NADPH Oxidase 5 Is Essential for Molting and Oviposition in a Rice Planthopper Nilaparvata lugens

 ,
, 
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Development-Stage and Tissue Sample Collection
2.3. Developmental Stage- and Tissue-Expression Patterns of Nox5 by Quantitative Real-Time PCR Analysis
2.4. RNA Interference (RNAi)
2.5. Observation of Oviposition and Egg Hatching
2.6. Measurement of H2O2 Production
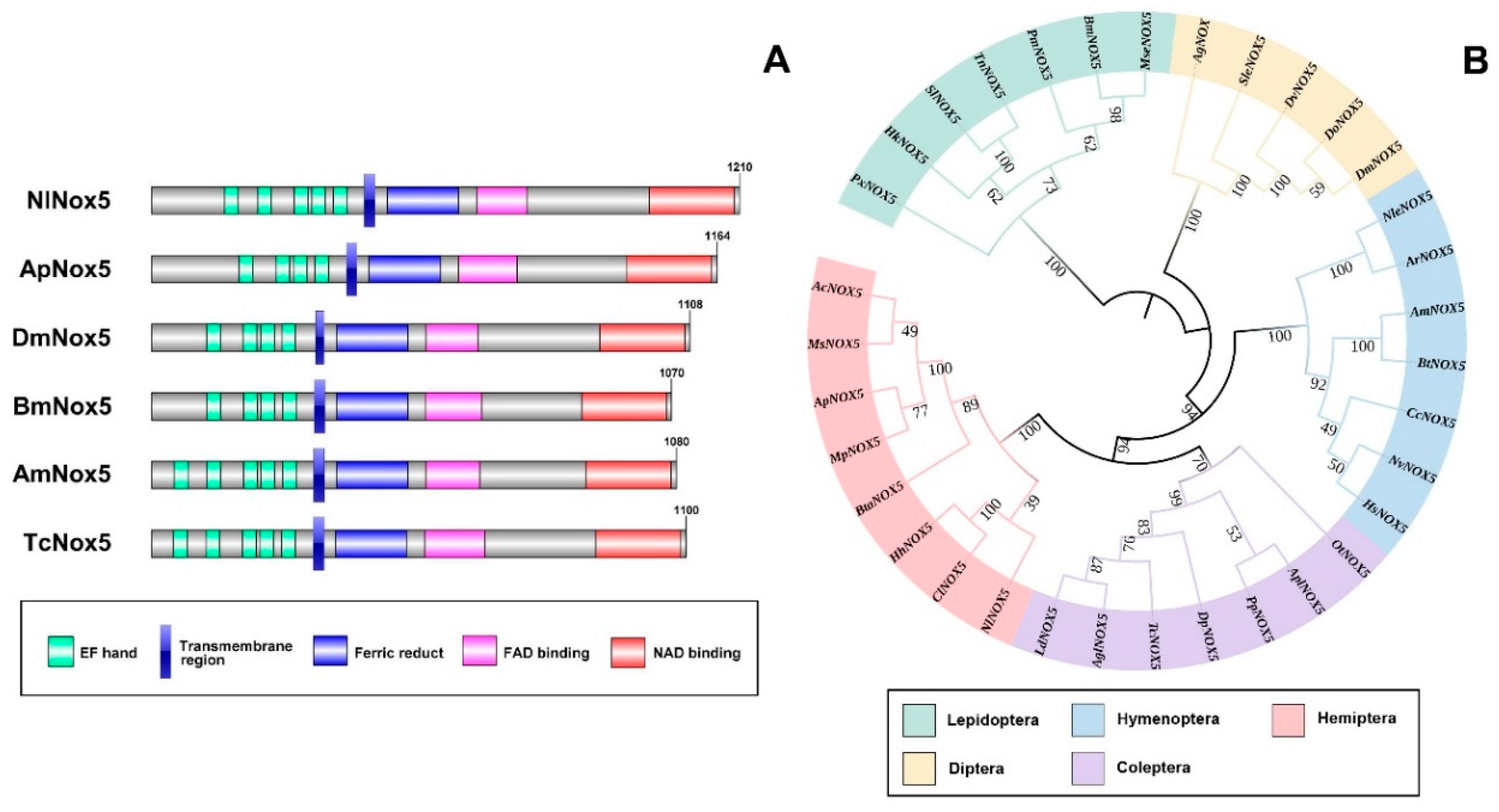
2.7. Analysis of Evolutionary Relationships
2.8. Statistical Analyses
2.9. Availability of Supporting Data
3. Results
3.1. Identification of an NADPH Oxidase 5 Gene in N. lugens
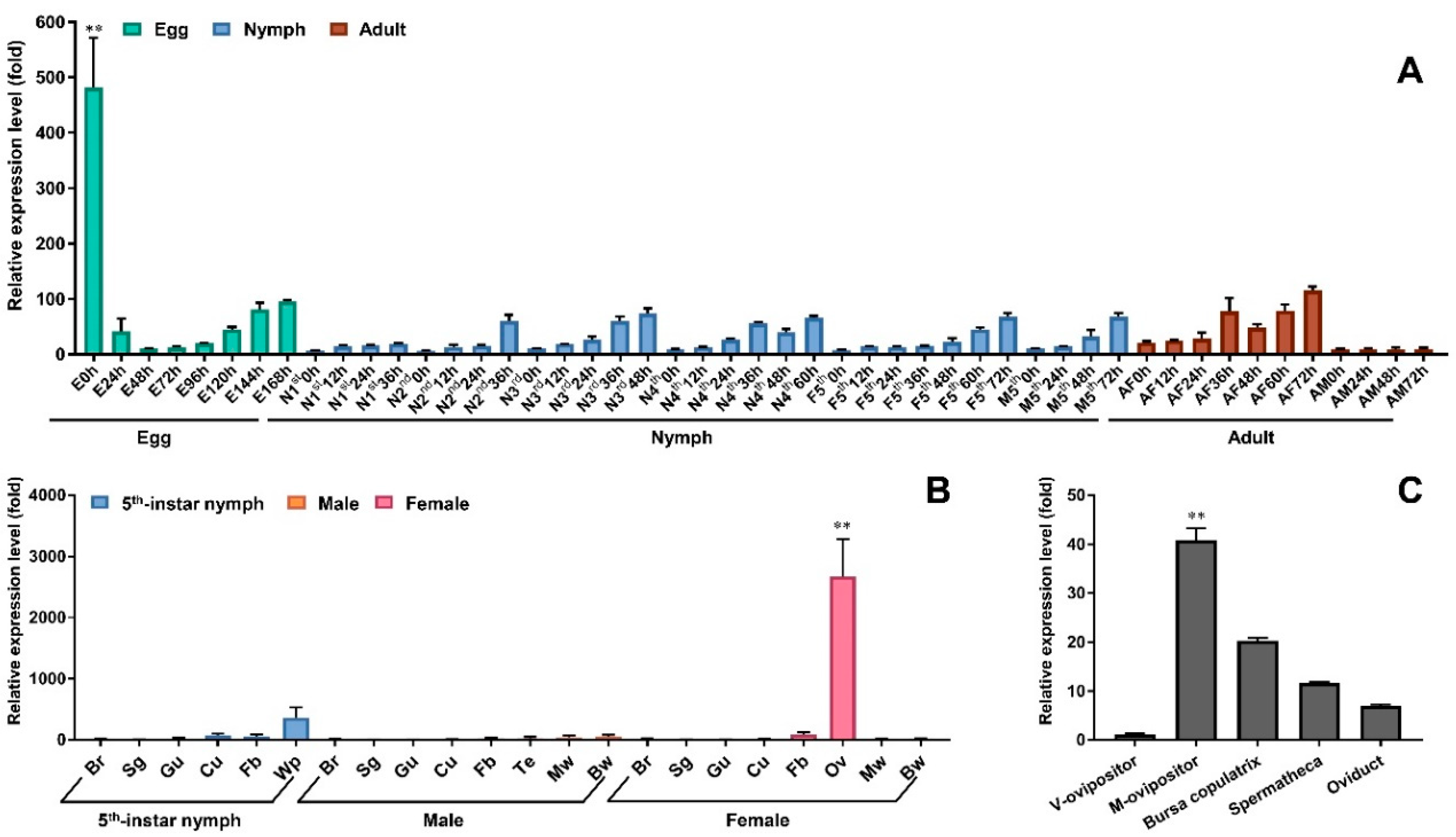
3.2. Temporospatial Expression Patterns of Nox5 Gene in N. lugens
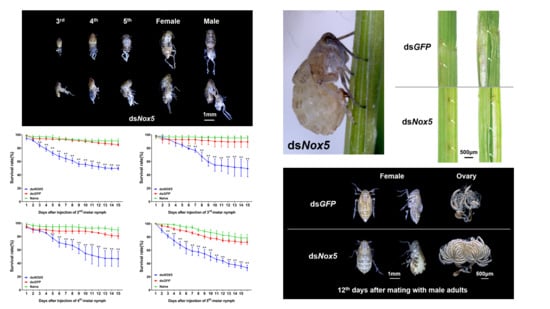
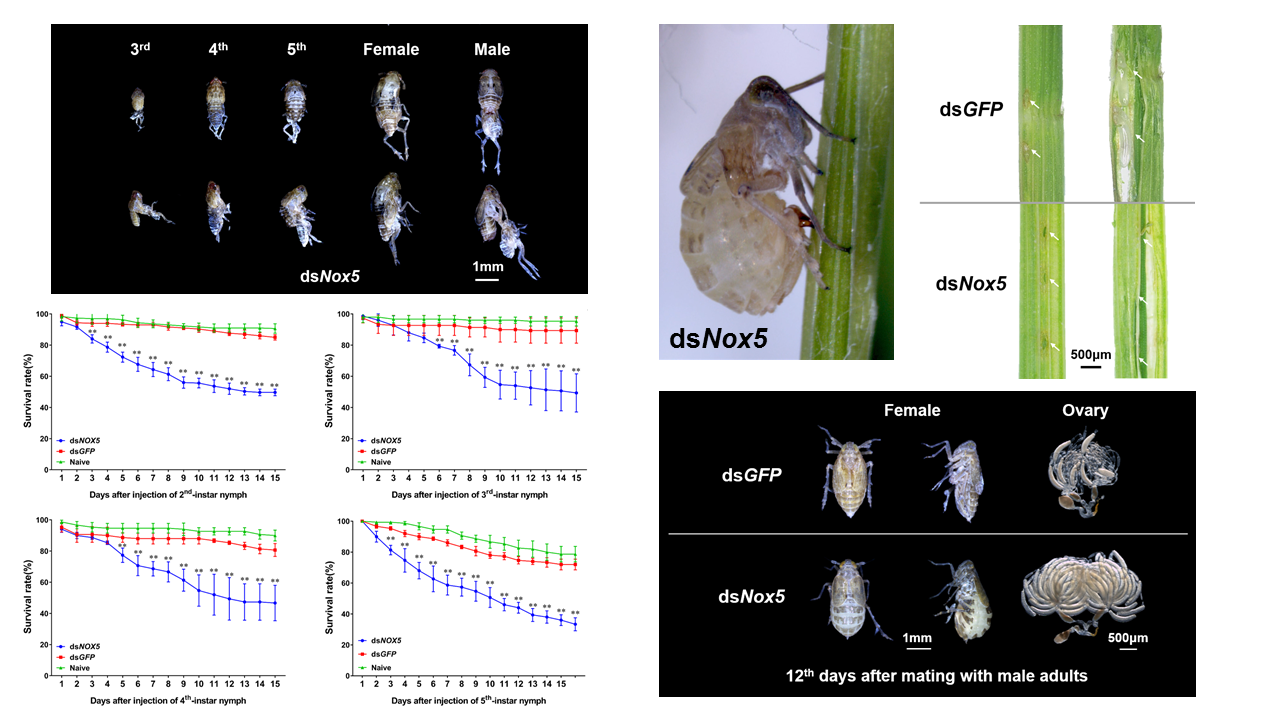
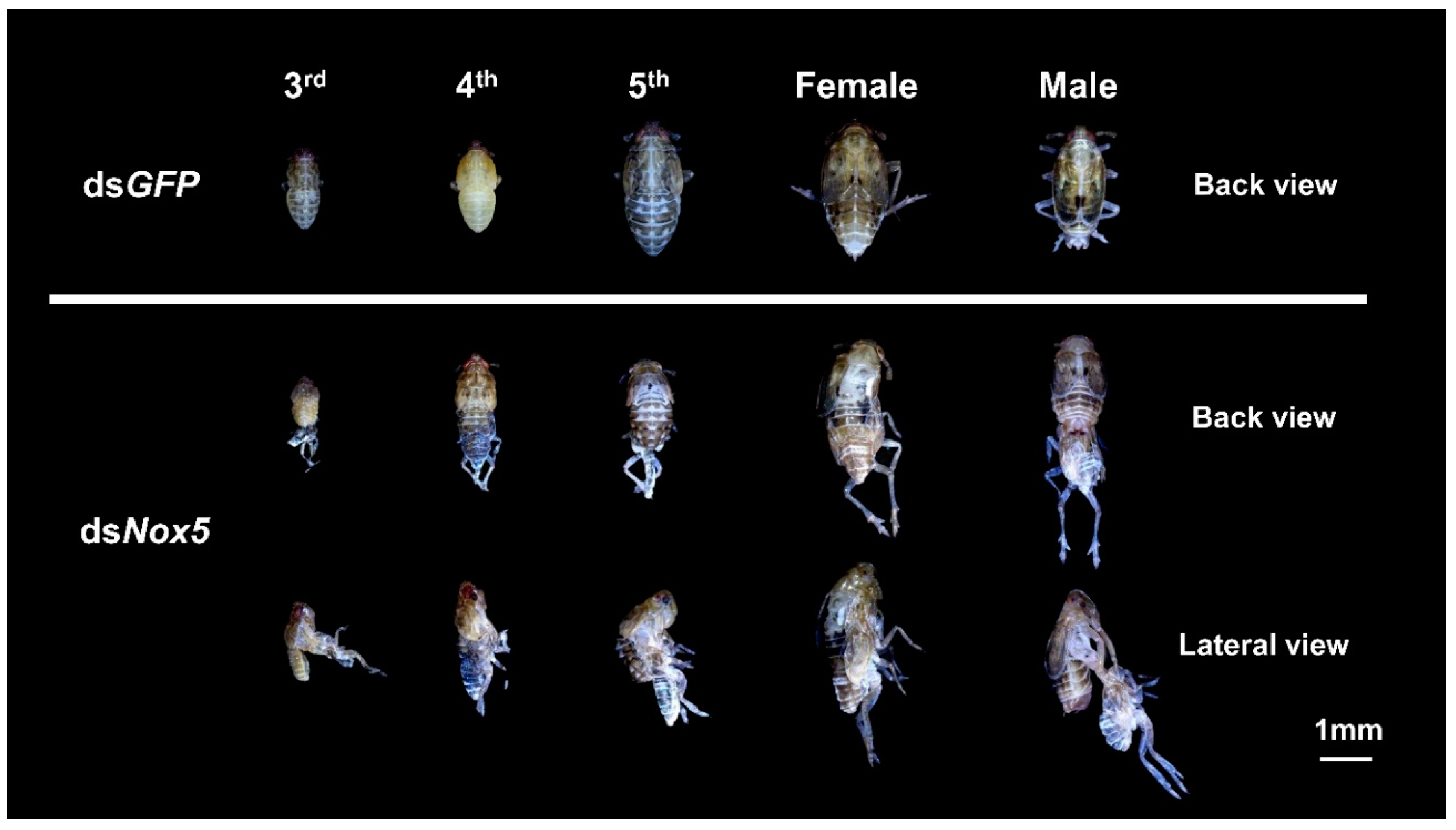
