Biodiversity and Spatiotemporal Variation of Longhorn Beetles (Coleoptera: Cerambycidae) in Tropical Forest of Thailand


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
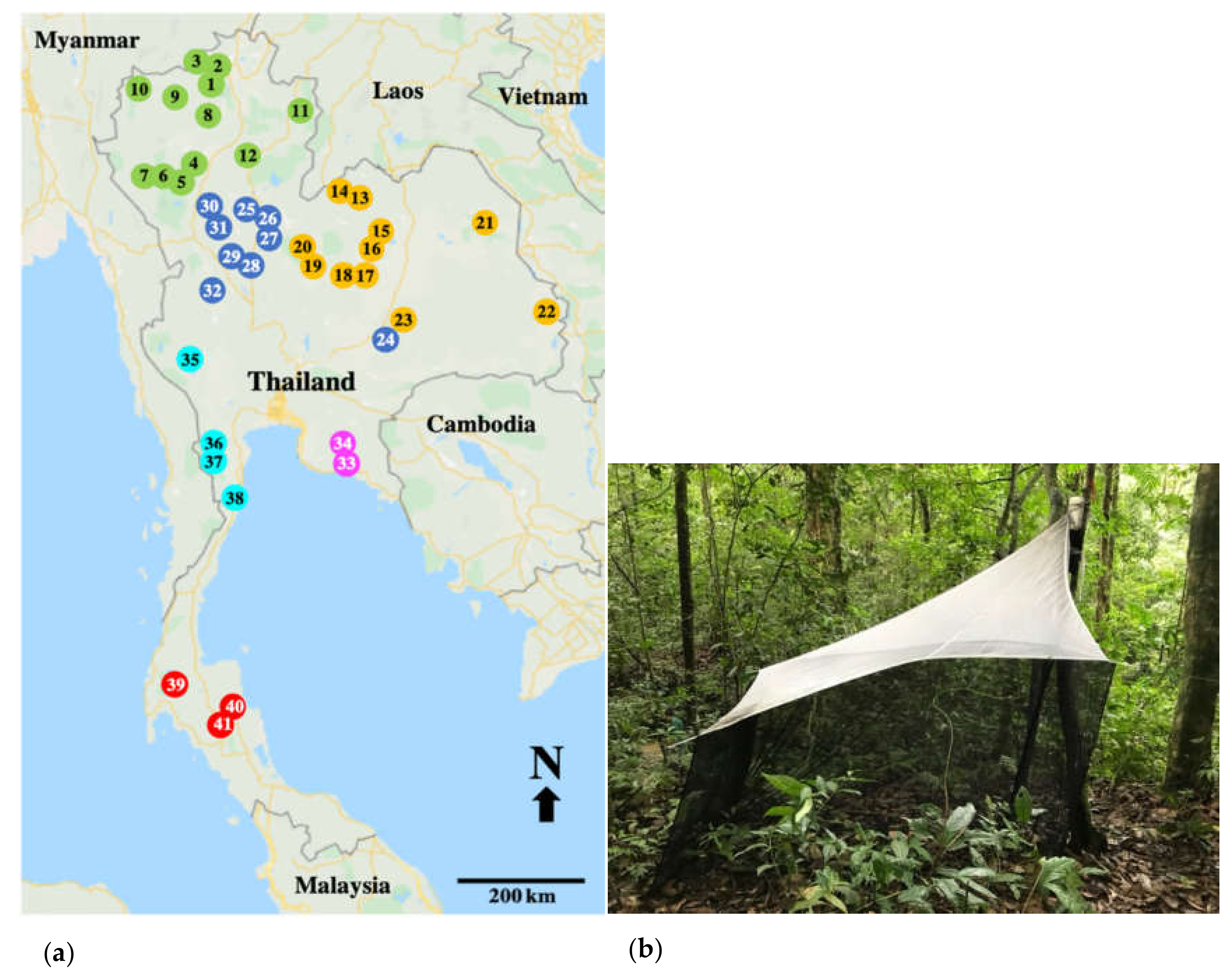
2.1. Study Area
2.2. Collection Methods
2.3. Species Identification
2.4. Statistical Analyses
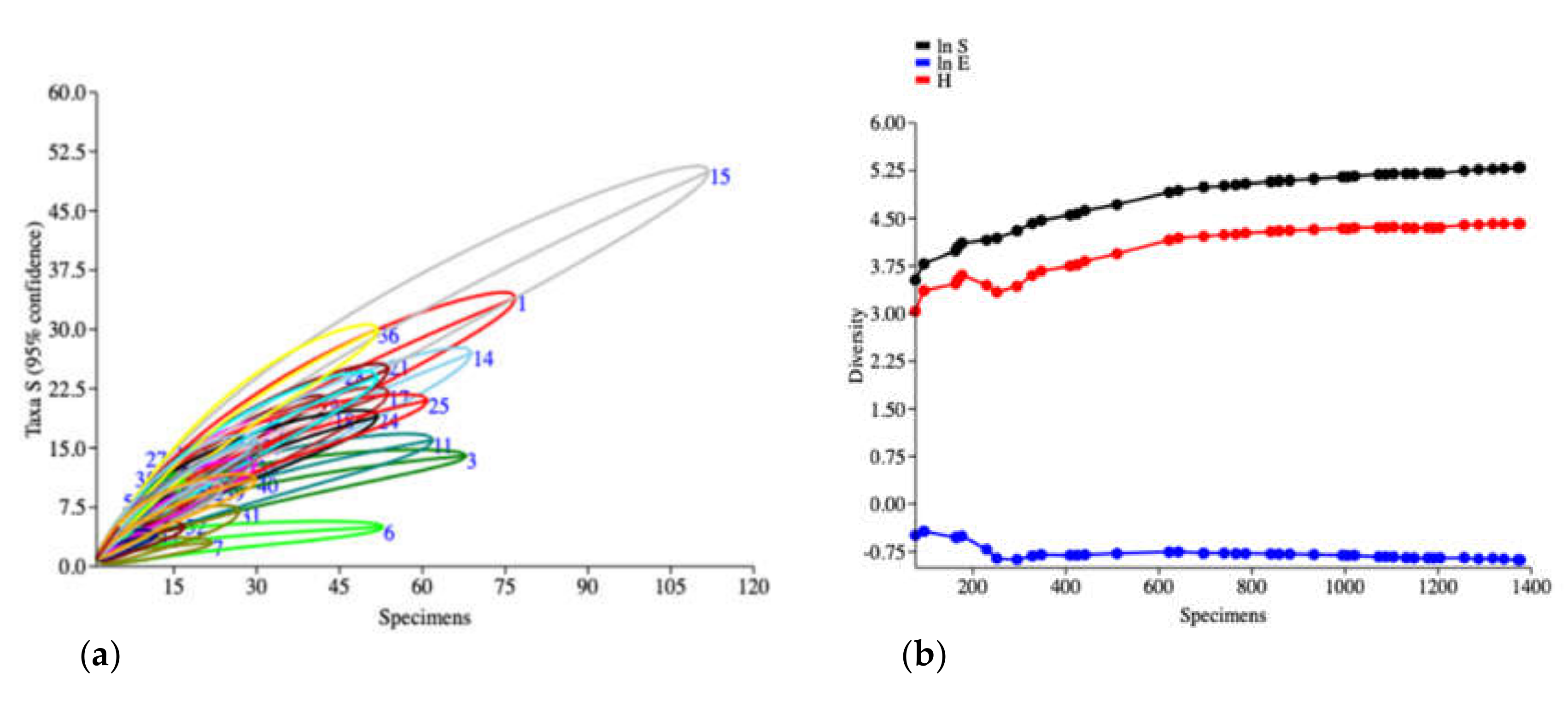
