Diapause Regulation in Newly Invaded Environments: Termination Timing Allows Matching Novel Climatic Constraints in the Box Tree Moth, Cydalima perspectalis (Lepidoptera: Crambidae)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
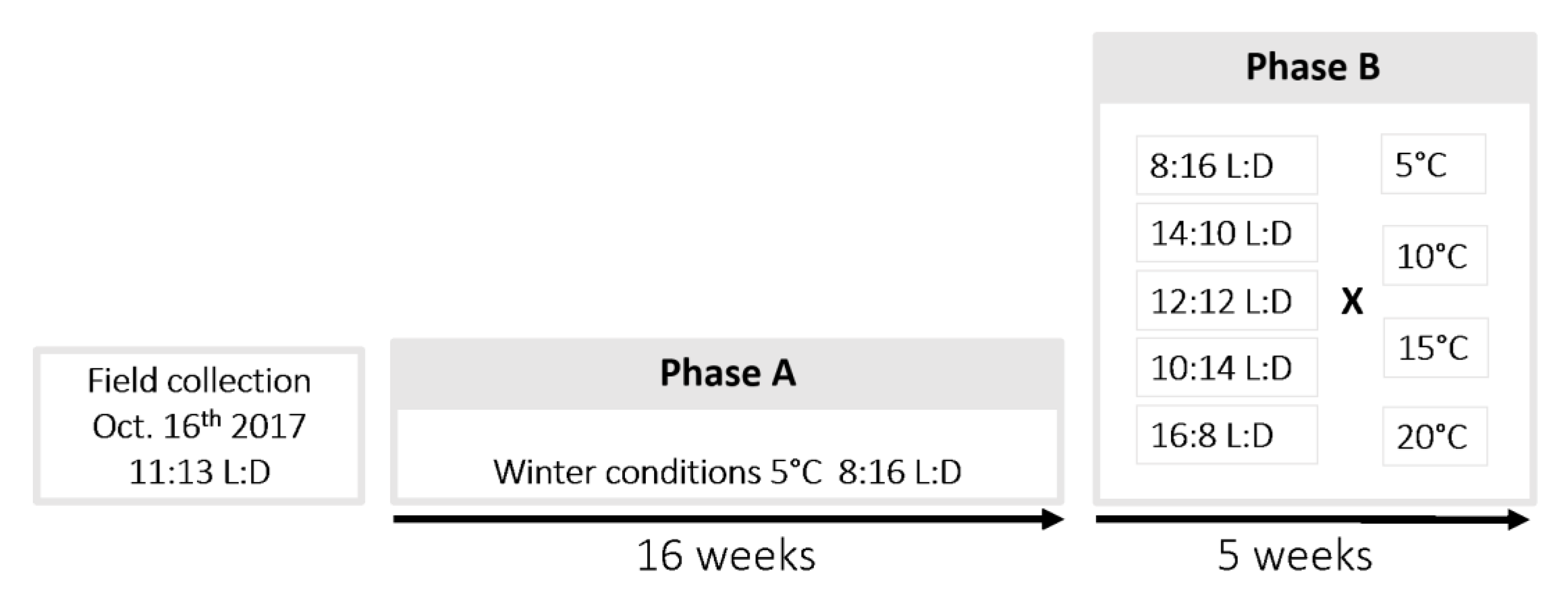
2.2. Conditions during Winter
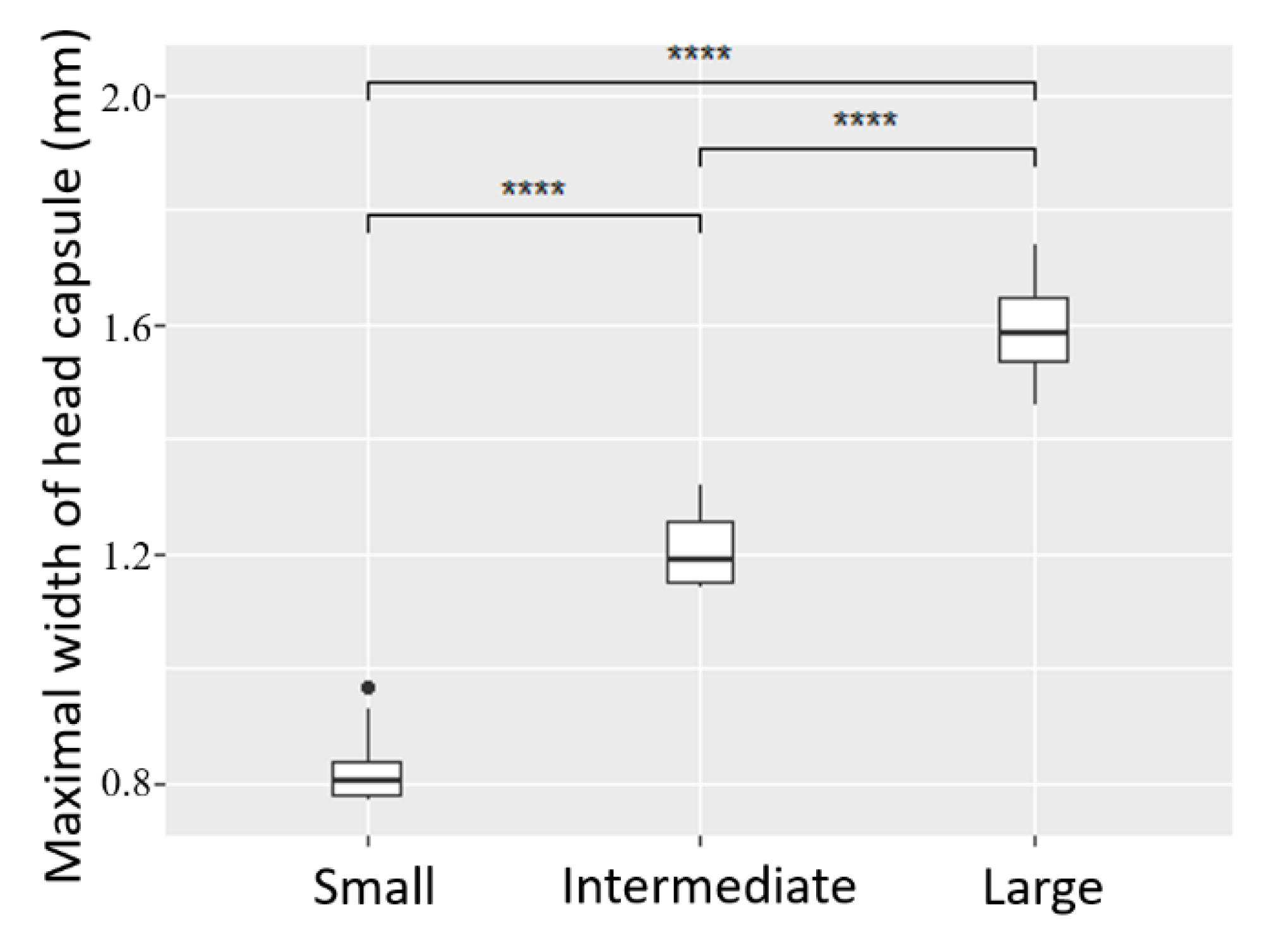
2.3. Instar Determination
2.4. Diapause Termination Conditions
2.5. Activity Monitoring
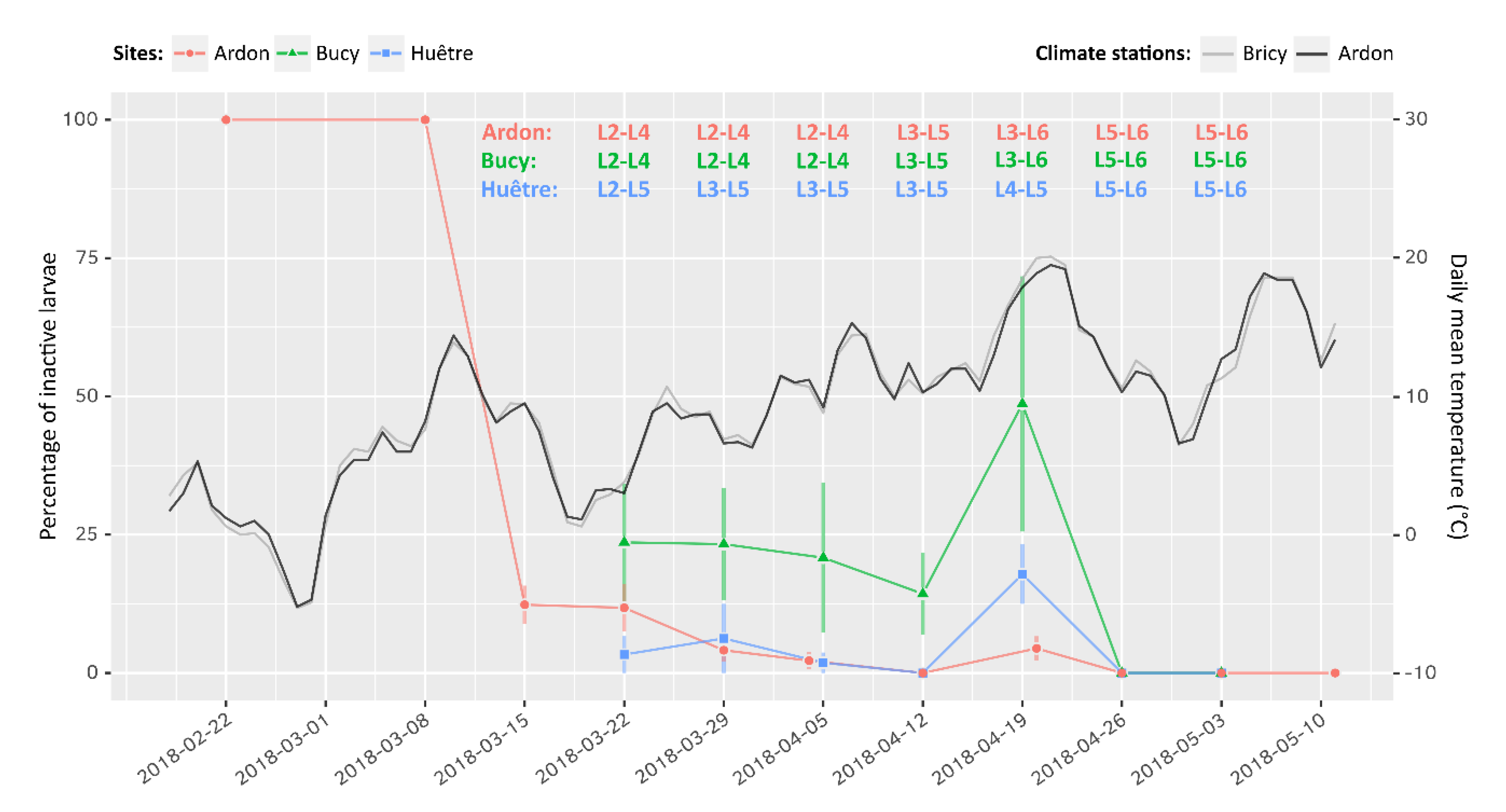
2.6. Monitoring the Return to an Active Phase in the Field
2.7. Statistical Analyses
3. Results
3.1. Instar Determination
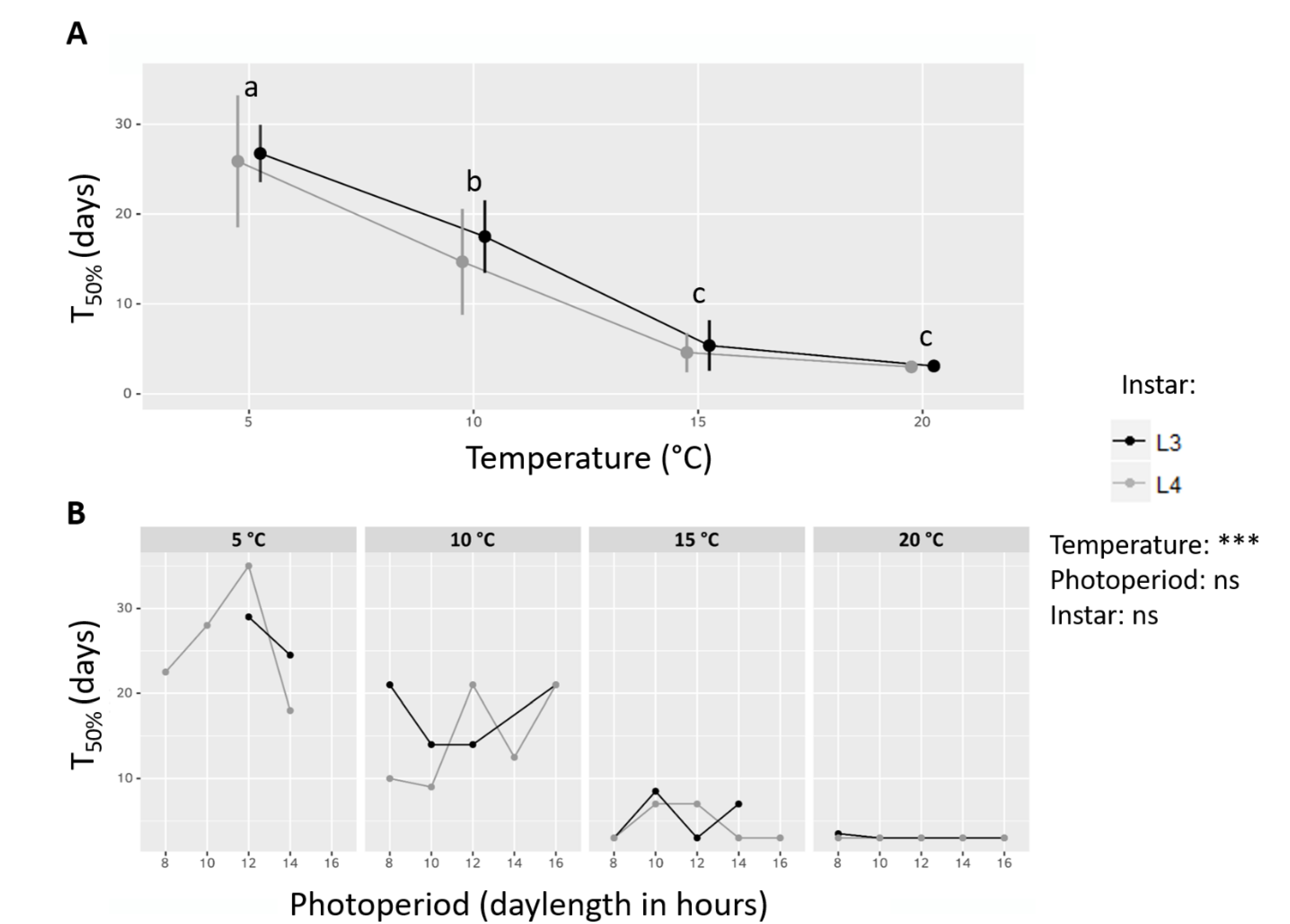
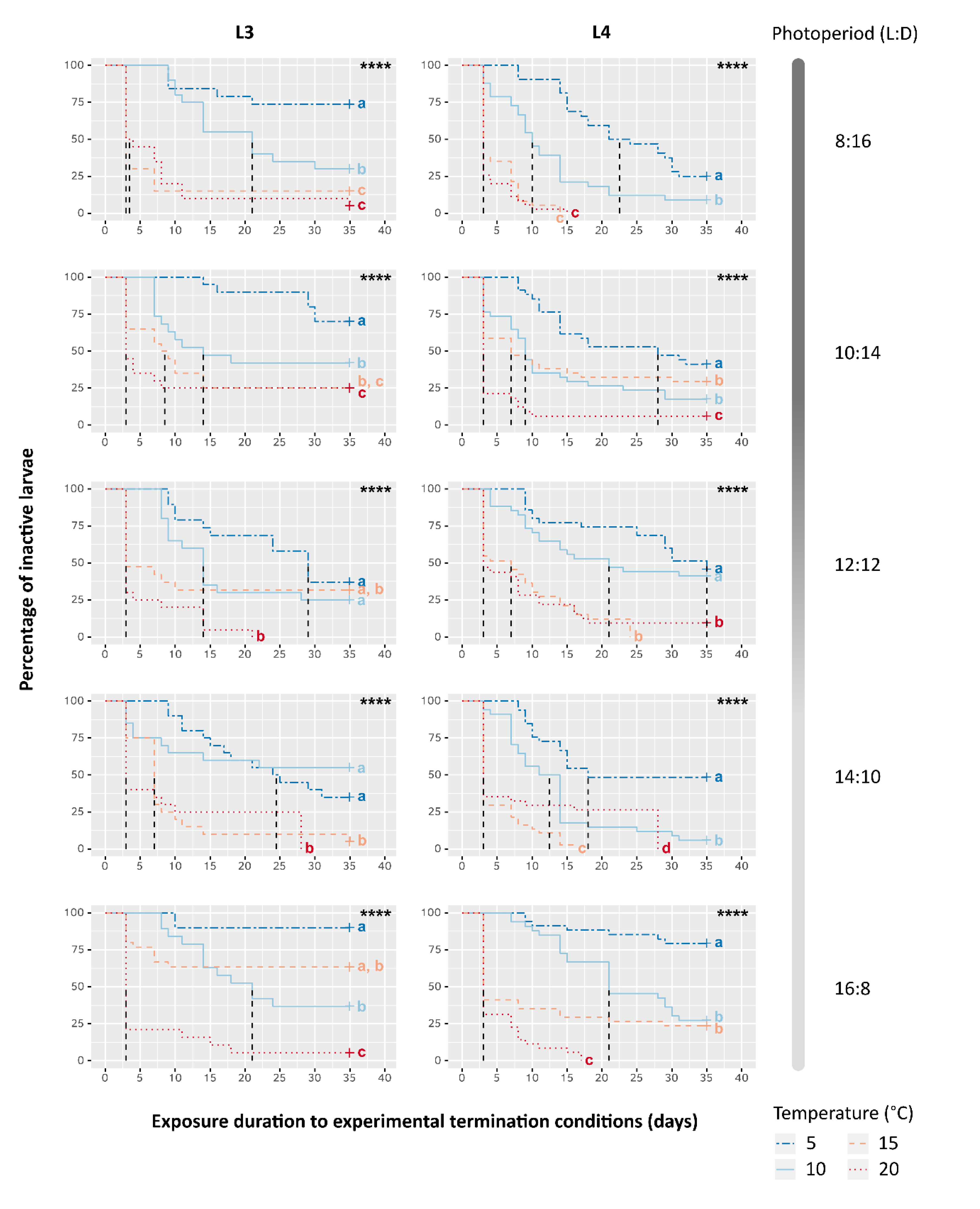
3.2. Effects of Temperature, Photoperiod and Larval Instar on Survival and Activity
3.3. Activity and Phenology in Nature
4. Discussion
4.1. The Influence of Temperature on Activity Resumption
4.2. Full Development in Warm Condition Only
4.3. Does the Box Tree Moth Incur a Post-Diapause Stage that May Improve Flexibility to Varying Seasonal Constraints?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Falk-Petersen, J.; Bøhn, T.; Sandlund, O.T. On the Numerous Concepts in Invasion Biology. Biol. Invasions 2006, 8, 1409–1424. [Google Scholar] [CrossRef] [Green Version]
- Roques, A. Taxonomy, time and geographic patterns. Chapter 2. BioRisk 2010, 4, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Roques, A.; Auger-Rozenberg, M.