
Spatial Distribution of Aedes aegypti Oviposition Temporal Patterns and Their Relationship with Environment and Dengue Incidence

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
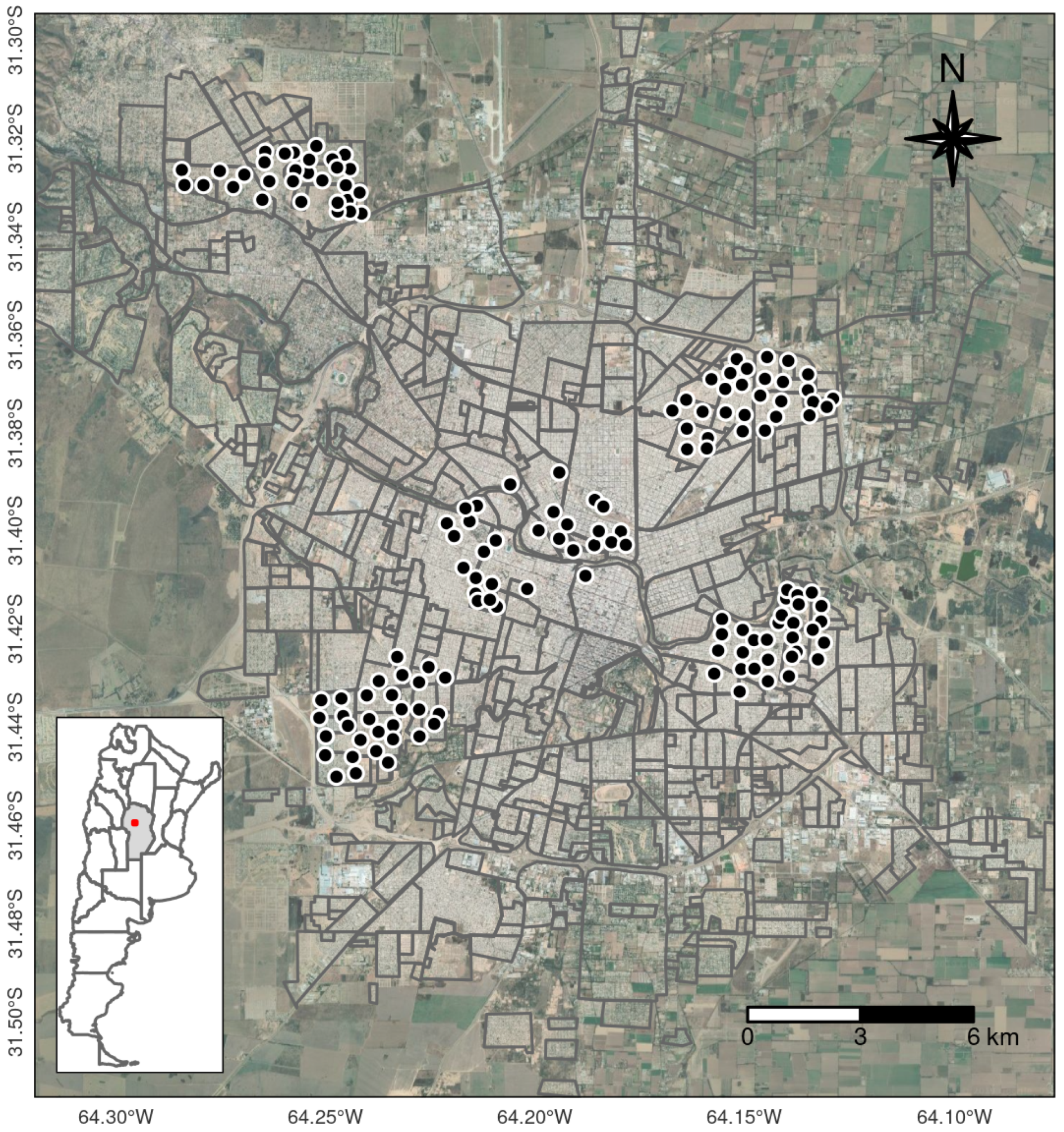
2.1. Study Area
2.2. Mosquito Data
2.3. Remote Sensing Data
2.4. Dengue Data
2.5. Data Analyses
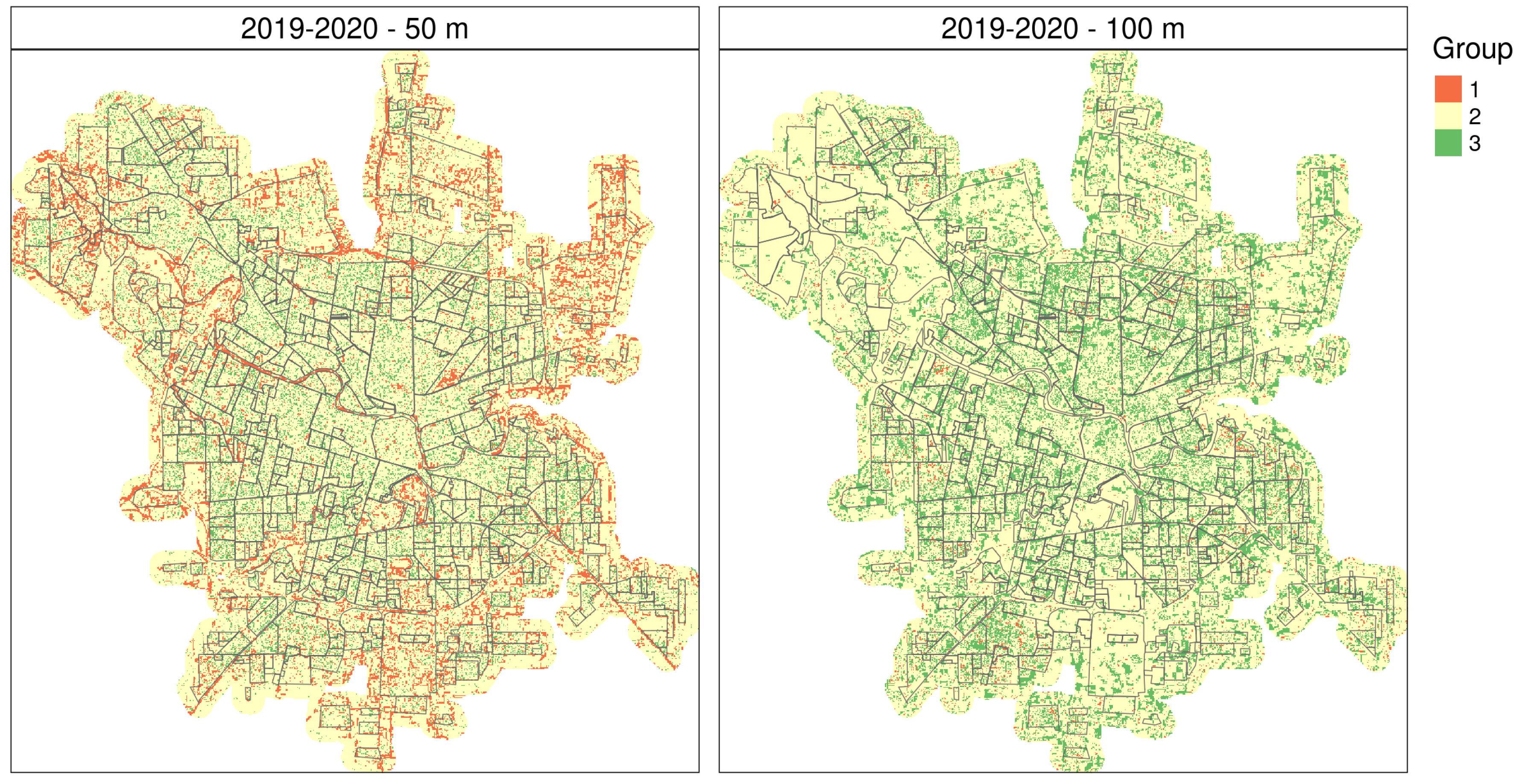
2.5.1. Time Series Clustering
2.5.2. Association with Environmental Features
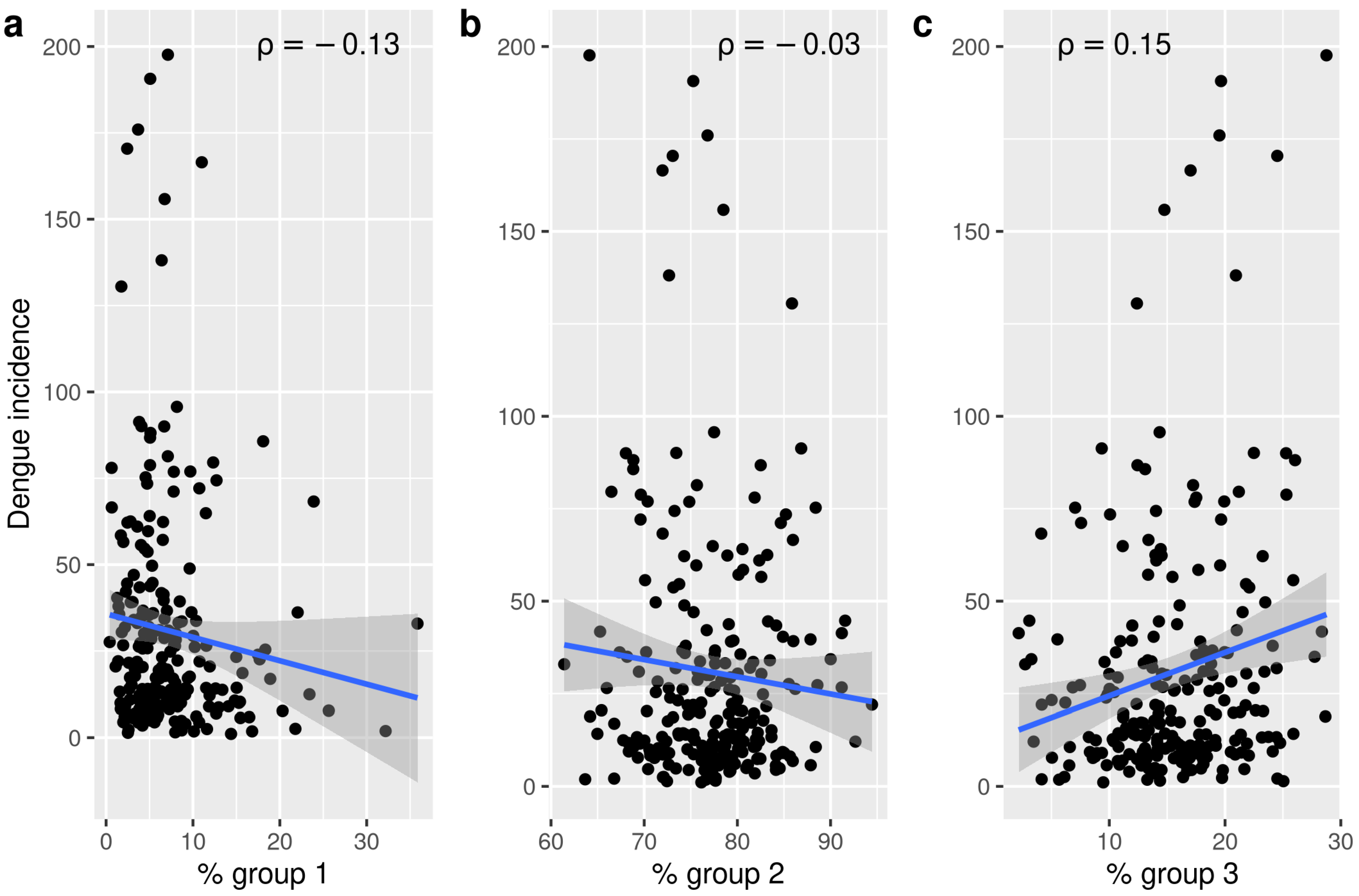
2.6. Oviposition Temporal Patterns and Dengue
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vezzani, D.; Carbajo, A.E. Aedes aegypti, Aedes albopictus, and dengue in Argentina: Current knowledge and future directions. Memórias do Instituto Oswaldo Cruz 2008, 103, 66–74. [Google Scholar] [CrossRef]
- Liu-Helmersson, J.; Rocklov, J.; Sewe, M.; Brannstrom, A. Climate change may enable Aedes aegypti infestation in major European cities by 2100. Environ. Res. 2019, 172, 693–699. [Google Scholar] [CrossRef]
- Rubio, A.; Cardo, M.V.; Vezzani, D.; Carbajo, A.E. Aedes aegypti spreading in South America: New coldest and southernmost records. Memórias do Instituto Oswaldo Cruz 2020, 115, e190496. [Google Scholar] [CrossRef]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A Review. Memórias do Instituto Oswaldo Cruz 2013, 108, 11–17. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Chase, C.; Vasquez, C.; Carvajal, A.; Medina, J.; Petrie, W.D.; Beier, J.C. Urbanization creates diverse aquatic habitats for immature mosquitoes in urban areas. Sci. Rep. 2019, 9, 15335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the Global Burden of Disease Study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Seijo, A.; Romer, Y.; Espinosa, M.; Monroig, J.; Giamperetti, S.; Ameri, D.; Antonelli, L.G. Outbreak of indigenous dengue in the Buenos Aires Metropolitan Area. Experience of the F. J. Muñíz Hospital. Medicina 2009, 69, 593–600. [Google Scholar] [PubMed]
