Population Dynamics of Drosophila suzukii in Coastal and Mainland Sweet Cherry Orchards of Greece

 ,
, 


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sites and Trap Allocation
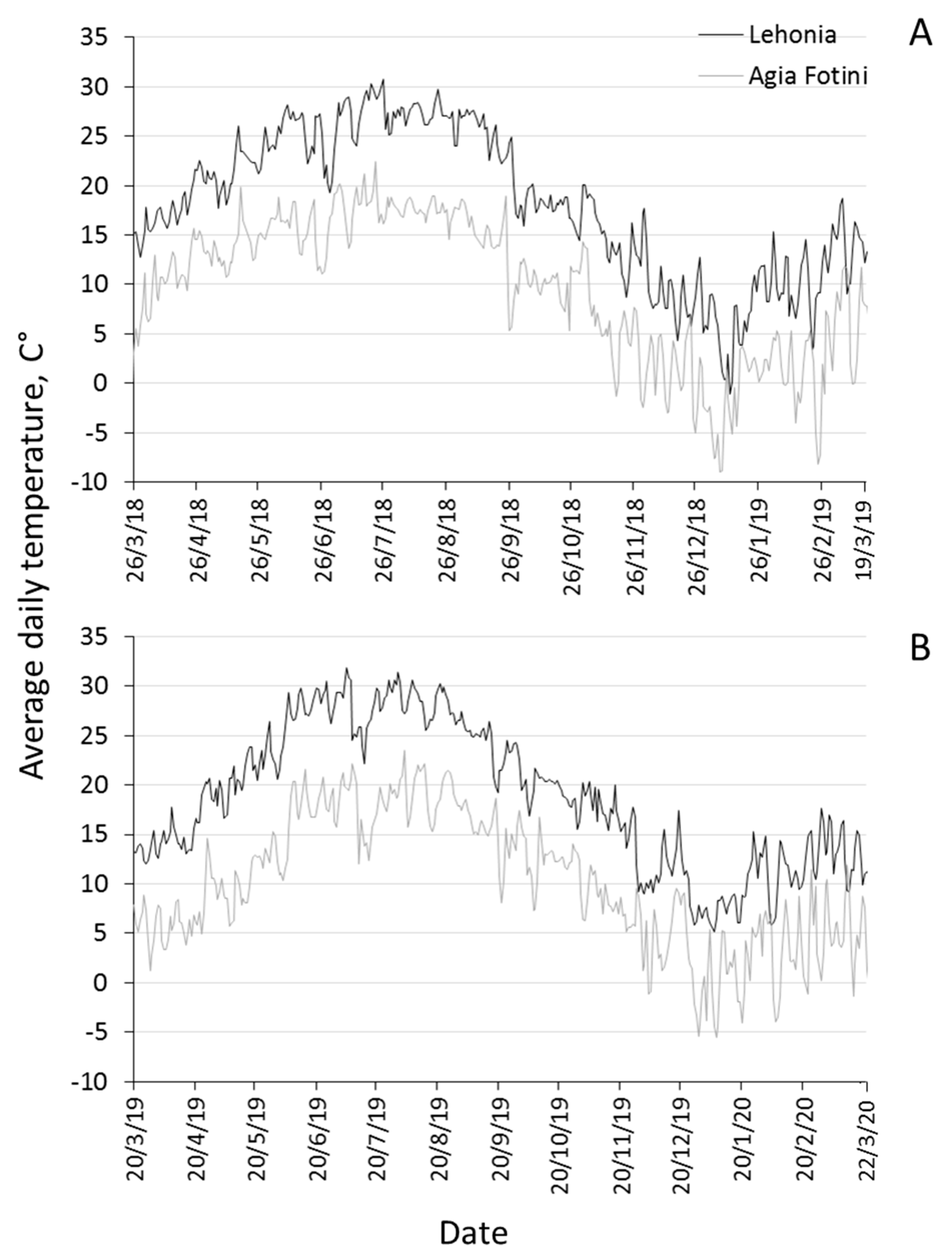
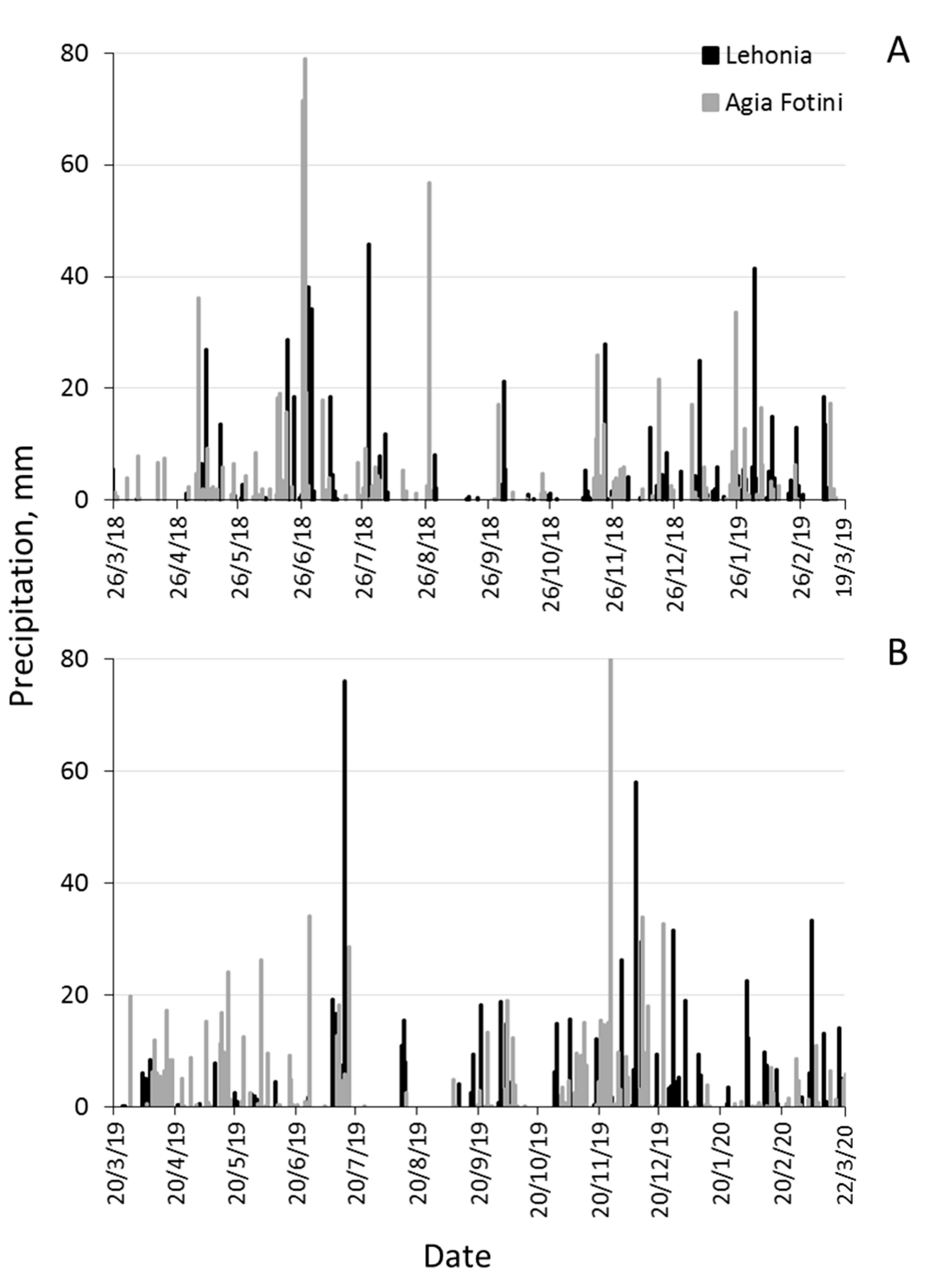
2.2. Climatic Data
2.3. Sweet-Cherry Fruit Sampling
2.4. Statistical Analysis
3. Results
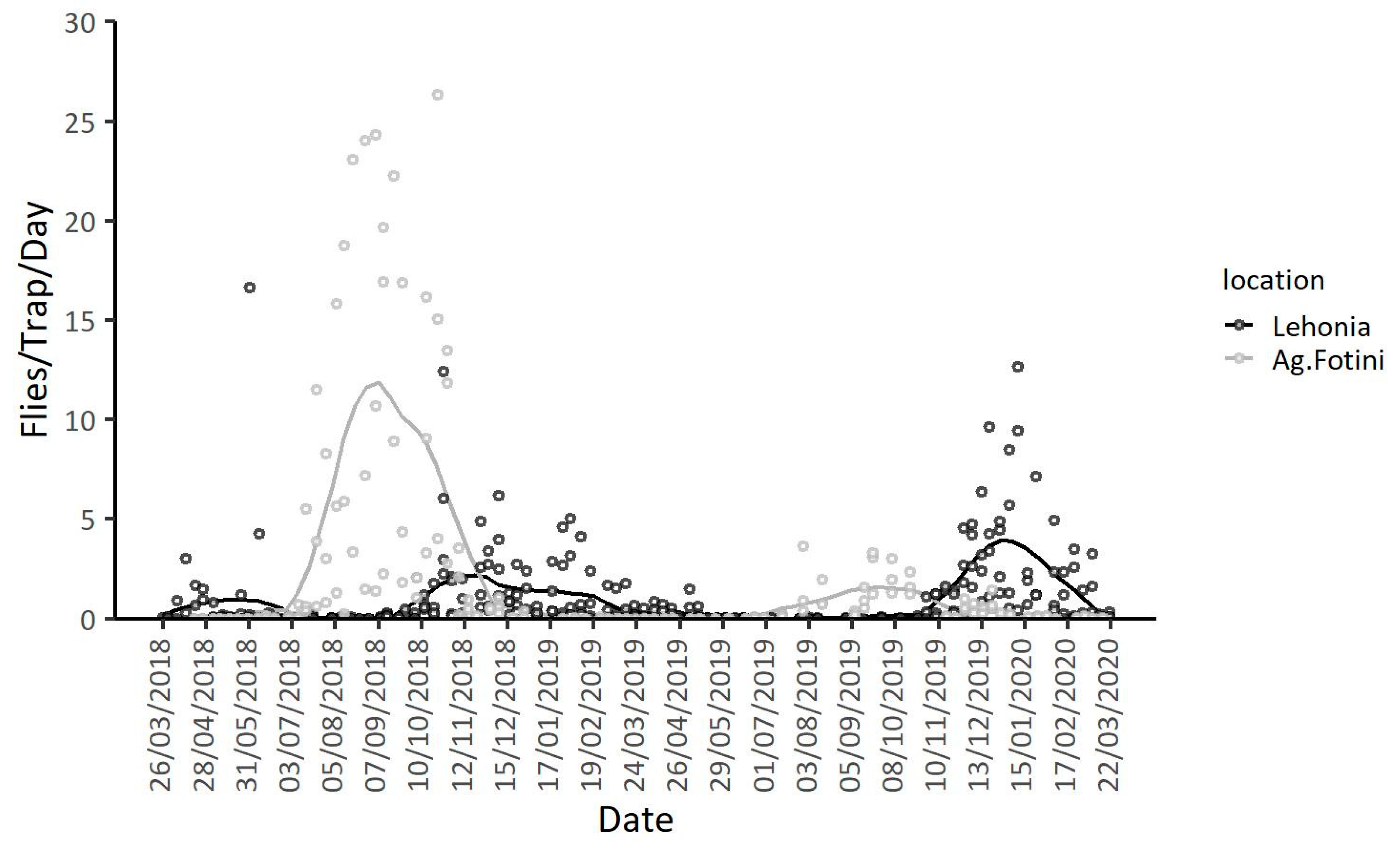
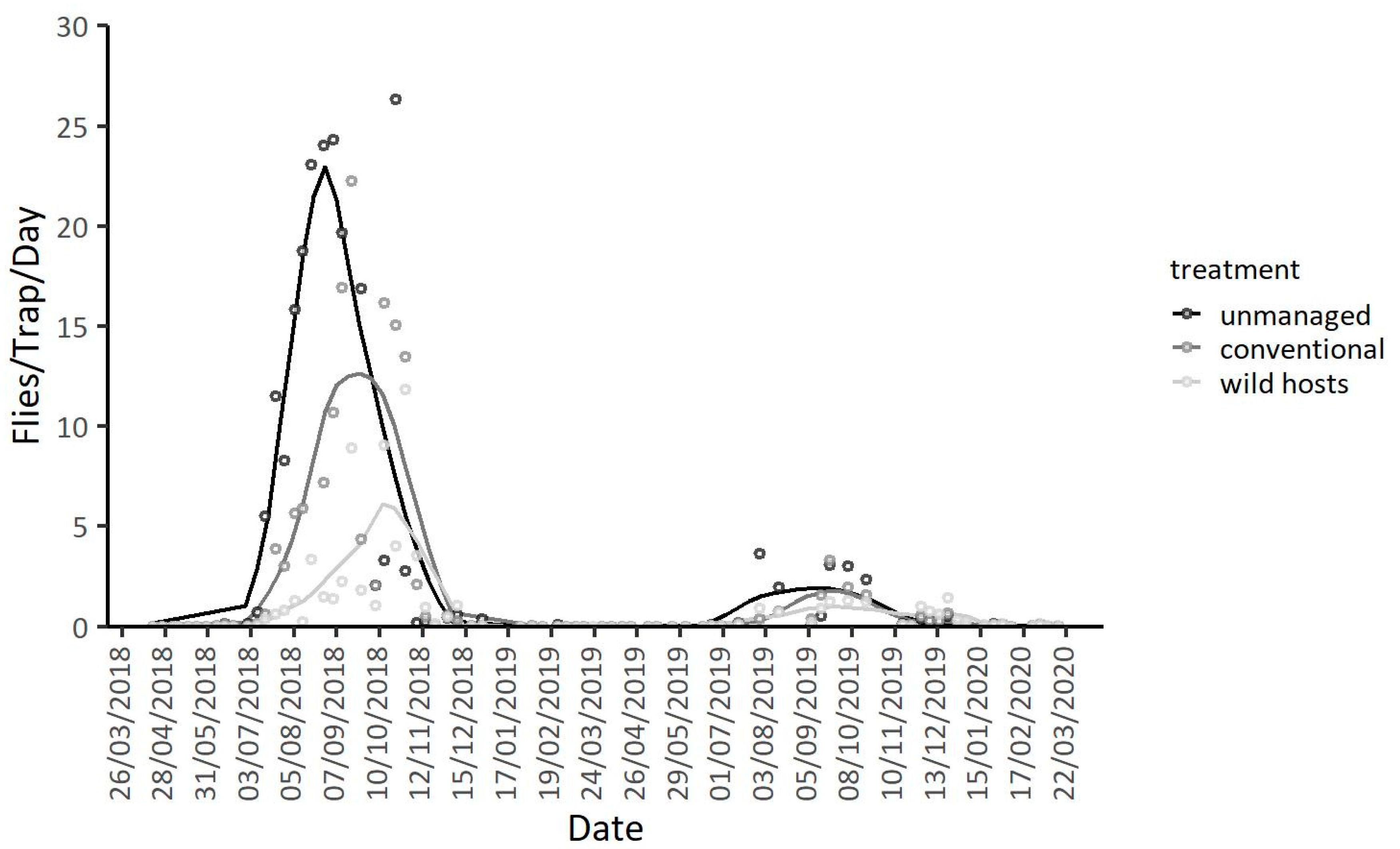
3.1. Population Dynamics in the Two Geographic Locations
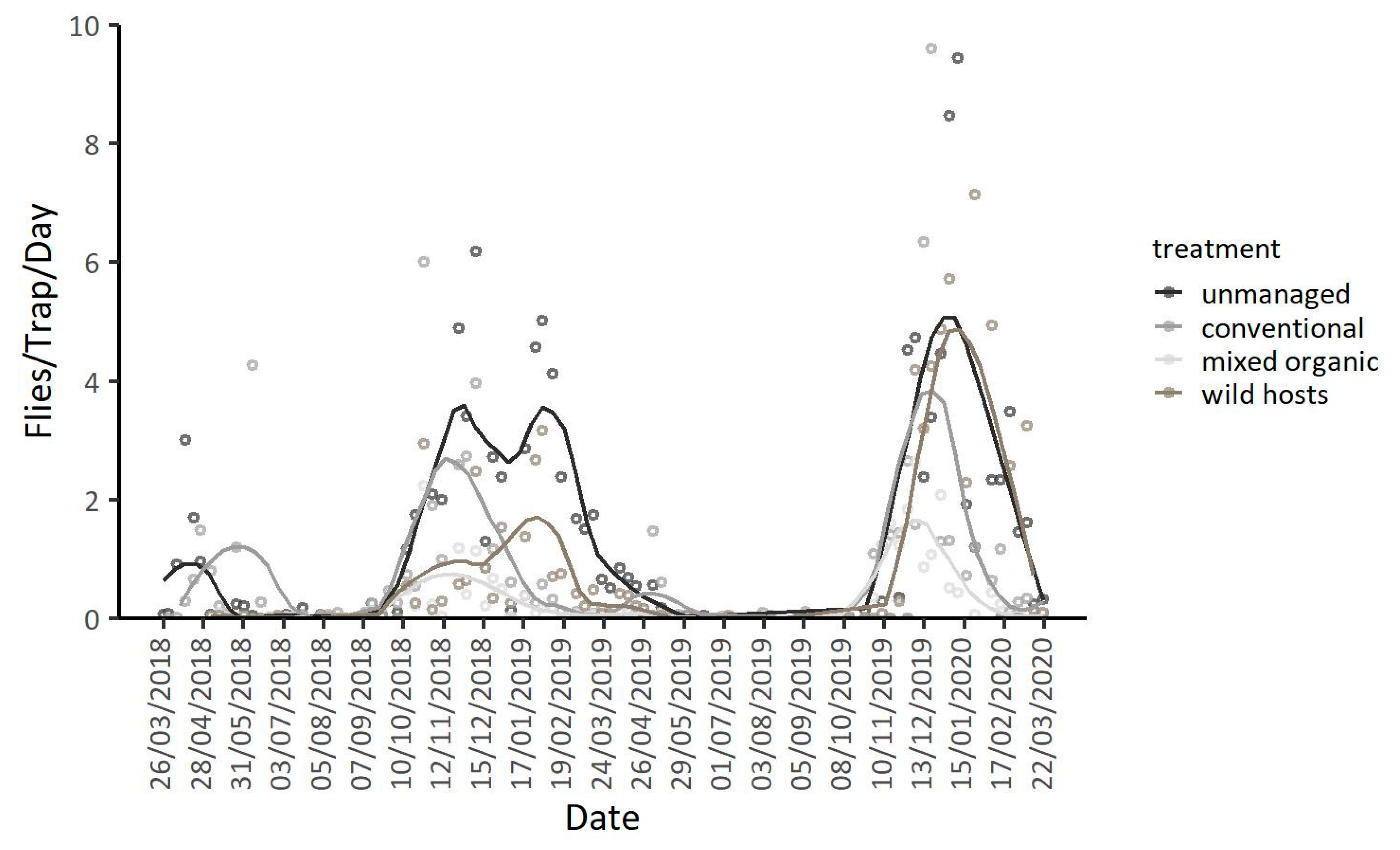