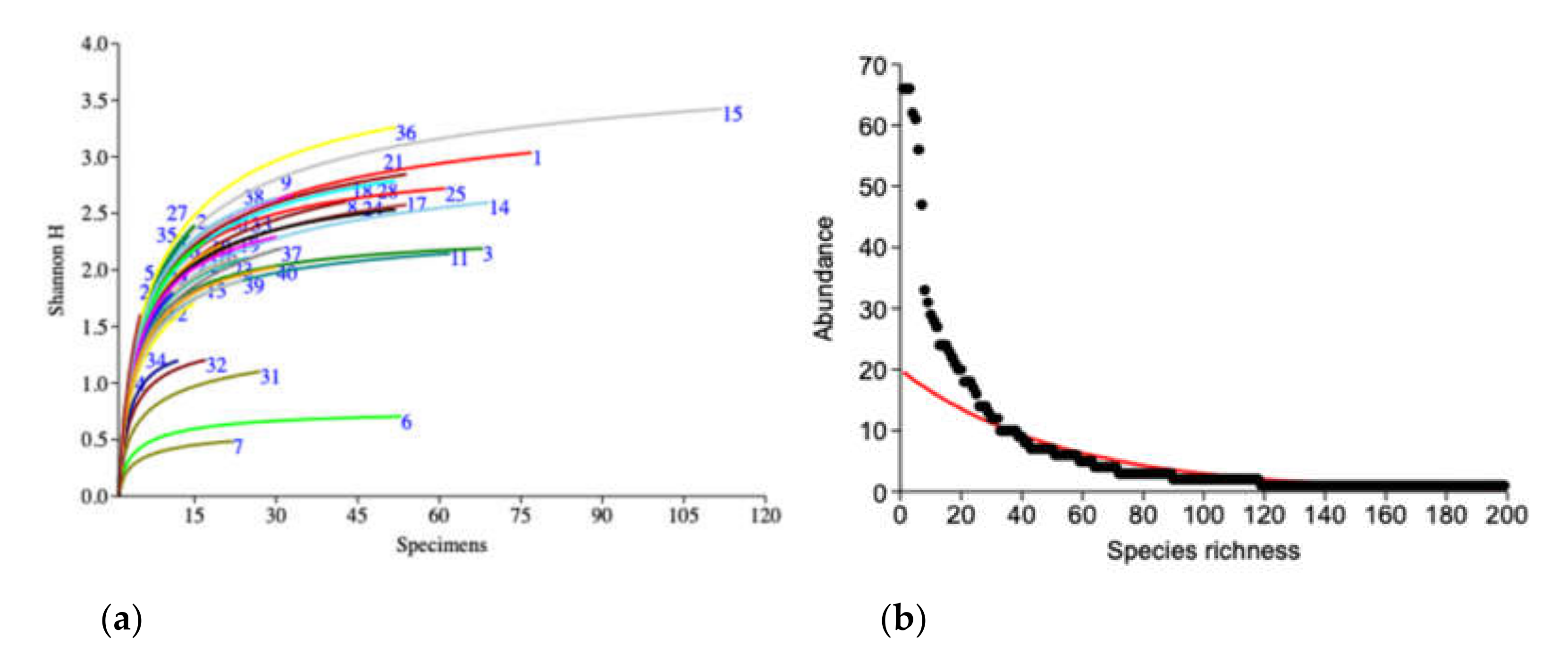
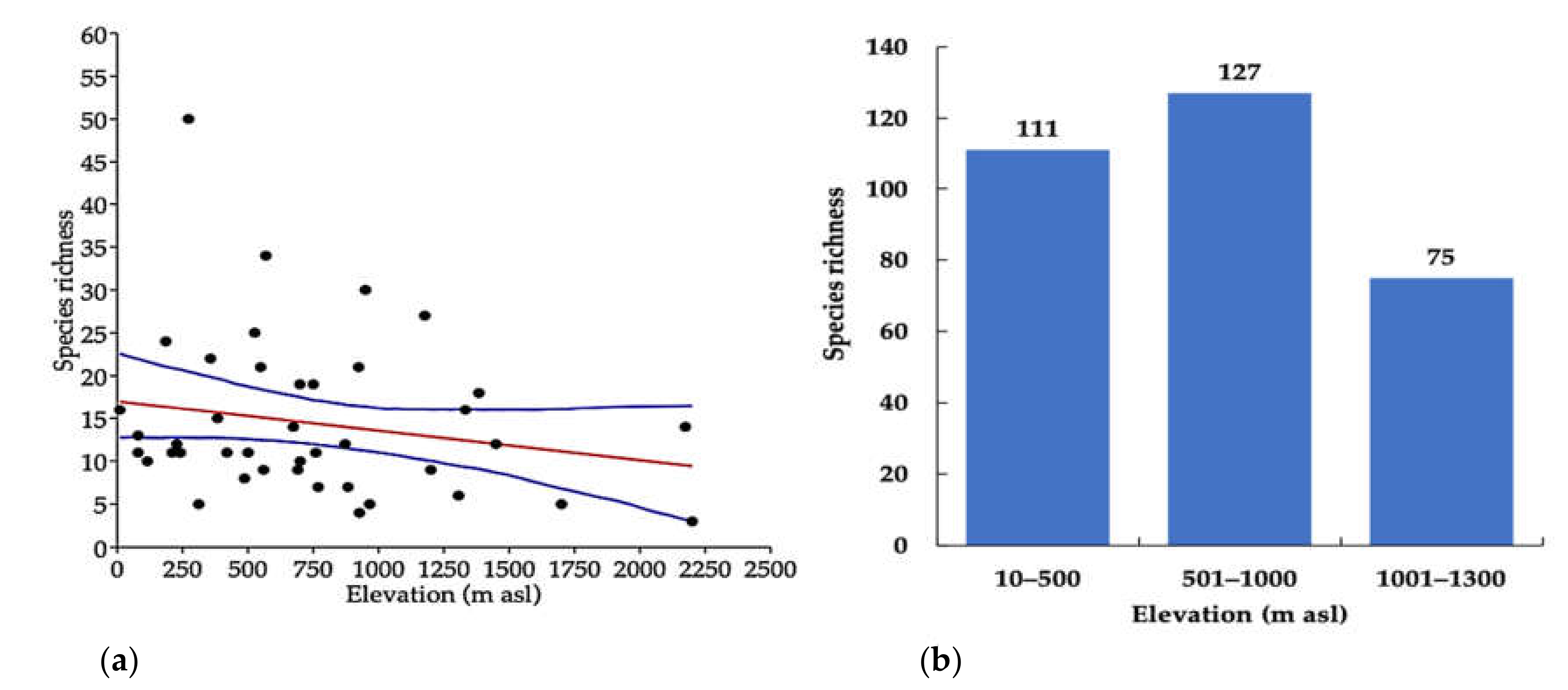
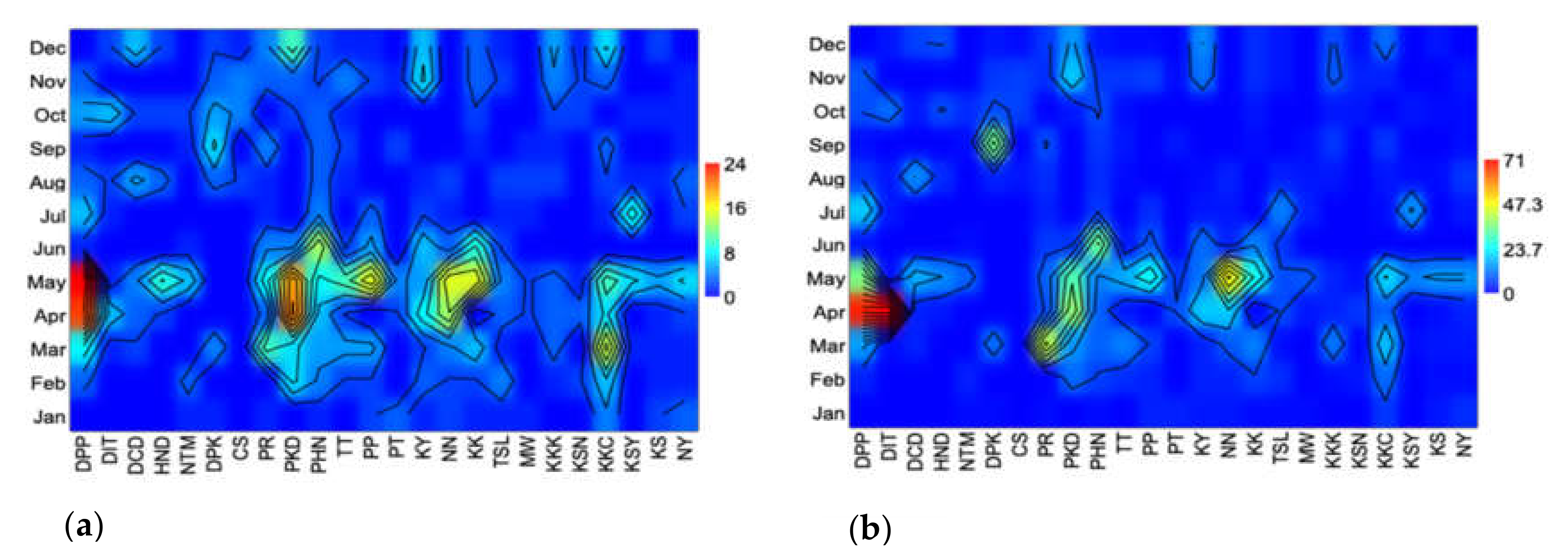
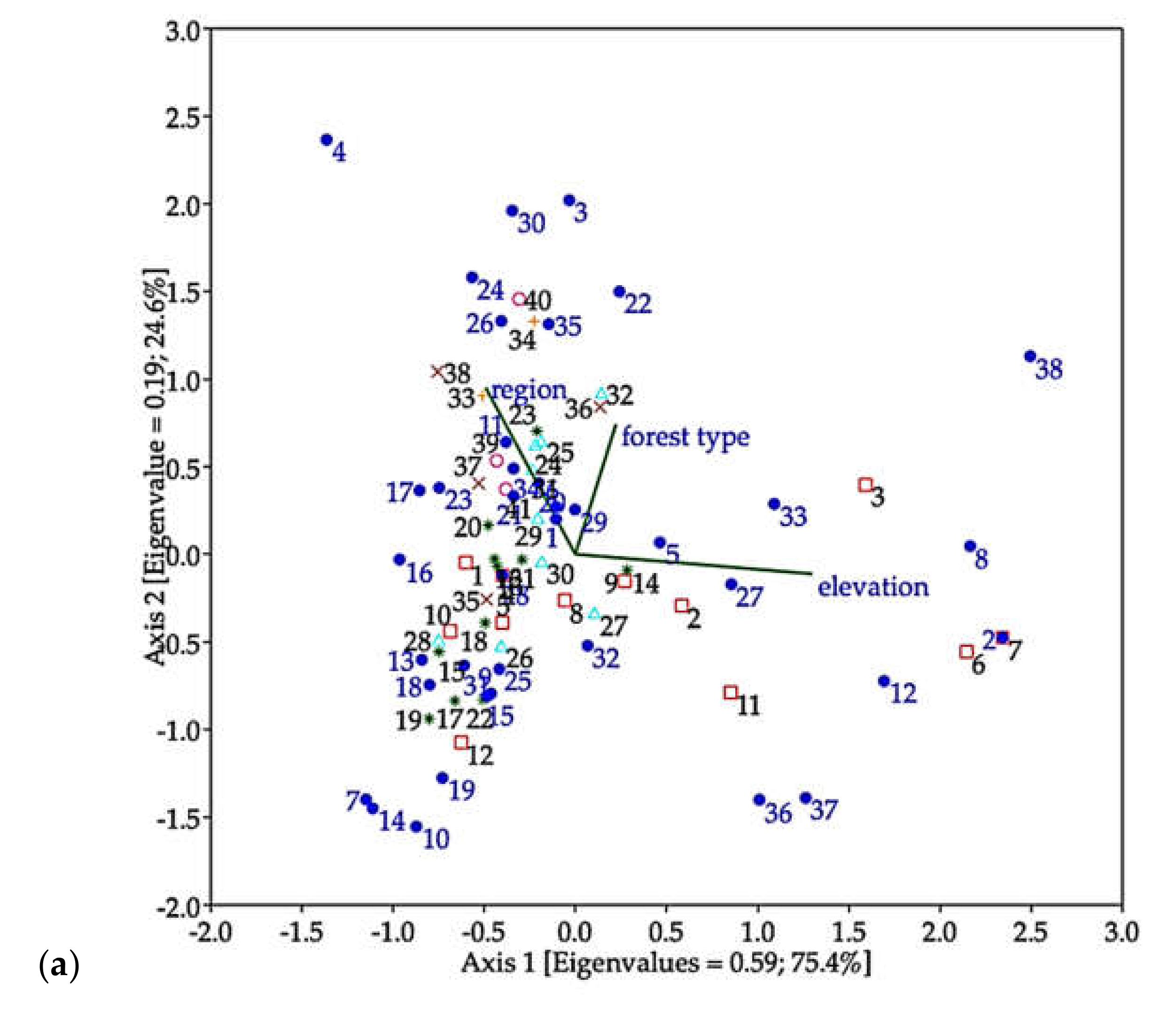
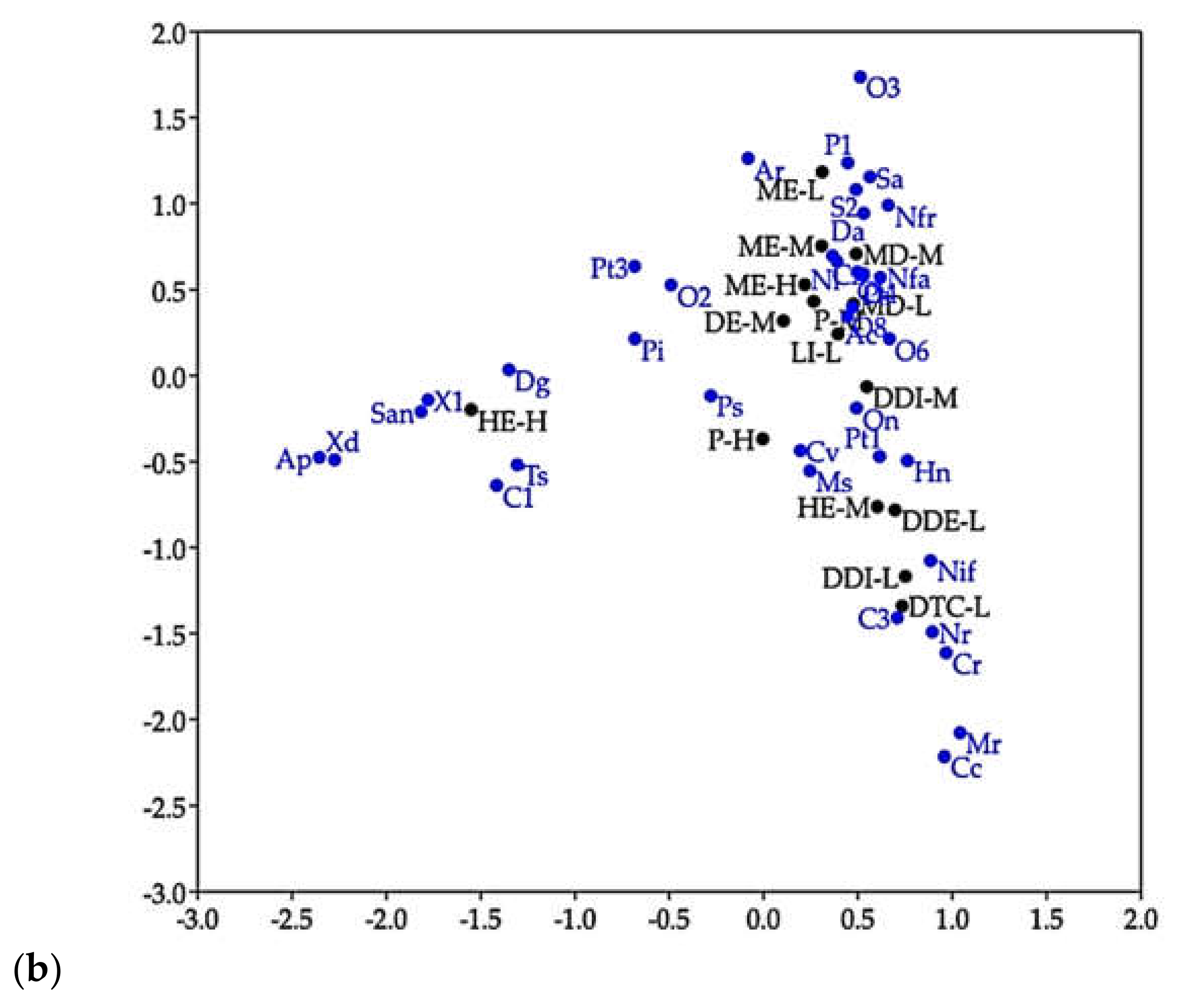
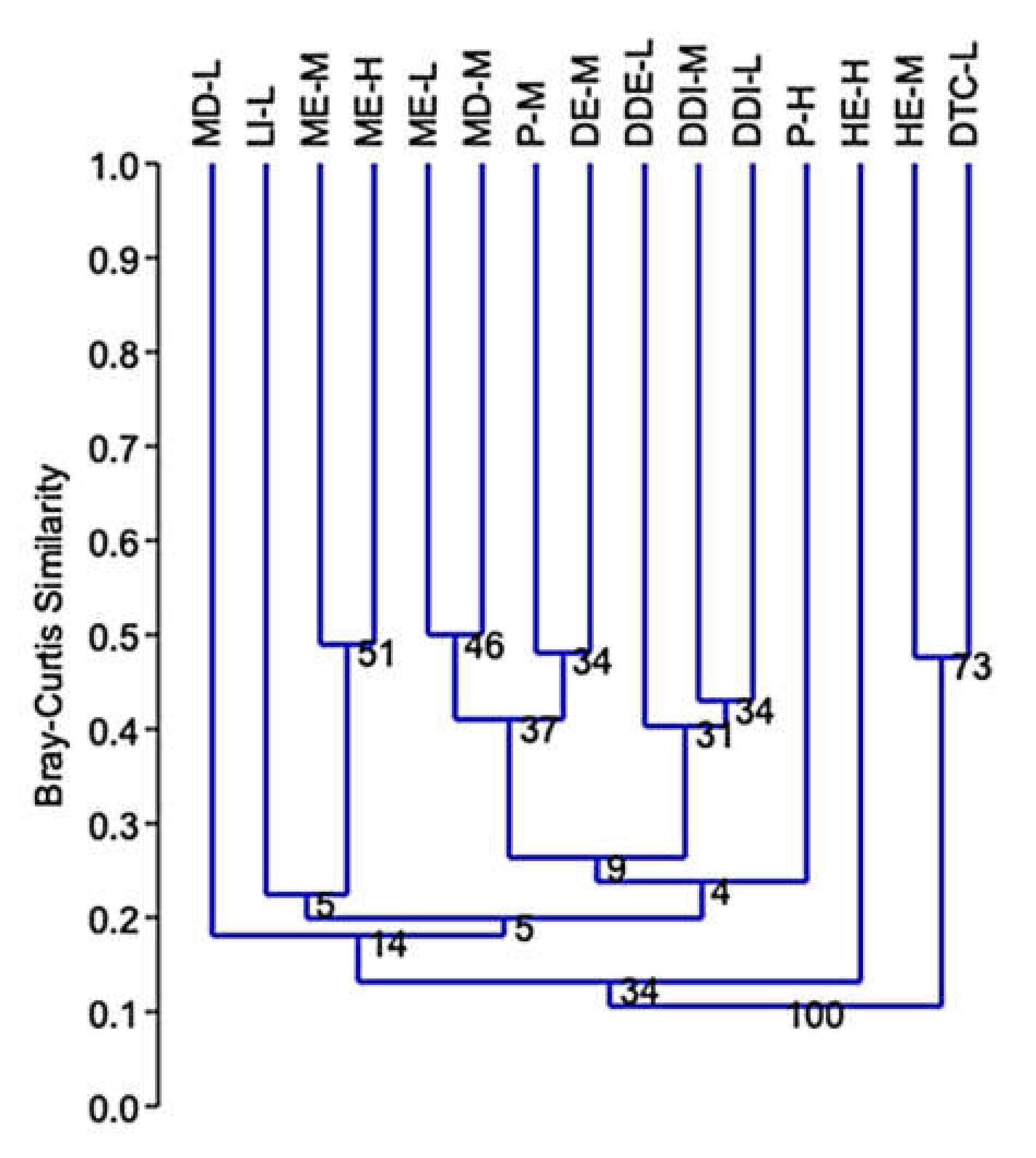
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monne’, M.L.; Momme’, M.A.; Wang, Q. General morphology, Classification, and Biology of Cerambycidae. In Cerambycidae of the World Biology and Pest Management; Wang, Q., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 1–70. [Google Scholar]
- Slipinski, A.; Escalona, H. Australian Longhorn Beetles (Coleoptera: Cerambycidae) Volume 1: Introduction and Subfamily Lamiinae; CSIRO Publishing: Collingwood, Australia, 2013; p. 504. [Google Scholar]
- Wang, J.; Dai, X.; Xu, X.; Zhang, Z.; Yu, D.; Storey, K.B.; Zhang, J. The complete mitochondrial genomes of five longicorn beetles (Coleoptera: Cerambycidae) and phylogenetic relationships within Cerambycidae. PeerJ 2019, 7, e7633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nearns, E.H.; Lord, N.P.; Lingafelter, S.W.; Santos-Silva, A.; Miller, K.B.; Zaspel, J.M. Longicorn ID Tool for Diagnosing Cerambycoid Families, Subfamilies, and Tribes. Available online: https://cerambycids.com/LongicornID/ (accessed on 19 August 2020).
- CABI. Current Year. Invasive Species Compendium. Wallingford, UK: CAB International. Available online: www.cabi.org/isc (accessed on 19 August 2020).