- Estallo, E.L.; Carbajo, A.E.; Grech, M.G.; Frías-Céspedes, M.; López, L.; Lanfri, M.A.; Ludueña-Almeida, F.F.; Almirón, W.R. Spatio-temporal dynamics of dengue 2009 outbreak in Córdoba City, Argentina. Acta Trop. 2014, 136, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Rotela, C.; Lopez, L.; Céspedes, M.F.; Barbas, G.; Lighezzolo, A.; Porcasi, X.; Lanfri, M.A.; Scavuzzo, C.M.; Gorla, D.E. Analytical report of the 2016 dengue outbreak in Córdoba city, Argentina. Geospat. Health 2017. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Tinunin, D.T.; Benitez, E.M.; Ludueña-Almeida, F.F.; Romero, M.; Stewart-Ibarra, A.M.; Estallo, E.L. Arbovirus emergence in the temperate city of Córdoba, Argentina, 2009–2018. Sci. Data 2019, 6, 276. [Google Scholar] [CrossRef] [Green Version]
- Ministerio de Salud de la Nación. Boletín Integrado de Vigilancia; Semana 32; Ministerio de Salud de la Nación: de Julio, Argentina, 2020. [Google Scholar]
- Bowman, L.R.; Donegan, S.; McCall, P.J. Is Dengue Vector Control Deficient in Effectiveness or Evidence? Systematic Review and Meta-analysis. PLoS Neglected Trop. Dis. 2016, 10, e0004551. [Google Scholar] [CrossRef] [Green Version]
- Getis, A.; Morrison, A.C.; Gray, K.; Scott, T.W. Characteristics of the spatial pattern of the Dengue vector, Aedes aegypti, in Iquitos, Perú. Am. J. Trop. Med. Hyg. 2003, 69, 494–505. [Google Scholar] [CrossRef]
- Sallam, M.F.; Fizer, C.; Pilant, A.N.; Whung, P.Y. Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping. Int. J. Environ. Res. Public Health 2017, 14, 1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, R.; Li, Z.; Catry, T.; Roux, E.; Mangeas, M.; Handschumacher, P.; Gaudart, J.; Tran, A.; Demagistri, L.; Faure, J.F.; et al. A Mapping Review on Urban Landscape Factors of Dengue Retrieved from Earth Observation Data, GIS Techniques, and Survey Questionnaires. Remote Sens. 2020, 12, 932. [Google Scholar] [CrossRef] [Green Version]
- Porcasi, X.; Rotela, C.H.; Introini, M.V.; Frutos, N.; Lanfri, S.; Peralta, G.; Elia, E.A.D.; Lanfri, M.A.; Scavuzzo, C.M. An operative dengue risk stratification system in Argentina based on geospatial technology. Geospat. Health 2012, 6, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Estallo, E.L.; Sangermano, F.; Grech, M.; Ludueña-Almeida, F.; Frías-Cespedes, M.; Ainete, M.; Almirón, W.; Livdahl, T. Modelling the distribution of the vector Aedes aegypti in a central Argentine city: Modelling Aedes aegypti distribution. Med. Vet. Entomol. 