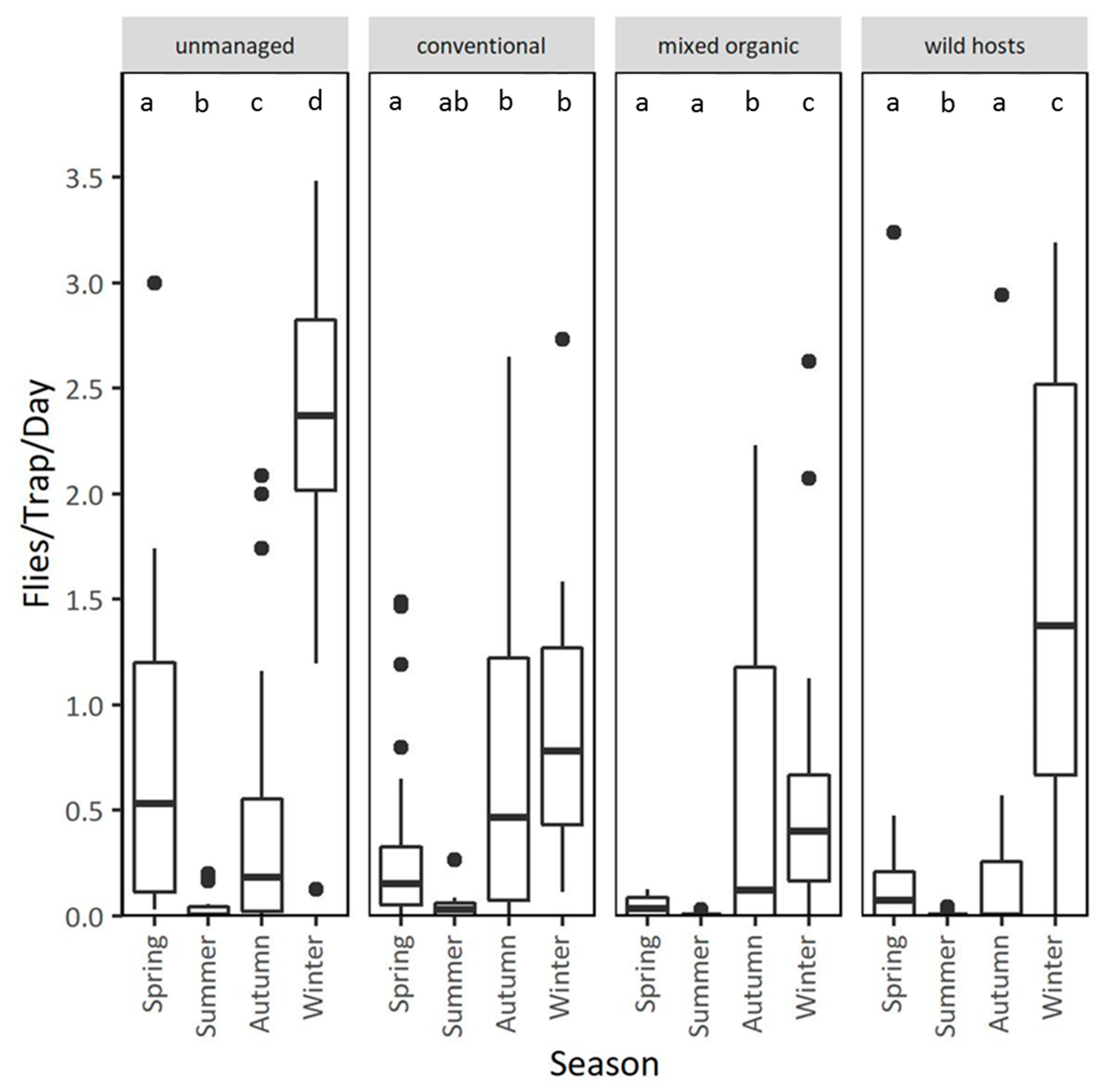
3.2. Population Dynamics in Different Types of Orchards at the Coastal Area Lehonia
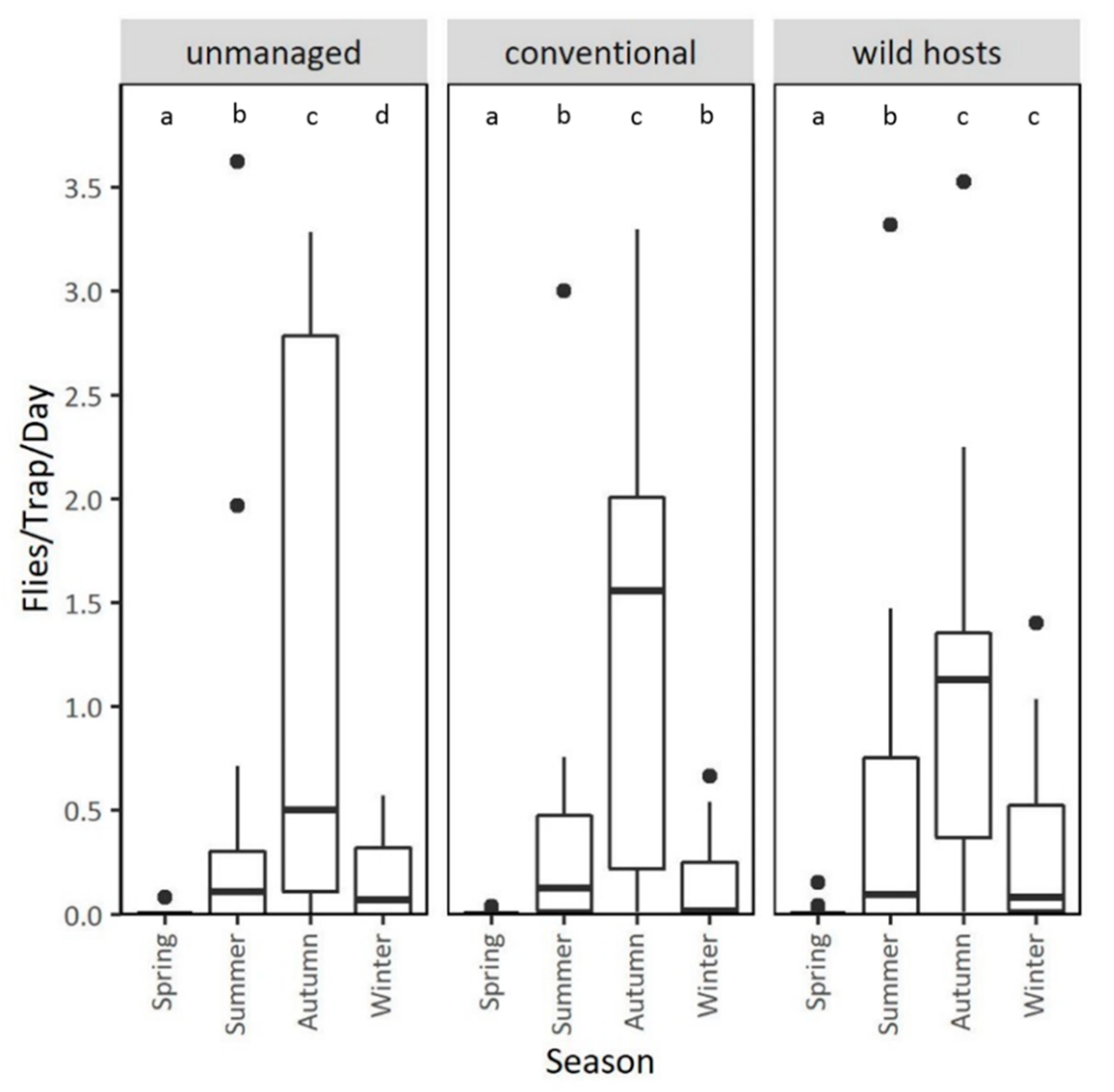
3.3. Population Dynamics in Different Types of Orchards at the Mainland Area Agia Fotini
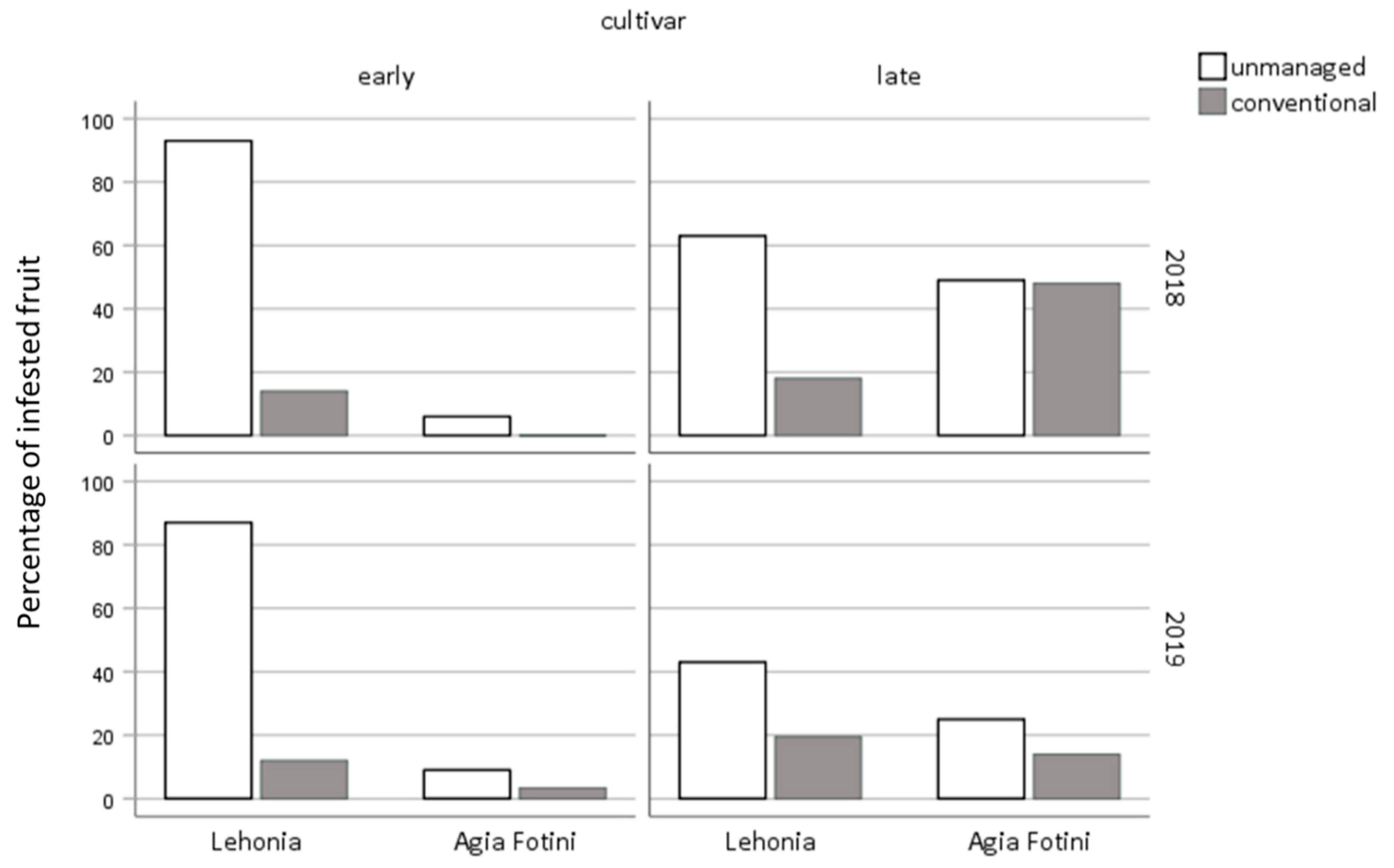
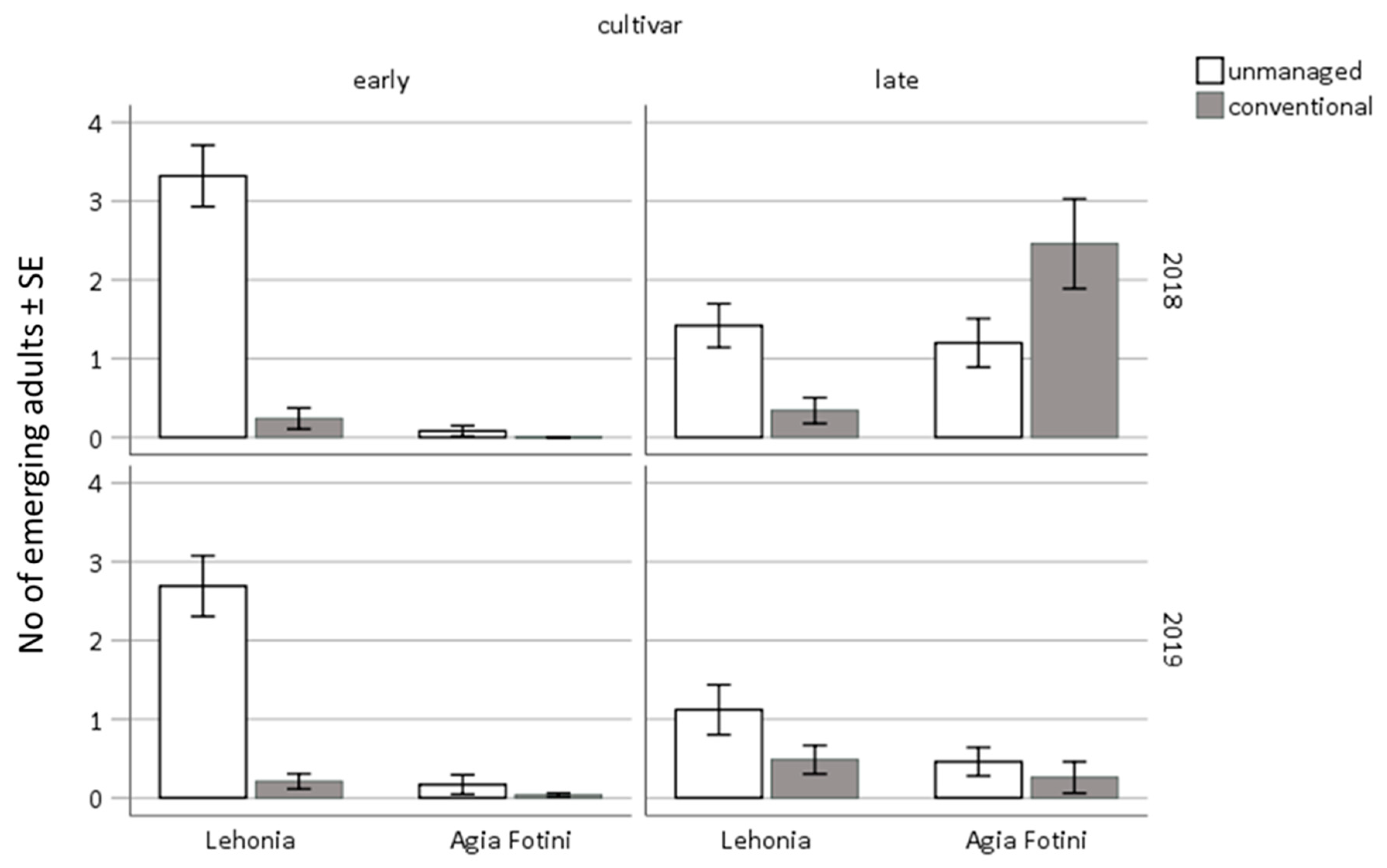
3.4. Sweet-Cherry Infestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. B. Insectol. 2012, 65, 149–160. [Google