-A.; Blackburn, T.M.; Garnas, J.; Pyšek, P.; Rabitsch, W.; Richardson, D.M.; Wingfield, M.J.; Liebhold, A.M.; Duncan, R.P. Temporal and interspecific variation in rates of spread for insect species invading Europe during the last 200 years. Biol. Invasions 2016, 18, 907–920. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Papp, V.; Ladányi, M.; Vétek, G. Temperature-dependent development of Aproceros leucopoda (Hymenop-tera: Argidae), an invasive pest of elms in Europe. J. Appl. Entomol. 2018, 142, 589–597. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Nahrung, H.F.; Swain, A.J. Strangers in a strange land: Do life history traits differ for alien and native colonisers of novel environments? Biol. Invasions 2015, 17, 699–709. [Google Scholar] [CrossRef]
- Fahrner, S.; Aukema, B.H. Correlates of spread rates for introduced insects. Global Ecol. Biogeogr. 2018, 27, 734–743. [Google Scholar] [CrossRef]
- Renault, D.; Laparie, M.; McCauley, S.J.; Bonte, D. Environmental Adaptations, Ecological Filtering, and Dispersal Central to Insect Invasions. Annu. Rev. Entomol. 2018, 63, 345–368. [Google Scholar] [CrossRef]
- Kornberg, H.; Crawley, M.; Lawton, J.; Usher, M.; Southwood, R.; O’Connor, R.; Gibbs, A. The Population Biology of Invaders: Discussion. Philos. Trans. R. Soc. B 1986, 314, 730–731. [Google Scholar] [CrossRef]
- Ehrlich, P. Attributes of invaders and the invading processes vertebrates. In Biological Invasions: A Global Perspective; Drake, J.A., Mooney, H.A., di Castri, F., Groves, R.H., Kruger, F.J., Rejmánek, M., Williamson, M., Eds.; Wiley: Chichester, UK, 1989; pp. 315–328. [Google Scholar]