2018, 32, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez, E.M.; Ludueña-Almeida, F.; Frías-Céspedes, M.; Almirón, W.R.; Estallo, E.L. Could land cover influence Aedes aegypti mosquito populations? Med. Vet. Entomol. 2020, 34, 138–144. [Google Scholar] [CrossRef]
- Lorenz, C.; Castro, M.C.; Trindade, P.M.P.; Nogueira, M.L.; de Oliveira Lage, M.; Quintanilha, J.A.; Parra, M.C.; Dibo, M.R.; Fávaro, E.A.; Guirado, M.M.; et al. Predicting Aedes aegypti infestation using landscape and thermal features. Sci. Rep. 2020, 10, 21688. [Google Scholar] [CrossRef]
- Andreo, V.; Cuervo, P.F.; Porcasi, X.; Lopez, L.; Guzman, C.; Scavuzzo, C.M. Towards a workflow for operational mapping of Aedes aegypti at urban scale based on remote sensing. Remote Sens. Appl. Soc. Environ. 2021, 23, 100554. [Google Scholar] [CrossRef]
- Aguirre, E.; Andreo, V.; Porcasi, X.; Lopez, L.; Guzman, C.; González, P.; Scavuzzo, C.M. Implementation of a proactive system to monitor Aedes aegypti populations using open access historical and forecasted meteorological data. Ecol. Inform. 2021, 64, 101351. [Google Scholar] [CrossRef]
- Estallo, E.L.; Benitez, E.M.; Lanfri, M.A.; Scavuzzo, C.M.; Almirón, W.R. MODIS Environmental Data to Assess Chikungunya, Dengue, and Zika Diseases Through Aedes (Stegomia) aegypti Oviposition Activity Estimation. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 9, 5461–5466. [Google Scholar] [CrossRef]
- Scavuzzo, J.M.; Trucco, F.; Espinosa, M.; Tauro, C.B.; Abril, M.; Scavuzzo, C.M.; Frery, A.C. Modeling Dengue vector population using remotely sensed data and machine learning. Acta Trop. 2018, 185, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudele, O.; Frery, A.C.; Zanandrez, L.F.R.; Eiras, A.E.; Gamba, P. Dengue Vector Population Forecasting Using Multisource Earth Observation Products and Recurrent Neural Networks. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 4390–4404. [Google Scholar] [CrossRef]