Scholar]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Arnó, J.; Solà, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Timmeren, S.V.; Bruck, D.J. Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef]
- Barata, A.; Santos, S.C.; Malfeito-Ferreira, M.; Loureiro, V. New insights into the ecological interaction between grape berry microorganisms and Drosophila flies during the development of sour rot. Microb. Ecol. 2012, 64, 416–430. [Google Scholar] [CrossRef]
- Hamby, K.A.; Hernández, A.; Boundy-Mills, K.; Zalom, F.G. Associations of yeasts with Spotted-Wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in cherries and raspberries. Appl. Environ. Microb. 2012, 78, 4869–4873. [Google Scholar] [CrossRef] [Green Version]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.C.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S. Integrating temperature-dependent life table data into a matrix projection model for Drosophila suzukii population estimation. PLoS ONE 2014, 9, 106909. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- dos Santos, L.A.; Mendes, M.F.; Krüger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R. Global potential distribution of Drosophila suzukii (Diptera; Drosophilidae). PLoS ONE 2017, 12, 174318. [Google Scholar] [CrossRef] [Green Version]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Dos Santos, R.S.S.; Garcia, F.R.M.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila suzukii in Southern Neotropical Region: Current status and future perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef]
- Cini, A.; Anfora, G.; Escudero-Colomar, L.A.; Grassi, A.; Santosuosso, U.; Seljak, G.; Papini, A. Tracking the invasion of the alien fruit pest Drosophila suzukii in Europe. J. Pest Sci. 2014, 87, 559–566. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- EPPO Global Database. Available online: https://gd.eppo.int/ (accessed on 20 April 2020).
- Papachristos, D.P.; Matakoulis, C.; Papadopoulos, N.T.; Lagkouranis, A.; Zarpas, K.D.; Milonas, P.G. First record of the harmful pest Drosophila suzukii (Diptera: Drosophilidae) in Greece. In Proceedings of the 15th Panhellenic Entomological