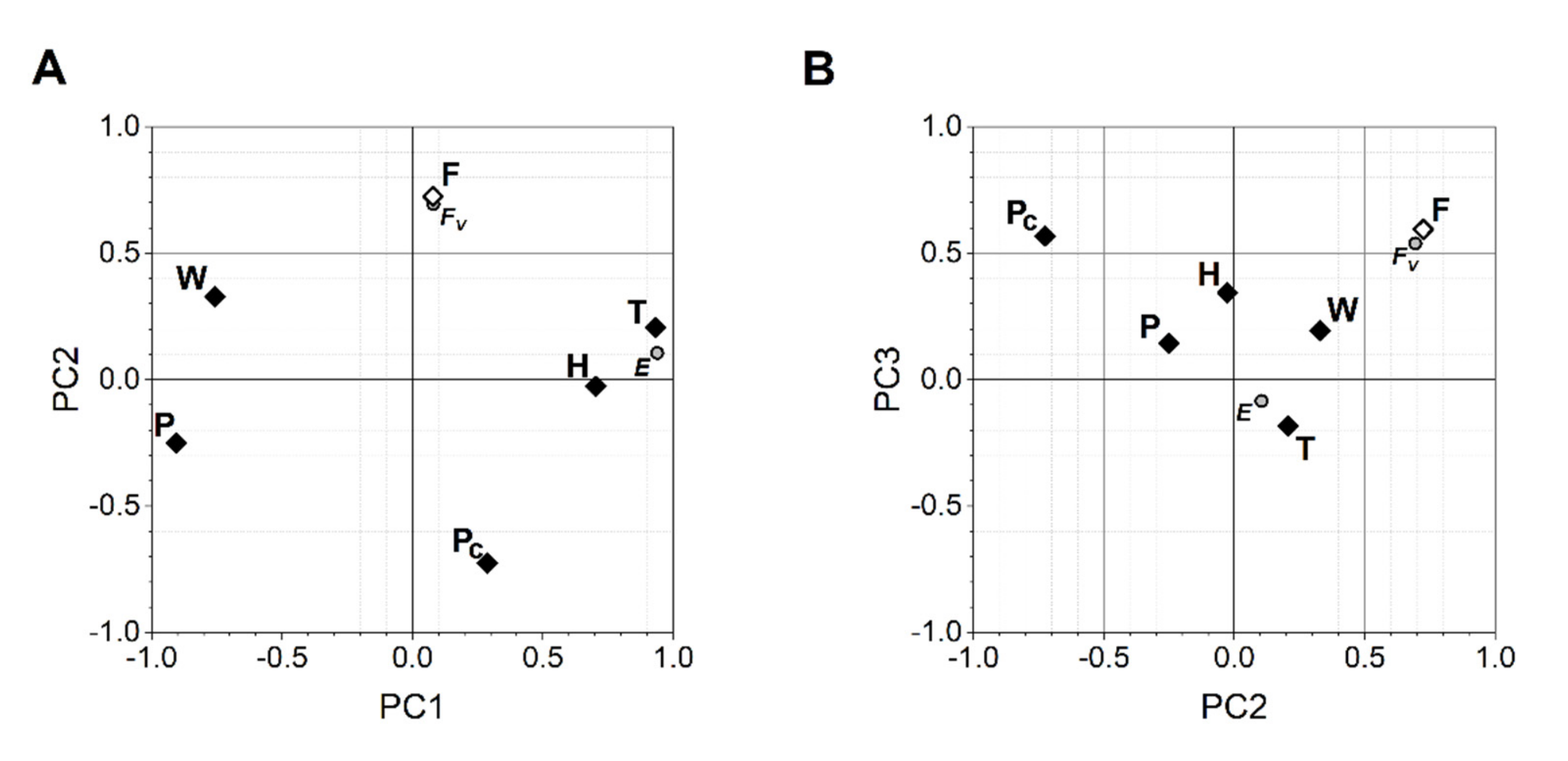
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change. Available online: https://www.ipcc.ch/sr15/ (accessed on 12 October 2021).