- Andreo, V.; Izquierdo-Verdiguier, E.; Zurita-Milla, R.; Rosa, R.; Rizzoli, A.; Papa, A. Identifying Favorable Spatio-Temporal Conditions for West Nile Virus Outbreaks by Co-Clustering of Modis LST Indices Time Series. In Proceedings of the IGARSS 2018—2018 IEEE International Geoscience and Remote Sensing Symposium, Valencia, Spain, 22–27 July 2018; pp. 4670–4673. [Google Scholar] [CrossRef]
- Albrieu-Llinás, G.; Espinosa, M.O.; Quaglia, A.; Abril, M.; Scavuzzo, C.M. Urban environmental clustering to assess the spatial dynamics of Aedes aegypti breeding sites. Geospat. Health 2018, 13, 135–142. [Google Scholar] [CrossRef]
- Aghabozorgi, S.; Seyed Shirkhorshidi, A.; Ying Wah, T. Time-series clustering—A decade review. Inf. Syst. 2015, 53, 16–38. [Google Scholar] [CrossRef]
- Franses, P.H.; Wiemann, T. Intertemporal Similarity of Economic Time Series: An Application of Dynamic Time Warping. Comput. Econ. 2020, 56, 59–75. [Google Scholar] [CrossRef]
- Belgiu, M.; Csillik, O. Sentinel-2 cropland mapping using pixel-based and object-based time-weighted dynamic time warping analysis. Remote Sens. Environ. 2017, 204, 509–523. [Google Scholar] [CrossRef]
- Naranjo-Fernández, N.; Guardiola-Albert, C.; Aguilera, H.; Serrano-Hidalgo, C.; Montero-González, E. Clustering Groundwater Level Time Series of the Exploited Almonte-Marismas Aquifer in Southwest Spain. Water 2020, 12, 1063. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hepner, G.F. The Dynamic-Time-Warping-based k-means++ clustering and its application in phenoregion delineation. Int. J. Remote Sens. 2017, 38, 1720–1736. [Google Scholar] [CrossRef]
- Andreo, V.; Porcasi, X.; Rodriguez, C.; Lopez, L.; Guzman, C.; Scavuzzo, C.M. Time Series Clustering Applied to Eco-Epidemiology: The case of Aedes aegypti in Córdoba, Argentina. In Proceedings of the 2019 XVIII Workshop on Information Processing and Control (RPIC), Bahía Blanca, Argentina, 18–20 September 2019; pp. 93–98. [Google Scholar] [CrossRef]