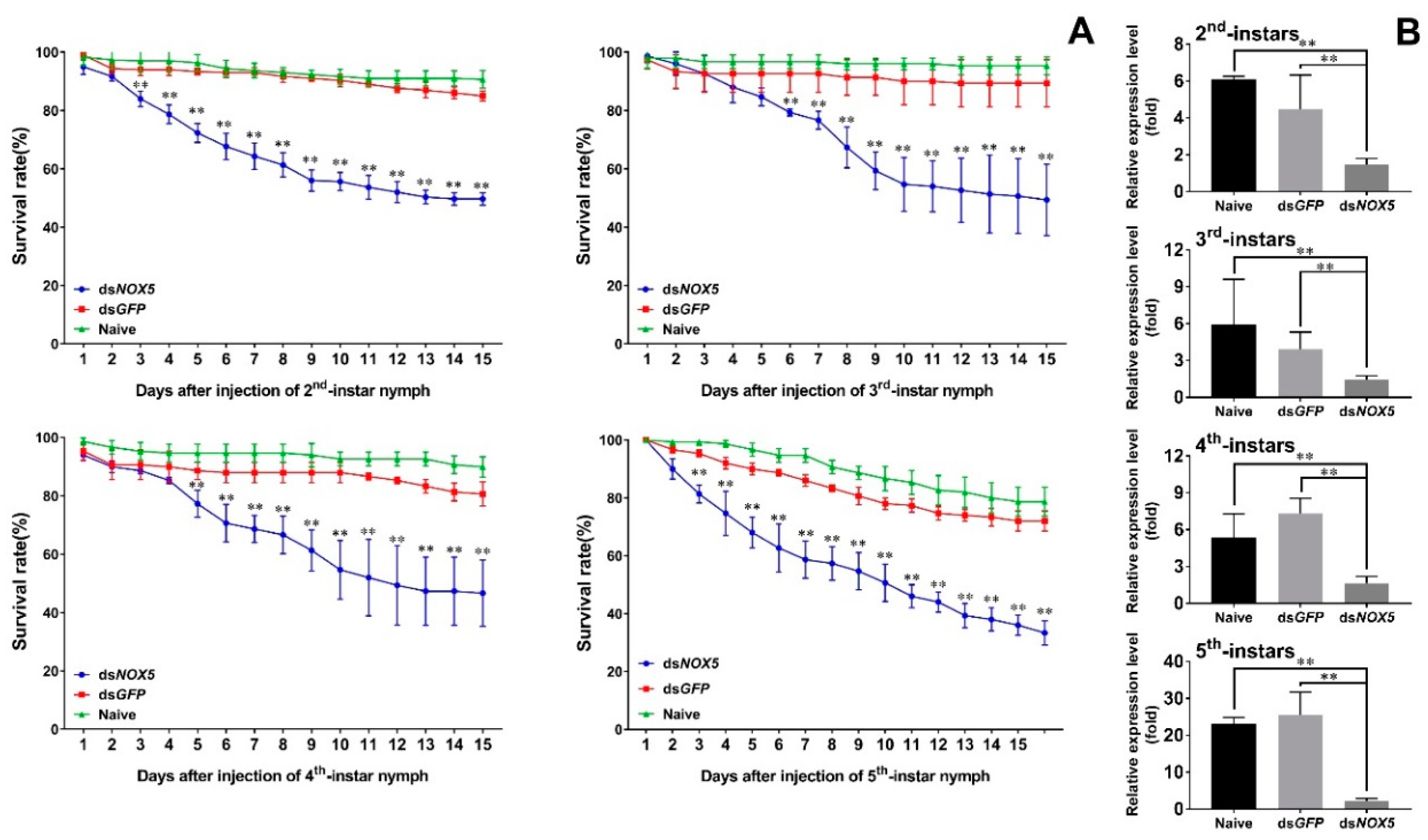
3.3. Effect of Nox5 Knockdown on Development of Nymphs
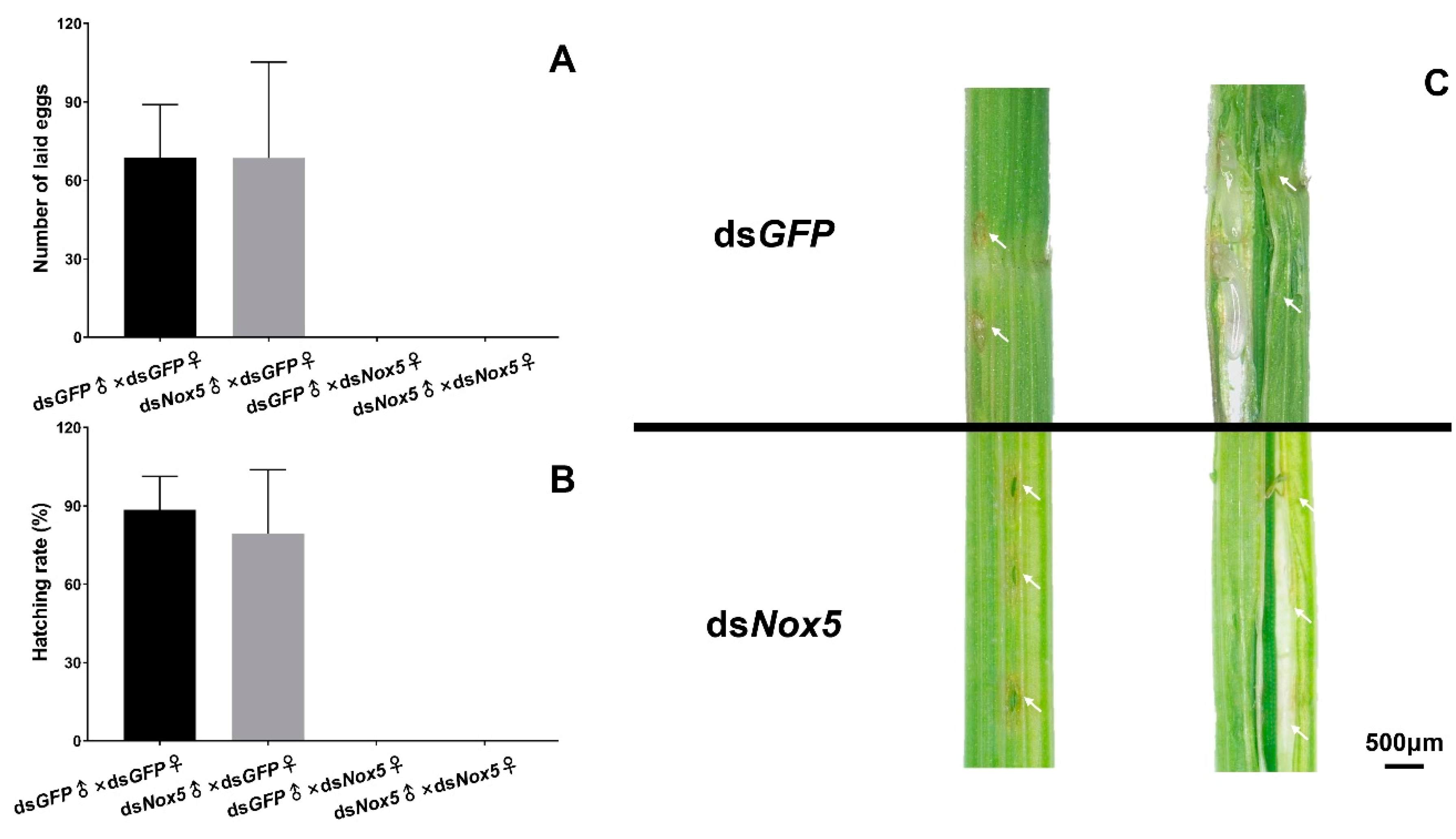
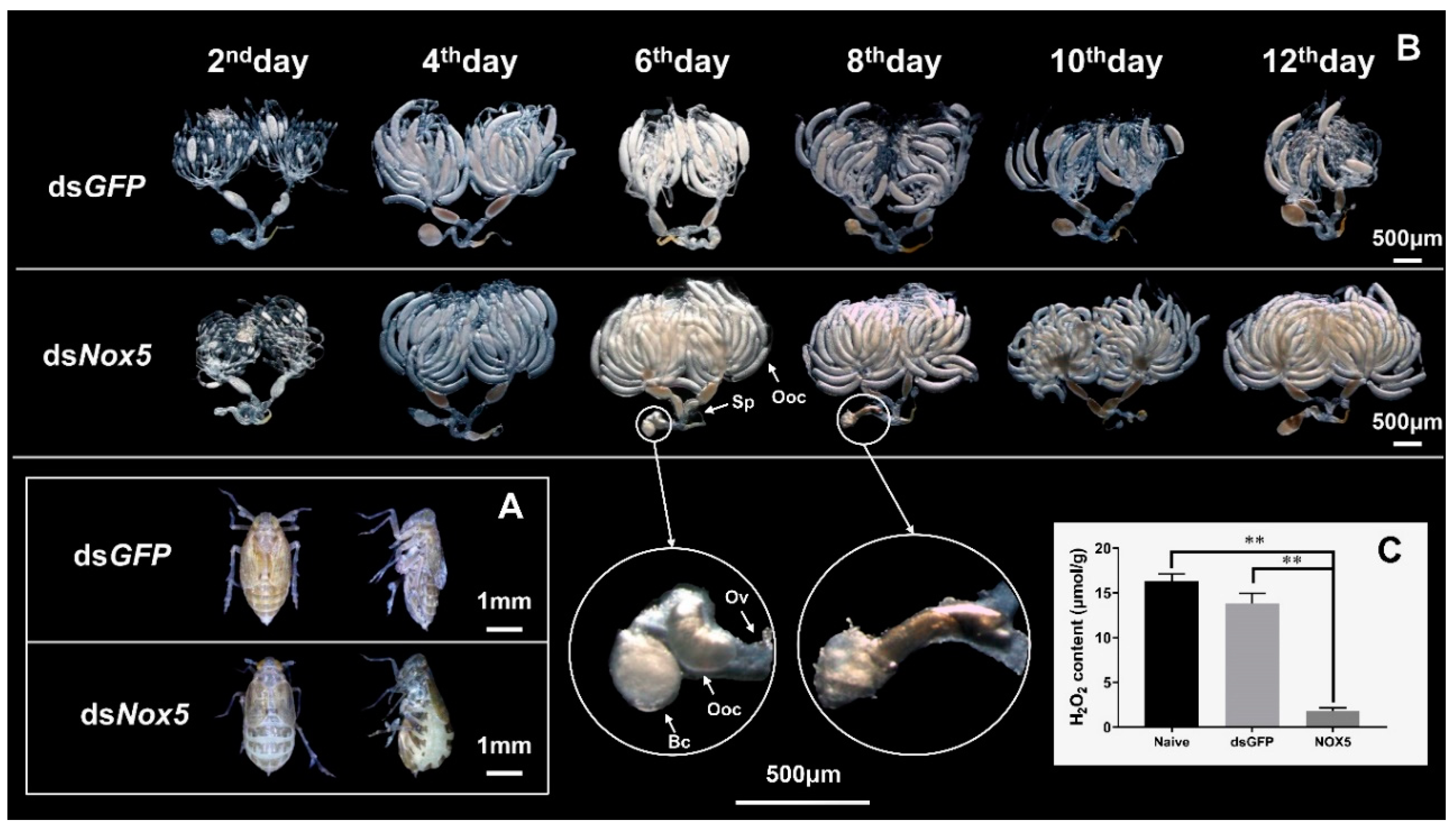
3.4. Effect of Nox5 Knockdown on Oviposition of Female Adults
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bao, Y.Y.; Zhang, C.X. Recent advances in molecular biology research of a rice pest, the brown planthopper. J. Integr. Agric. 2019, 18, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Minkenberg, O.; Tatar, M.; Rosenheim, J.A. Egg load as a major source of variability in insect foraging and oviposition behavior. Oikos 1992, 65, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Deady, L.D.; Shen, W.; Mosure, S.A.; Spradling, A.C.; Sun, J. Matrix metalloproteinase 2 is required for ovulation and corpus luteum formation in Drosophila. PLoS Genet. 