- Gray, A.J.; Mack, R.N.; Harper, J.L.; Usher, M.B.; Joysey, K.; Kornberg, H. Do Invading Species have Definable Genetic Characteristics? [and Discussion]. Philos. T. Roy. Soc. B 1986, 314, 655–674. [Google Scholar] [CrossRef]
- Danks, H.V. Insect Dormancy: An Ecological Perspective. Q. Rev. Biol. 1987, 62, 452–453. [Google Scholar] [CrossRef]
- Hodek, I. Controversial aspects of diapause development. Eur. J. Entomol. 2013, 99, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Roff, D.A.; Deland, J.-P. The Joint Evolution of Diapause and Insecticide Resistance: A Test of an Optimality Model. Ecology 1995, 76, 1497–1505. [Google Scholar] [CrossRef]
- Panov, V.E.; Piotr, I.; Riccardi, N. Role of diapause in dispersal and invasion success by aquatic invertebrates. J. Limnol. 2004, 63, 56–69. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, P.A. Photoperiodic Diapause and the Establishment of Aedes albopictus (Diptera: Culicidae) in North America. J. Med. Entomol. 2016, 53, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Shinkanji, N. The life-cycle of the box-tree pyralid, Glyphodes perspectalis (Walker) (Lepidoptera: Pyralidae). III. Photoperiodic induction of larval diapause. Jpn. J. Appl. Entomol. Z. 1993, 37, 45–51. [Google Scholar] [CrossRef]
- Wan, H.; Haye, T.; Kenis, M.; Nacambo, S.; Xu, H.; Zhang, F.; Li, H. Biology and natural enemies of Cydalima perspectalis in Asia: Is there biological control potential in Europe? J. Appl. Entomol. 2014, 138, 715–722. [Google Scholar] [CrossRef]