- Michael, T.S.; Turgeon, J.J.; de Groot, P.; Gasman, B. Asian longhorned beetle Anoplophora glabripennis (Motschulsky): Lessons learned and opportunities to improve the process of eradication and management. Am. Entomol. 2009, 55, 21–25. [Google Scholar]
- Monteys, V.S.; Tutusaus, G.T. A new alien invasive longhorn beetle, Xylotrechus chinensis (Cerambycidae), is infesting mulberries in Catalonia (Spain). Insects 2018, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loni, A.; Jucker, C.; Belokobylskij, S.; Lupi, D. First record of Rhoptrocentrus piceus Marshall (Hymenoptera, Braconidae, Doryctinae) as parasitoid of Psacothea hilaris hilaris (Pascoe) (Coleoptera, Cerambycidae). ZooKeys 2015, 482, 1–8. [Google Scholar]
- Gutowski, J.M. Pollination of the orchid dactylorhiza fuchsii by longhorn beetles in primeval forests of northeastern Poland. Biol. Conserv. 1990, 51, 287–297. [Google Scholar] [CrossRef]
- Martínez, R.D.; Basterra, L.A.; Acuña, L.; Balmori, J.A. Morphology and material composition of the mouthparts of Stromatium unicolor Olivier 1795 (Coleoptera: Cerambycidae) for bionic application. Forests 2020, 11, 715. [Google Scholar] [CrossRef]
- Noguera, F.A.; Zaragoza-Caballero, S.; Chemsak, J.A.; Rodriguez-Palafox, A.; Ramirez, E.; Gonzalez-Soriano, E.; Ayala, R. Diversity of the family Cerambycidae (Coleoptera) of the tropical dry forest of Mexico, I. Sierra de Huautla, Morelos. Ann. Entomol. Soc. Am. 2020, 95, 617–627. [Google Scholar] [CrossRef]
- Pavuk, D.M.; Wadsworth, A.M. Longhorned beetle (Coleoptera: Cerambycidae) diversity in a fragmented temperate forest landscape. F1000Research 2012, 1, 25. [Google Scholar] [CrossRef]
- Martínez-Hernández, J.G.; Corona-López, A.M.; Flores-Palacios, A.; Rös, M.; Toledo-Hernöndez, V.H. Seasonal diversity of Cerambycidae (Coleoptera) is more complex than thought: Evidence from a tropical dry forest of Mexico. PeerJ 2019, 7, e7866. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Flores, O.; Toledo-Hernández, V.H. Diversity of the longhorned beetles (Coleoptera: Cerambycidae) from Cuatro Ciénegas Basin, Coahuila, Mexico. Biodivers. Data J. 2020, 8, e54495. [Google Scholar] [CrossRef]
- Hiroshi, M.; Noerdjito, W.A.; Sugiarto. Longicorn beetles from Gunung Halimun National Park, West Java, Indonesia from 1997–2002 (Coleoptera, Disteniidae and Cerambycidae). Bull. FFPRI 2002, 1, 189–223. [Google Scholar]
- Meng, L.Z.; Martin, K.; Weigel, A.; Yang, X.D. Tree diversity mediates the distribution of longhorn beetles (Coleoptera: Cerambycidae) in a changing tropical landscape (Southern Yunnan, SW China). PLoS ONE 2013, 8, e75481. [Google Scholar] [CrossRef]
- Fahri, F.; Atmowidi, T.; Noerdjito, W.A. Diversity and abundance of cerambycid beetles in the four major land-use types found in Jambi province, Indonesia. HAYATI J. Biosci. 2016, 23, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.K.; Yamasako, J.; Kato, T.; Saito, M.; Ito, M. Fauna of Cerambycidae (Insecta: Coleoptera) in Komaba Campus of the University of Tokyo, a highly urbanised area in Japan. Biodivers. Data J. 2017, 5, e22296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutacharern, C.; Tubtim, N.; Dokmai, C. Checklists of Insects and Mites in Thailand; Department of National Parks, Wildlife and Plant Conservation, Ministry of natural Resources and Environment: Bangkok, Thailand, 2007; p. 319.
- Pase, H.A.; Coster, J.E. Some Cerambycidae (Coleoptera) from the Korat plateau region of Thailand. Can. Entomol. 1973, 105, 1059–1064. [Google Scholar] [CrossRef]
- Beaver, R.A. Longicorn beetles (Coleoptera: Cerambycidae) from northern Thailand. Nat. Hist. Bull. Siam Soc. 1975, 26, 51–60. [Google Scholar]
- Ponpinij, S. Taxonomy of the Longhorn Beetles (Coleoptera: Cerambycidae) in Northern of Thailand. Ph.D. Thesis, Kasetsart University, Bangkok, Thailand, 2011; p. 703. [Google Scholar]
- Thai Meteorological Department. Available online: https://tmd.go.th/info/info.php?FileID=23/ (accessed on 20 August 2020).