2015, 11, e1004989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Young, J.F.; Sun, J. NADPH oxidase-generated reactive oxygen species in mature follicles are essential for Drosophila ovulation. Proc. Natl. Acad. Sci. USA 2018, 115, 7765–7770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.H.; Shen, Y.; Li, D.T.; Huang, H.J.; Lu, J.B.; Zhang, C.X. A mucin-like protein is essential for oviposition in Nilaparvata lugens. Front. Physiol. 2019, 10, 551. [Google Scholar] [CrossRef]
- Lou, Y.H.; Lu, J.B.; Li, D.T.; Ye, Y.X.; Luo, X.M.; Zhang, C.X. Amelogenin domain-containing NlChP38 is necessary for normal ovulation in the brown planthopper. Insect Mol. Biol. 2019, 28, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.Y.; Wang, Y.; Wu, W.J.; Zhao, D.; Xue, J.; Zhang, B.Q.; Shen, Z.C.; Zhang, C.X. De novo intestine-specific transcriptome of the brown planthopper Nilaparvata lugens revealed potential functions in digestion, detoxification and immune response. Genomics 2012, 99, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.Y.; Qu, L.Y.; Zhao, D.; Chen, L.B.; Jin, H.Y.; Xu, L.M.; Cheng, J.A.; Zhang, C.X. The genome- and transcriptome-wide analysis of innate immunity in the brown planthopper, Nilaparvata lugens. BMC Genom. 2013, 14, 160. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.Y.; Qin, X.; Yu, B.; Chen, L.B.; Wang, Z.C.; Zhang, C.X. Genomic insights into the serine protease gene family and expression profile analysis in the planthopper, Nilaparvata lugens. BMC Genom. 2014, 15, 507. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Zhou, X.; Zhang, C.X.; Yu, L.L.; Fan, H.W.; Wang, Z.; Xu, H.J.; Xi, Y.; Zhu, Z.R.; Zhou, W.W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genome Biol. 2014, 15, 122014. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Bao, Y.Y.; Li, B.l.; Cheng, Y.B.; Peng, Z.Y.; Liu, H.; Xu, H.J.; Zhu, Z.R.; Lou, Y.G.; Cheng, J.A.; et al. Transcriptome analysis of the brown planthopper Nilaparvata lugens. PLoS ONE 2010, 5, e14233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babior, B.M. NADPH oxidase. Curr. Opin. Immunol. 2004, 16, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Ritsick, D.R.; Edens, W.A.; Finnerty, V.; Lambeth, J.D. Nox regulation of smooth muscle contraction. Free Radic. Biol. Med. 2007, 43, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.I.; Griendling, K.K. Nox proteins in signal transduction. Free Radic. Biol. Med. 2009, 47, 1239–1253. [Google Scholar] [CrossRef] [Green Version]
- Lambeth, J.D. Nox enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc. Natl. Acad. Sci. USA 2011, 108, 1462–1467. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.J.; Chen, T.; Ma, X.F.; Xue, J.; Pan, P.L.; Zhang, X.C.; Cheng, J.A.; Zhang, C.X. Genome-wide screening for components of small interfering RNA (siRNA) and micro-RNA (miRNA) pathways in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Mol. Biol. 2013, 22, 635–647. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, Y.Z.; Ogihara, M.H.; Takeshima, M.; Fujinaga, D.; Liu, C.W.; Zhu, Z.; Kataoka, H.; Bao, Y.Y. Functional analysis of ecdysteroid biosynthetic enzymes of the rice planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2020, 123, 103428. [Google Scholar] [CrossRef]
- Huang, H.J.; Liu, C.W.; Huang, X.H.; Zhou, X.; Zhuo, J.C.; Zhang, C.X.; Bao, Y.Y. Screening and functional analyses of Nilaparvata lugens salivary proteome. J. Proteome Res. 2016, 15, 1883–1896. [Google Scholar] [CrossRef]