- Bras, A.; Avtzis, D.N.; Kenis, M.; Li, H.; Vétek, G.; Bernard, A.; Courtin, C.; Rousselet, J.; Roques, A.; Auger-Rozenberg, M.-A. A complex invasion story underlies the fast spread of the invasive box tree moth (Cydalima perspectalis) across Europe. J. Pest Sci. 2019, 92, 1187–1202. [Google Scholar] [CrossRef]
- Nacambo, S.; Leuthardt, F.L.G.; Wan, H.; Li, H.; Haye, T.; Baur, B.; Weiss, R.M.; Kenis, M. Development characteristics of the box-tree moth Cydalima perspectalis and its potential distribution in Europe. J. Appl. Entomol. 2014, 138, 14–26. [Google Scholar] [CrossRef]
- Frank, S. Watch for potential new boxwood pest report. Purdue Lansc. Rep. 2019, 19, 1–4. [Google Scholar]
- Bras, A. Identification des Facteurs Sous-Tendant L’invasion Fulgurante d’un Insecte Asiatique en Europe, la Pyrale du Buis: Approche Génétique et Biologique. Ph.D. Thesis, Université d’Orléans, Orléans, France, 2018. [Google Scholar]
- Eschen, R.; Douma, J.C.; Grégoire, J.-C.; Mayer, F.; Rigaux, L.; Potting, R.P.J. A risk categorisation and analysis of the geographic and temporal dynamics of the European import of plants for planting. Biol. Invasions 2017, 19, 3243–3257. [Google Scholar] [CrossRef]
- Székely, L.; Dinc, V.; Mihai, C. Cydalima perspectalis (Walker, 1859), a new species for the Romanian fauna (Lepidoptera: Crambidae: Spilomelinae). Bull. Inf. Entomol. 2011, 22, 73–78. [Google Scholar]
- Nacambo, S. Parasitisme, Développement, Modèle Climatique et Impact de Cydalima Perspectalis en Europe. Master Thesis, University of Neuchâtel, Neuchâtel, Switzerland, 2012. [Google Scholar]