- Ponpinij, S.; Hormchan, P.; Rojanavongse, V. Checklist of new records of Subfamily Lamiinae (Coleoptera: Cerambycidae) in northern Thailand. Kasetsart J. (Nat. Sci.). 2011, 45, 841–855. [Google Scholar]
- Keyzer, R.D.; Niisato, T. An additional species of the genus Longipalpus (Coleoptera: Cerambycidae) from Thailand. Jpn. J. Entomol. 1989, 57, 333–336. [Google Scholar]
- Lingafelter, S.W.; Hoebeke, E.R. Revision of the Genus Anoplophora (Coleoptera: Cerambycidae); Entomological Society of Washington: Washington, DC, USA, 2002; p. 236. [Google Scholar]
- Lazarev, M.A.; Murzin, S.V. Two new species of Cleomenini (Coleoptera: Cerambycidae) from Myanmar and Thailand. Mun. Entomol. Zool. 2020, 15, 477–480. [Google Scholar]
- Holzschuh, C. Beschreibung von zehn neuen Bockkäferarten (Coleoptera: Cerambycidae) und einer neuen Gattung aus Südostasien. Z. Arb. Osterr. Entomol. 2013, 65, 5–21. [Google Scholar]
- Holzschuh, C. Die Gattung Exocentrus (Coleoptera: Cerambycidae: Lamiinae auf dem asiatischen Festland: Neue Synonymien und neue Taxa. Z. Arb. Osterr. Entomol. 2015, 67, 45–72. [Google Scholar]
- Holzschuh, C. Neue Clytini (Coleoptera: Cerambycidae) aus Laos und zur Synnonymie einiger arten. Z. Arb. Osterr. Entomol. 2016, 68, 103–127. [Google Scholar]
- Miroshnikov, A.I. The longicorn beetle tribe Cerambycini Latreille, 1802 (Coleoptera: Cerambycidae: Cerambycinae) in the fauna of Asia. 1. New or little-known taxa, mainly from Indochina and Borneo, with reviews of some genera. Cauc. Entomol. Bull. 2017, 13, 161–233. [Google Scholar] [CrossRef]
- Miroshnikov, A.I. The longicorn beetle tribe Cerambycini Latreille, 1802 (Coleoptera: Cerambycidae: Cerambycinae) in the fauna of Asia. 4. New or little-known taxa, mainly from Indochina and Borneo, with reviews or annotated checklists of species of some genera. Cauc. Entomol. Bull. 2018, 14, 197–246. [Google Scholar] [CrossRef]
- Srisuka, W.; Takaoka, H.; Otsuka, Y.; Fukuda, M.; Thongsahuan, S.; Taai, K.; Choochote, W.; Saeung, A. Biodiversity, seasonal abundance and distribution of black flies (Diptera: Simuliidae) in six different regions in Thailand. Parasit. Vectors 2017, 10, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron 2001, 4, 1–9. [Google Scholar]
- Skvarla, M.J.; Dowling, A.P.G. A comparison of trapping techniques (Coleoptera: Carabidae, Buprestidae, Cerambycidae, and Curculionoidea excluding Scolytinae). J. Insect Sci. 2017, 17, 1–28. [Google Scholar] [CrossRef]
- Burington, Z.L.; Niclan-Luna, D.J.; Pollet, M.; Stireman, J.O., III. Latitudinal patterns in tachinid parasitoid diversity (Diptera: Tachinidae): A review of the evidence. Insect Conserv. Diver. 2020, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Wilkie, K.T.R.; Mertl, A.L.; Traniello, J.F.A. Correction: Species Diversity and Distribution Patterns of the Ants of Amazonian Ecuador. PLoS ONE 2010, 5, 10. [Google Scholar]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Hauser, M.; le Cesne, M.; Surin, C.; Saokhod, R.; Nama, S.; Soulier-Perkins, A.; Daugeron, C.; et al. Spatiotemporal dynamics of insect diversity in tropical seasonal forests is linked to season and elevation, a case from northern Thailand. Raffles Bull. Zool. 2018, 66, 382–393. [Google Scholar]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Daugeron, C.; Srisuka, W. Composition and organization of highly specioes Empidoidea (Diptera) communities in tropical montane of northern Thailand. Zootaxa 2019, 4590, 1. [Google Scholar] [CrossRef] [PubMed]
- Erwin, T.L. Tropical forests: Their richness in Coleoptera and other arthropod species. Coleopts Bull. 1982, 36, 74–75. [Google Scholar]
- Stork, N.E. How many species of insects and other terrestrial arthropod there on Earth? Annu. Rev. Entomol. 2017, 63, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Gressitt, J.L.; Rondon, J.A.; Breuning, S.V. Cerambycid-beetles of Laos (Longicornes du Laos). Pac. Insects Monogr. 1970, 24, 651. [Google Scholar]
- Heffern, D.J. Catalog and Bibliography of Longhorned Beetles from Borneo (Coleoptera: Cerambycidae). Electronic Version. 2005. Available online: https://www.zin.ru/animalia/Coleoptera/pdf/borneo_catalog_electronic_version_2005-1.pdf (accessed on 18 October 2020).
- Kariyanna, B.; Mohan, M.; Gupta, R.; Vitali, F. The checklist of longhorn beetles (Coleoptera: Cerambycidae) from India. Zootaxa 2017, 4345, 317. [Google Scholar] [CrossRef]
- Peris-Felipo, F.J.; Falco-Gari, J.V.; Jimenez-Peydro, R. The diversity of Cerambycidae in the protected Mediterranean landscape of the Natural Park of Carrascal de La Font Roja, Spain. Bull. Insectology 2011, 64, 87–92. [Google Scholar]
- Peris-Felipo, F.J.; Jimenez-Peydro, R. Cerambycidae (Coleoptera) diversity and community structure in the Mediterranean forest of the Natural Park of Sierra Calderona (Spain). Frustula Entomol. 2010, 33, 180–191. [Google Scholar]
- Hanks, L.M.; Reagel, P.F.; Mitchell, R.F.; Wong, J.C.H.; Meier, L.R.; Silliman, C.A.; Graham, E.E.; Striman, B.L.; Robinson, K.P.; Mongold-Diers, J.A.; et al. Seasonal phenology of the cerambycid beetles of east-central Illinois. Ann. Entomol. Soc. Am. 2014, 107, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Handley, K.; Hough-Goldstein, J.; Hanks, L.M.; Millar, J.G.; D′amico, V. Species Richness and Phenology of Cerambycid Beetles in Urban Forest Fragments of Northern Delaware. Ann. Entomol. Soc. Am. 2015, 108, 251–262. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- McCoy, E.D. The distribution of insects along elevational gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maveety, S.A.; Browne, R.A.; Erwin, T.L. Carabidae diversity along an altitudinal gradient in a Peruvian cloud forest (Coleoptera). ZooKeys 2011, 147, 651–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzan, A.M.; Flinte, V.; Macedo, M.V.; Monteiro, R.F. Elevation and temporal distributions of Chrysomelidae in southeast Brazil with emphasis on the Galerucinae. ZooKeys 2015, 547, 103–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srisuka, W.; Takaoka, H.; Otsuka, Y.; Fukuda, M.; Thongsahuan, S.; Taai, K.; Choochote, W.; Saeung, A. Seasonal biodiversity of black flies (Diptera: Simuliidae) and evaluation of ecological factors influencing species distribution at Doi Pha Hom Pok National Park, Thailand. Acta Trop. 2015, 149, 212–219. [Google Scholar]