- INDEC (Instituto Nacional de Estadística y Censos). National Population, Homes and Houses Census; Censo Nacional de Población, Hogares y Viviendas, 2010. Available online: https://redatam.indec.gob.ar/argbin/RpWebEngine.exe/PortalAction (accessed on 10 March 2021).
- Espinosa, M.; Alvarez Di Fino, E.M.; Abril, M.; Lanfri, M.; Periago, M.V.; Scavuzzo, C.M. Operational satellite-based temporal modelling of Aedes population in Argentina. Geospat. Health 2018, 13, 247–258. [Google Scholar] [CrossRef] [Green Version]
- GRASS Development Team. Geographic Resources Analysis Support System (GRASS GIS) Software, Version 7.8; Open Source Geospatial Foundation, 2019. Available online: https://grass.osgeo.org/download (accessed on 23 March 2021).
- Arbelaitz, O.; Gurrutxaga, I.; Muguerza, J.; Pérez, J.M.; Perona, I. An extensive comparative study of cluster validity indices. Pattern Recognit. 2013, 46, 243–256. [Google Scholar] [CrossRef]
- Sarda-Espinosa, A. Dtwclust: Time Series Clustering Along with Optimizations for the Dynamic Time Warping Distance, R Package Version 5.5.6; 2019. Available online: https://cran.r-project.org/package=dtwclust (accessed on 21 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Hornik, K. Clue: Cluster Ensembles, R Package Version 0.3-59; 2021. Available online: https://cran.r-project.org/package=clue (accessed on 21 May 2021).
- Kuhn, M. Caret: Classification and Regression Training, R Package Version 6.0-88; 2021. Available online: https://cran.r-project.org/package=caret (accessed on 23 June 2021).
- Micieli, M.V.; Campos, R.E. Oviposition activity and seasonal pattern of a population of Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) in subtropical Argentina. Memórias do Instituto Oswaldo Cruz 2003, 98, 659–663. [Google Scholar] [CrossRef]
- Estallo, E.L.; Ludueña-Almeida, F.F.; Visintin, A.M.; Scavuzzo, C.M.; Introini, M.V.; Zaidenberg, M.; Almirón, W.R. Prevention of Dengue Outbreaks Through Aedes aegypti Oviposition Activity Forecasting Method. Vector-Borne Zoonotic Dis. 2011, 11, 543–549. [Google Scholar] [CrossRef]