- Fang, J.; Wang, Z.; Tang, Z. Atlas of Woody Plants in China. Distribution and Climate; Springer: Berlin, Germany, 2011; pp. 1–1902. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Hodek, I. Diapause development, diapause termination and the end of diapause. Eur. J. Entomol. 1996, 93, 475–487. [Google Scholar]
- Tabone, E.; Enriquez, T.; Giorgi, C.; Venard, M.; Colombel, E.; Gaglio, F.; Buradino, M.; Martin, J.-C. Mieux connaître la pyrale du buis Cydalima perspectalis. Phytoma 2015, 685, 18–20. [Google Scholar]
- R Core Team. R: A Langage and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www.R-project.org (accessed on 11 September 2020).
- Koštál, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef]
- Xiao, H.J. Photoperiod and temperature response of diapause induction in Diaphania perspectalis (Lepidoptera: Pyralidae). Chin. J. Entomol. 2011, 48, 116–120. [Google Scholar]
- Koštál, V.; Mollaei, M.; Schöttner, K. Diapause induction as an interplay between seasonal token stimuli, and modifying and directly limiting factors: Hibernation in Chymomyza costata: Environmental factors inducing diapause. Physiol. Entomol. 2016, 41, 344–357. [Google Scholar] [CrossRef]
- Philogène, B.J.R. Diapause. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1206–1210. [Google Scholar]
- Cheng, W.; Long, Z.; Zhang, Y.; Liang, T.; Zhu-Salzman, K. Effects of temperature, soil moisture and photoperiod on diapause termination and post-diapause development of the wheat blossom midge, Sitodiplosis mosellana (Géhin) (Diptera: Cecidomyiidae). J. Insect Physiol. 2017, 103, 78–85. [Google Scholar] [CrossRef] [PubMed]