- Xu, L.; Huang, H.J.; Zhou, X.; Liu, C.W.; Bao, Y.Y. Pancreatic lipase-related protein 2 is essential for egg hatching in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2017, 26, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, Y.Z.; Lou, W.H.; Zhang, C.X. Vitellogenin and vitellogenin-like genes in the brown planthopper. Front. Physiol. 2019, 10, 1181. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lai, Y.L.; Wang, G.D.; Chen, H.; Li, F.; Wang, S.B. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Peng, L.Y.; Wang, Z.C.; Wang, W.; Zhu, Z.; Huang, X.H.; Chen, L.B.; Song, Q.S.; Bao, Y.Y. Identification of novel antimicrobial peptides from rice planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2019, 113, 103215. [Google Scholar] [CrossRef]
- Lam, P.L.; Wong, M.M.; Hung, L.K.; Yung, L.H.; Tang, J.C.O.; Lam, K.H.; Chung, P.Y.; Wong, W.Y.; Ho, Y.W.; Wong, R.S.M.; et al. Miconazole and terbinafine induced reactive oxygen species accumulation and topical toxicity in human keratinocytes. Drug Chem. Toxicol. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.-Y.; Dai, Z.-W.; Yang, R.-R.; Zhu, Z.; Wang, W.; Zhou, X.; Bao, Y.-Y. NADPH Oxidase 5 Is Essential for Molting and Oviposition in a Rice Planthopper Nilaparvata lugens. Insects 2020, 11, 642. https://doi.org/10.3390/insects11090642
Peng L-Y, Dai Z-W, Yang R-R, Zhu Z, Wang W, Zhou X, Bao Y-Y. NADPH Oxidase 5 Is Essential for Molting and Oviposition in a Rice Planthopper Nilaparvata lugens. Insects. 2020; 11(9):642. https://doi.org/10.3390/insects11090642
Chicago/Turabian StylePeng, Lu-Yao, Zhen-Wei Dai, Rui-Rui Yang, Zhen Zhu, Wei Wang, Xiang Zhou, and Yan-Yuan Bao. 2020. "NADPH Oxidase 5 Is Essential for Molting and Oviposition in a Rice Planthopper Nilaparvata lugens" Insects 11, no. 9: 642. https://doi.org/10.3390/insects11090642