- Gillett, N.P.; Zwiers, F.W.; Weaver, A.J.; Stott, P.A. Detection of human influence on sea-level pressure. Nature 2003, 422, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Schaller, N.; Kay, A.; Lamb, R.; Massey, N.R.; van Oldenborgh, G.J.; Otto, F.E.L.; Sparrow, S.N.; Vautard, R.; Yiou, P.; Ashpole, I.; et al. Human influence on climate in the 2014 southern England winter floods and their impacts. Nat. Clim. Chang. 2016, 6, 627–634. [Google Scholar] [CrossRef]
- Guan, W.; Clay, S.J.; Sloan, G.J.; Pretlow, L.G. Effects of barometric pressure and temperature on acute ischemic stroke hospitalization in Augusta, GA. Transl. Stroke Res. 2018, 10, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Lickiewicz, J.; Piotrowicz, K.; Hughes, P.P.; Makara-Studzińska, M. Weather and aggressive behavior among patients in psychiatric hospitals-an exploratory study. Int. J. Environ. Res. Public Health 2020, 17, 9121. [Google Scholar] [CrossRef]
- Fagerlund, A.J.; Iversen, M.; Ekeland, A.; Moen, C.M.; Aslaksen, P.M. Blame it on the weather? The association between pain in fibromyalgia, relative humidity, temperature and barometric pressure. PLoS ONE 2019, 14, e0216902. [Google Scholar] [CrossRef] [Green Version]
- Leskey, T.; CPropoky, R.J. Influence of barometric pressure on odor discrimination and oviposition by adult plum curculios (Coleoptera: Curculionidae). Eur. J. Entomol. 2003, 100, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Fournier, F.; Pelletier, D.; Vigneault, C.; Boivin, G. Effect of barometric pressure on flight initiation by Trichogramma pretiosum and Trichogramma evanescens (Hymenoptera: Trichogrammatidae). Environ. Entomol. 2005, 34, 1534–1540. [Google Scholar] [CrossRef] [Green Version]
- Pellegrino, A.C.; Penaflor, M.F.G.V.; Nardi, C.; Bezner-Kerr, W.; Guglielmo, C.G.; Bento, J.M.S.; McNeil, J.N. Weather forecasting by insects: Modified sexual behaviour in response to atmospheric pressure changes. PLoS ONE 2013, 8, e75004. [Google Scholar] [CrossRef] [PubMed]
- Marchand, D.; McNeil, J.N. Effects of wind speed and atmospheric pressure on mate searching behavior in the aphid parasitoid Aphidius nigripes (Hymenoptera: Aphidiidae). J. Insect Behav. 2000, 13, 187–199. [Google Scholar] [CrossRef]
- Steinberg, S.; Dicke, M.; Vet, L.E.M.; Wanningen, R. Response of the braconid parasitoid Cotesia (=Apanteles) glomerata to volatile infochemicals: Effects of bioassay set-up, parasitoid age and experience and barometric flux. Entomologia Experimentalis et Applicata 1992, 63, 163–175. [Google Scholar] [CrossRef]
- McFarlane, D.J.; Rafter, M.A.; Booth, D.T.; Walter, G.H. Behavioral responses of a tiny insect, the flower thrips Frankliniella schultzei trybom (Thysanoptera, Thripidae), to atmospheric pressure change. J. Insect Behav. 2015, 1, e9. [Google Scholar] [CrossRef]
- Martini, X.; Rivera, M.; Hoyte, A.; Sétamou, M.; Stelinski, L. Effects of wind, temperature, and barometric pressure on asian citrus psyllid (Hemiptera: Liviidae) flight behavior. J. Econ. Entomol. 2018, 111, 2570–2577. [Google Scholar] [CrossRef] [PubMed]
- Ankney, P.F. A note on barometric pressure and behavior in Drosophila pseudoobscura. Behav. Genet. 1984, 14, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.J.; Guglielmo, C.G.; Moehring, A.J. A direct test of the effects of changing atmospheric pressure on the mating behavior of Drosophila melanogaster. Evol. Ecol. 2014, 28, 535–544. [Google Scholar] [CrossRef]
- Dagaeff, A.C.; Pocheville, A.; Nöbel, S.; Loyau, A.; Isabel, G.; Danchin, E. Drosophila mate copying correlates with atmospheric pressure in a speed learning situation. Anim. Behav. 2016, 121, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Izmaylov, D.M.; Obukhova, L.K.; Konradov, A.A. Correlations of life-span variation parameters in 128 successive generations of Drosophila melanogaster with changes in atmospheric pressure and geomagnetic activity. Int. J. Biometeorol. 2005, 49, 337–344. [Google Scholar] [CrossRef]