- López, C.; Eizaguirre, M. Diapause and biological cycle of Cydalima perspectalis (Walker) in the eastern Pyrenees. J. Appl. Entomol. 2019, 143, 1096–1104. [Google Scholar] [CrossRef]
- Chippendale, G.M.; Reddy, A.S. Temperature and photoperiodic regulation of diapause of the southwestern corn borer, Diatraea grandiosella. J. Insect Physiol. 1973, 19, 1397–1408. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Tsitsipis, J.A.; Karandinos, M.G. Diapause Termination in Sesamia nonagrioides (Lepidoptera: Noctuidae) Under Laboratory and Field Conditions. Environ. Entomol. 1998, 27, 53–58. [Google Scholar] [CrossRef]
- Li, A.; Xue, F.; Hua, A.; TANG, J. Photoperiodic clock of diapause termination in Pseudopidorus fasciata (Lepidoptera: Zygaenidae). Eur. J. Entomol. 2003, 100, 287–293. [Google Scholar] [CrossRef]
- Brunnarius, J.; Dumortier, B. Existence of a light-sensitive phase in the photoperiodic termination of diapause in Pieris brassicae L. (Insecta: Lepidoptera) and comparison with diapause induction. J. Comp. Physiol. A 1984, 155, 161–169. [Google Scholar] [CrossRef]
- Xiao, H.-J.; Mou, F.-C.; Zhu, X.-F.; Xue, F.-S. Diapause induction, maintenance and termination in the rice stem borer Chilo suppressalis (Walker). J. Insect Physiol. 2010, 56, 1558–1564. [Google Scholar] [CrossRef]
- Jiang, X.F.; Huang, S.H.; Luo, L.Z.; Liu, Y.; Zhang, L. Diapause termination, post-diapause development and reproduction in the beet webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). J. Insect Physiol. 2010, 56, 1325–1331. [Google Scholar] [CrossRef]