- German, A.; Espinosa, M.O.; Abril, M.; Scavuzzo, C.M. Exploring satellite based temporal forecast modelling of Aedes aegypti oviposition from an operational perspective. Remote Sens. Appl. Soc. Environ. 2018, 11, 231–240. [Google Scholar] [CrossRef]
- Benitez, E.M.; Estallo, E.L.; Grech, M.G.; Frías-Céspedes, M.; Almirón, W.R.; Robert, M.A.; Ludueña-Almeida, F.F. Understanding the role of temporal variation of environmental variables in predicting Aedes aegypti oviposition activity in a temperate region of Argentina. Acta Trop. 2021, 216, 105744. [Google Scholar] [CrossRef] [PubMed]
- Manica, M.; Rosà, R.; Torre, A.D.; Caputo, B. From eggs to bites: Do ovitrap data provide reliable estimates of Aedes albopictus biting females? PeerJ 2017, 5, e2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landau, K.I.; Leeuwen, W.J.D.V. Fine scale spatial urban land cover factors associated with adult mosquito abundance and risk in Tucson, Arizona. J. Vector Ecol. 2012, 37, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, M.O.; Polop, F.; Rotela, C.H.; Abril, M.; Scavuzzo, C.M. Spatial pattern evolution of Aedes aegypti breeding sites in an Argentinean city without a dengue vector control programme. Geospat. Health 2016, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Whiteman, A.; Li, A.; Rapp, T.; Delmelle, E.; Chen, G.; Brown, C.L.; Robinson, P.; Coffman, M.J.; Janies, D.; et al. An operational machine learning approach to predict mosquito abundance based on socioeconomic and landscape patterns. Landsc. Ecol. 2019, 34, 1295–1311. [Google Scholar] [CrossRef]
- Cromwell, E.A.; Stoddard, S.T.; Barker, C.M.; Rie, A.V.; Messer, W.B.; Meshnick, S.R.; Morrison, A.C.; Scott, T.W. The relationship between entomological indicators of Aedes aegypti abundance and dengue virus infection. PLoS Neglected Trop. Dis. 2017, 11, e0005429. [Google Scholar] [CrossRef]
- Lee, K.Y.; Chung, N.; Hwang, S. Application of an artificial neural network (ANN) model for predicting mosquito abundances in urban areas. Ecol. Inform. 2016, 36, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, M.; Weinberg, D.; Rotela, C.H.; Polop, F.; Abril, M.; Scavuzzo, C.M. Temporal Dynamics and Spatial Patterns of Aedes aegypti Breeding Sites, in the Context of a Dengue Control Program in Tartagal (Salta Province, Argentina). PLoS Neglected Trop. Dis. 2016, 10, e0004621. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance | Centroid | Pre-Processing | Configurations |