- Lefebvre, V.; Villemant, C.; Fontaine, C.; Daugeron, C. Altitudinal, temporal and trophic partitioning of flower-visitors in Alpine communities. Sci. Rep. 2018, 8, 4706. [Google Scholar] [CrossRef]
- Fontana, V.; Guariento, E.; Hilpold, A.; Niedrist, G.; Steinwandter, M.; Spitale, D.; Nascimbene, J.; Tappeiner, U.; Seeber, J. Species richness and beta diversity patterns of multiple taxa along an elevational gradient in pastured grasslands in the European Alps. Sci Rep. 2020, 10, 12516. [Google Scholar] [CrossRef] [PubMed]
- Wolda, H. Insect seasonality: Why? Ann. Rev. Ecol. Syst. 1988, 19, 1–18. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K.; Fluckinger, P.F. Forest edges are biodiversity hotspots: Also for Neuroptera. Acta Zool. Acad. Sci. Hung. 2020, 48, 75–87. [Google Scholar]
- Weiss, M.; Procházka, J.; Schlaghamerský, J.; Cizek, L. Fine-Scale Vertical Stratification and Guild Composition of Saproxylic Beetles in Lowland and Montane Forests: Similar Patterns despite Low Faunal Overlap. PLoS ONE 2016, 11, e0149506. [Google Scholar] [CrossRef] [Green Version]
- Costello, S.L.; Negrón, J.F.; Jacobi, W.J. Wood-boring insect abundance in fire-injured ponderosa pine. Agric. Forest Entomol. 2011, 13, 373–381. [Google Scholar] [CrossRef]
- Da Silva, N.R.P.; Frizzas, M.R.; Oliveira, C.M. Seasonality in insect abundance in the “Cerrado” of Goiás State, Brazil. Rev. Bras. Entomol. 2011, 55, 79–87. [Google Scholar]
- Ramos-Robles, M.; Vargas-Cardoso, O.R.; Corona-Lopez, A.M.; Flores-Palacios, A.; Toledo-Hernandez, V.H. Spatio-temporal variation of Cerambycidae- host tree interaction networks. PLoS ONE 2020, 15, e0228880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCravy, K.W. A review of sampling and monitoring methods for beneficial arthropods in agroecosystems. Insects 2018, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.; Calvin, D.; Mayhew, P. The effectiveness and optimal use of Malaise traps for monitoring parasitoid wasps. Insect Conserv. Divers. 2008, 1, 22–31. [Google Scholar] [CrossRef]
- Ngo, H.T.; Gibbs, J.; Griswold, T.; Packer, L. Evaluating bee (Hymenoptera: Apoidea) diversity using Malaise traps in coffee landscapes of Costa Rica. Can. Entomol. 2013, 145, 435–453. [Google Scholar] [CrossRef] [Green Version]
- Evans, A. Invertebrates: Malaise trapping Version 1.0. Available online: https://www.doc.govt.nz/globalassets/documents/science-and-technical/inventory-monitoring/im-toolbox-invertebrates-malaise-trapping.pdf (accessed on 31 December 2020).
- Grootaert, P.; Pollet, M.; Dekoninck, W.; van Achterberg, C. Sampling insects: General techniques, strategies and remarks. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories and Monitoring, Part 2; Eymann, J., Degreef, J., Häuser, C., Monje, J.C., Samyn, Y., VandenSpiegel, D., Eds.; Abc Taxa: Brussels, Belgium, 2010; Volume 8, pp. 377–399. [Google Scholar]
- Van Achterberg, C.; Shaw, M.R.; Grootaert, P. Flight interception traps for arthropods. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories and Monitoring, Part 2; Eymann, J., Degreef, J., Häuser, C., Monje, J.C., Samyn, Y., VandenSpiegel, D., Eds.; Abc Taxa: Brussels, Belgium, 2010; Volume 8, pp. 423–462. [Google Scholar]
- Boulter, S.L.; Lambkin, C.L.; Starick, N.T. Assessing the abundance of seven major arthropod groups along an altitudinal gradient and across seasons in subtropical rainforest. Mem. Qld. Mus. Nat. 2011, 55, 303–313. [Google Scholar]
- Geiger, M.; Moriniere, J.; Hausmann, A.; Haszprunar, G.; Wägele, W.; Hebert, P.; Rulik, B. Testing the Global Malaise Trap Program–How well does the current barcode reference library identify flying insects in Germany? Biodivers. Data J. 2016, 4, e10671. [Google Scholar] [CrossRef] [Green Version]
- Henry, S.C.; McQuillan, P.B.; Kirkpatrick, J.B. An Alpine Malaise trap. ALP Entomol. 2018, 2, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, D.; Hartop, E.; Forshage, M.; Jaschhof, M.; Ronquist, F. The Swedish Malaise Trap Project: A 15 Year Retrospective on a countrywide insect inventory. Biodivers. Data J. 2020, 8, e47255. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yotkham, S.; Suttiprapan, P.; Likhitrakarn, N.; Sulin, C.; Srisuka, W. Biodiversity and Spatiotemporal Variation of Longhorn Beetles (Coleoptera: Cerambycidae) in Tropical Forest of Thailand. Insects 2021, 12, 45. https://doi.org/10.3390/insects12010045
Yotkham S, Suttiprapan P, Likhitrakarn N, Sulin C, Srisuka W. Biodiversity and Spatiotemporal Variation of Longhorn Beetles (Coleoptera: Cerambycidae) in Tropical Forest of Thailand. Insects. 2021; 12(1):45. https://doi.org/10.3390/insects12010045
Chicago/Turabian StyleYotkham, Sirapat, Piyawan Suttiprapan, Natdanai Likhitrakarn, Chayanit Sulin, and Wichai Srisuka. 2021. "Biodiversity and Spatiotemporal Variation of Longhorn Beetles (Coleoptera: Cerambycidae) in Tropical Forest of Thailand" Insects 12, no. 1: 45. https://doi.org/10.3390/insects12010045