- Yang, H.-Z.; Tu, X.-Y.; Xia, Q.-W.; He, H.-M.; Chen, C.; Xue, F.-S. Photoperiodism of diapause induction and diapause termination in Ostrinia furnacalis. Entomol. Exp. Appl. 2014, 153, 34–46. [Google Scholar] [CrossRef]
- Oltean, I.; Hulujan, I.; Varga, M.; Totos, Ş.; Florian, T. Cydalima Perspectalis Walker (Lepidoptera, Crambidae) a New Dangerous Pest Report on Buxus Sempervirens in Cluj Area. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Agric. 2017, 74, 26. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-P.; Yang, Q.-S.; Zhou, X.-M.; Xu, S.; Lei, C.-L. Effects of photoperiod and temperature on diapause induction and termination in the swallowtail, Sericinus montelus. Physiol. Entomol. 2009, 34, 158–162. [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Shiryaeva, N.V.; Shurov, V.I. The box tree moth—A new invasive pest in the Caucasian Forests. Plant Health Res. Pract. 2014, 1, 32–39. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | 5 | 10 | 15 | 20 | ||||
|---|---|---|---|---|---|---|---|---|
| Larval Instar | L3 | L4 | L3 | L4 | L3 | L4 | L3 | L4 |
| Photoperiod (L:D) | ||||||||
| 8:16 | 20 | 34 | 20 | 34 | 20 | 34 | 20 | 34 |
| 10:14 | 20 | 34 | 20 | 34 | 20 | 34 | 20 | 34 |
| 12:12 | 20 | 34 | 20 | 34 | 20 | 34 | 20 | 34 |
| 14:10 | 20 | 34 | 20 | 34 | 20 | 34 | 20 | 34 |
| 16:8 | 20 | 34 | 20 | 34 | 20 | 34 | 20 | 34 |
| Experimental Condition | Set Temperature (°C) | Actual Mean Temperature ± SD (°C) |