- Bosco, G.; Clamer, M.; Messulam, E.; Dare, C.; Yang, Z.; Zordan, M.; Reggiani, C.; Hu, Q.; Megighian, A. Effects of oxygen concentration and pressure on Drosophila melanogaster: Oxidative stress, mitochondrial activity, and survivorship. Arch. Insect Biochem. Physiol. 2015, 88, 222–234. [Google Scholar] [CrossRef]
- Dillon, M.E.; Frazier, M.R. Drosophila locomotion in cold, thin air. J. Exp. Biol. 2006, 209, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Gruntenko, N.E.; Rauschenbach, I.Y. Interplay of JH, 20E and biogenic amines under normal and stress conditions and its effect on reproduction. J. Insect Physiol. 2008, 54, 902–908. [Google Scholar] [CrossRef]
- Gruntenko, N.Е.; Rauschenbach, I.Y. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A mini-review. Gen. Comp. Endocrinol. 2018, 258, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.M.; Hoi, J.T.; Eissenberg, J.C.; Shoemaker, J.D.; Neckameyer, W.S.; Ilvarsonn, A.M.; Harshman, L.G.; Schlegel, V.L.; Zempleni, J. Feeding Drosophila a biotin-deficient diet for multiple generations increases stress resistance and lifespan and alters gene expression and histone biotinylation patterns. J. Nutr. 2007, 137, 2006–2012. [Google Scholar] [CrossRef] [Green Version]
- Le Bourg, E. Neglecting larval rearing conditions in Drosophila melanogaster can negatively impact research on ageing. Biogerontology 2021, 22, 369–373. [Google Scholar] [CrossRef] [PubMed]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Karpova, E.K.; Komyshev, E.G.; Genaev, M.A.; Adonyeva, N.V.; Afonnikov, D.A.; Eremina, M.A.; Gruntenko, N.E. Quantifying Drosophila adults with the use of a smartphone. Biol. Open 2020, 9, bio054452. [Google Scholar] [CrossRef]
- Bloch Qazi, M.C.; Heifetz, Y.; Wolfner, M.F. The developments between gametogenesis and fertilization: Ovulation and female sperm storage in Drosophila melanogaster. Dev. Biol. 2003, 256, 195–211. [Google Scholar] [CrossRef] [Green Version]
- Gruntenko, N.E.; Chentsova, N.A.; Andreenkova, E.V.; Bownes, M.; Segal, D.; Adonyeva, N.V.; Rauschenbach, I.Y. Stress response in a juvenile hormone-deficient Drosophila melanogaster mutant apterous56f. Insect Mol. Biol. 2003, 12, 353–363. [Google Scholar] [CrossRef]
- Horiuchi, M.; Endo, J.; Handa, Y.; Nose, H. Barometric pressure change and heart rate response during sleeping at ~ 3000 m altitude. Int. J. Biometeorol. 2018, 62, 909–912. [Google Scholar] [CrossRef]
- Haddad, G.G.; Sun, Y.; Wyman, R.J.; Xu, T. Genetic basis of tolerance to O2 deprivation in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1997, 94, 10809–10812. [Google Scholar] [CrossRef] [Green Version]
- Malacrida, S.; Giannella, A.; Ceolotto, G.; Reggiani, C.; Vezzoli, A.; Mrakic-Sposta, S.; Moretti, S.; Turner, R.; Falla, M.; Brugger, H.; et al. Transcription factors regulation in human peripheral white blood cells during hypobaric hypoxia exposure: An in-vivo experimental study. Sci. Rep. 2019, 9, 9901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrakic-Sposta, S.; Gussoni, M.; Dellanoce, C.; Marzorati, M.; Montorsi, M.; Rasica, L.; Pratali, L.; D’Angelo, G.; Martinelli, M.; Bastiani, L.; et al. Effects of acute and sub-acute hypobaric hypoxia on oxidative stress: A field study in the Alps. Eur. J. Appl. Physiol. 2021, 121, 297–306. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adonyeva, N.V.; Menshanov, P.N.; Gruntenko, N. A Link between Atmospheric Pressure and Fertility of Drosophila Laboratory Strains. Insects 2021, 12, 947. https://doi.org/10.3390/insects12100947
Adonyeva NV, Menshanov PN, Gruntenko N. A Link between Atmospheric Pressure and Fertility of Drosophila Laboratory Strains. Insects. 2021; 12(10):947. https://doi.org/10.3390/insects12100947
Chicago/Turabian StyleAdonyeva, Natalya V., Petr N. Menshanov, and Nataly Gruntenko. 2021. "A Link between Atmospheric Pressure and Fertility of Drosophila Laboratory Strains" Insects 12, no. 10: 947. https://doi.org/10.3390/insects12100947