|---|---|---|---|
| DTW | DBA | Normalization | 1600 |
| DTW_LB | DBA | Normalization | 1600 |
| DTW | PAM | Normalization | 1600 |
| DTW_LB | PAM | Normalization | 1600 |
| SBD | PAM | Normalization | 160 |
| SBD | Shape extraction | Normalization | 160 |
| Series | Rep | k | n | Dist | Cent | Dist Wind Size | Norm Dist | Cent Wind Size | Norm Cent | Znorm Cent | Sil | D | COP | DB | DB* | Votes |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Full series | 10 | 3 | 89,44,10 | dtw_basic | dba | 4 | L2 | 4 | L2 | 0.095 | 0.311 | 2.026 | 0.581 | 0.581 | 1 | |
| 10 | 3 | 89,44,10 | dtw_lb | dba | 4 | L2 | 4 | L2 | 0.158 | 0.076 | 1.697 | 0.413 | 0.404 | 0 | ||
| 2 | 3 | 16,26,101 | dtw_basic | pam | 5 | L2 | 0.100 | 0.315 | 1.867 | 0.739 | 0.702 | 3 | ||||
| 7 | 3 | 110,9,24 | dtw_lb | pam | 3 | L2 | 0.192 | 0.099 | 1.700 | 0.445 | 0.436 | 1 | ||||
| 7 | 3 | 6,111,26 | sbd | shape | TRUE | 0.181 | 0.123 | 1.700 | 0.575 | 0.562 | 0 | |||||
| 1 | 3 | 25,12,106 | sbd | pam | 0.098 | 0.203 | 2.023 | 0.537 | 0.489 | 0 | ||||||
| 2017–2018 | 10 | 4 | 30,71,14,28 | dtw_basic | dba | 4 | L2 | 4 | L2 | 0.058 | 0.233 | 1.889 | 0.615 | 0.612 | 1 | |
| 1 | 3 | 54,37,52 | dtw_lb | dba | 2 | L2 | 2 | L2 | 0.119 | 0.098 | 1.602 | 0.454 | 0.454 | 1 | ||
| 7 | 3 | 70,30,43 | dtw_basic | pam | 5 | L1 | 0.049 | 0.234 | 1.551 | 0.711 | 0.688 | 2 | ||||
| 5 | 5 | 44,49,29,10,11 | dtw_lb | pam | 1 | L2 | 0.106 | 0.167 | 1.423 | 0.534 | 0.517 | 0 | ||||
| 8 | 3 | 48,30,65 | sbd | shape | TRUE | 0.097 | 0.191 | 2.422 | 0.665 | 0.627 | 1 | |||||
| 10 | 3 | 41,44,58 | sbd | pam | 0.076 | 0.112 | 1.958 | 0.507 | 0.492 | 0 | ||||||
| 2018–2019 | 8 | 3 | 10,89,44 | dtw_basic | dba | 4 | L2 | 4 | L2 | 0.140 | 0.301 | 1.985 | 0.692 | 0.663 | 1 | |
| 3 | 4 | 37,42,13,51 | dtw_lb | dba | 3 | L2 | 3 | L2 | 0.132 | 0.053 | 1.510 | 0.429 | 0.423 | 0 | ||