|---|---|---|
| Winter | 5 | 6.0 ± 1.0 |
| Temperature 5 (T5) | 5 | 8.6 ± 0.6 |
| Temperature 10 (T10) | 10 | 12.2 ± 0.5 |
| Temperature 15 (T15) | 15 | 16.6 ± 0.5 |
| Temperature 20 (T20) | 20 | 21.4 ± 0.7 |
| Factor | χ² | Df | p-Value |
|---|---|---|---|
| Temperature | 351.0 | 3 | *** |
| Larval instar | 21.1 | 1 | **** |
| Photoperiod | 39.2 | 4 | **** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poitou, L.; Bras, A.; Pineau, P.; Lorme, P.; Roques, A.; Rousselet, J.; Auger-Rozenberg, M.-A.; Laparie, M. Diapause Regulation in Newly Invaded Environments: Termination Timing Allows Matching Novel Climatic Constraints in the Box Tree Moth, Cydalima perspectalis (Lepidoptera: Crambidae). Insects 2020, 11, 629. https://doi.org/10.3390/insects11090629
Poitou L, Bras A, Pineau P, Lorme P, Roques A, Rousselet J, Auger-Rozenberg M-A, Laparie M. Diapause Regulation in Newly Invaded Environments: Termination Timing Allows Matching Novel Climatic Constraints in the Box Tree Moth, Cydalima perspectalis (Lepidoptera: Crambidae). Insects. 2020; 11(9):629. https://doi.org/10.3390/insects11090629
Chicago/Turabian StylePoitou, Laura, Audrey Bras, Patrick Pineau, Philippe Lorme, Alain Roques, Jérôme Rousselet, Marie-Anne Auger-Rozenberg, and Mathieu Laparie. 2020. "Diapause Regulation in Newly Invaded Environments: Termination Timing Allows Matching Novel Climatic Constraints in the Box Tree Moth, Cydalima perspectalis (Lepidoptera: Crambidae)" Insects 11, no. 9: 629. https://doi.org/10.3390/insects11090629