| 10 | 4 | 24,21,77,21 | dtw_basic | pam | 5 | L2 | 0.084 | 0.295 | 1.883 | 0.703 | 0.679 | 1 | ||||
| 7 | 3 | 25,39,79 | dtw_lb | pam | 5 | L1 | 0.254 | 0.014 | 0.586 | 0.120 | 0.117 | 1 | ||||
| 6 | 3 | 10,88,45 | sbd | shape | TRUE | 0.111 | 0.136 | 2.667 | 0.702 | 0.654 | 2 | |||||
| 4 | 3 | 37,37,69 | sbd | pam | 0.075 | 0.082 | 2.307 | 0.639 | 0.627 | 0 |
| Series | Cluster | n | MSD | MED | Duration (Days) | Max Eggs | Mean (Max Eggs) | Date of Max (Eggs) |
|---|---|---|---|---|---|---|---|---|
| 2017–2018 | 1 | 70 | 20 November 2017 | 7 May 2018 | 168 | 205 | 73 | 1 February 2018 |
| 2 | 30 | 4 December 2017 | 7 May 2018 | 154 | 155 | 55 | 15 January 2018 | |
| 3 | 43 | 27 November 2017 | 30 April 2018 | 154 | 196 | 74 | 29 January 2018 | |
| 2018–2019 | 1 | 10 | 25 October 2018 | 29 April 2019 | 186 | 274 | 127 | 11 February 2019 |
| 2 | 88 | 5 November 2018 | 29 April 2019 | 175 | 594 | 190 | 21 January 2019 | |
| 3 | 45 | 29 October 2018 | 29 April 2019 | 182 | 615 | 206 | 4 February 2019 |
| 2017–2018 | 2018–2019 | ||
|---|---|---|---|
| Train | 50 m | 0.733 (0.083) | 0.822 (0.051) |
| 100 m | 0.730 (0.080) | 0.778 (0.090) | |
| Test | 50 m | 0.381 | 0.524 |
| 100 m | 0.476 | 0.500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreo, V.; Porcasi, X.; Guzman, C.; Lopez, L.; Scavuzzo, C.M. Spatial Distribution of Aedes aegypti Oviposition Temporal Patterns and Their Relationship with Environment and Dengue Incidence. Insects 2021, 12, 919. https://doi.org/10.3390/insects12100919
Andreo V, Porcasi X, Guzman C, Lopez L, Scavuzzo CM. Spatial Distribution of Aedes aegypti Oviposition Temporal Patterns and Their Relationship with Environment and Dengue Incidence. Insects. 2021; 12(10):919. https://doi.org/10.3390/insects12100919
Chicago/Turabian StyleAndreo, Verónica, Ximena Porcasi, Claudio Guzman, Laura Lopez, and Carlos M. Scavuzzo. 2021. "Spatial Distribution of Aedes aegypti Oviposition Temporal Patterns and Their Relationship with Environment and Dengue Incidence" Insects 12, no. 10: 919. https://doi.org/10.3390/insects12100919