Congress, Kavala, Greece, 22–25 October 2013; Andreadis, S.S., Pappas, M.L., Eds.; Hellenic Entomological Society: Kavala, Greece, 2013. [Google Scholar]
- Máca, J. Drosophila suzukii (Matsumura) found on the Greek island Crete. Drosoph. Inf. Serv. 2014, 97, 28–29. [Google Scholar]
- Garantonakis, N.; Varikou, K.; Birouraki, A. Spread of Drosophila suzukii in orchards of Crete. Entomol. Gen. 2016, 36, 63–70. [Google Scholar] [CrossRef]
- Fytrou, N.; Kapantaidaki, D.E.; Antonatos, S.; Zarpas, K.; Papadopoulos, N.T.; Papachristos, D.P. Mitochondrial DNA sequence diversity of Drosophila suzukii in Greece. In Proceedings of 17th Panhellenic Entomological Congress, Athens, Greece, 19–22 September 2017; Litra, I., Perdikis, D., Eds.; Hellenic Entomological Society: Athens, Greece, 2017. [Google Scholar]
- FAO. Available online: http://www.fao.org/faostat/en/#data/ (accessed on 20 April 2020).
- Mitsui, H.; Beppu, K.; Kimura, M.T. Seasonal life cycles and resource uses of flower-and fruit-feeding drosophilid flies (Diptera: Drosophilidae) in central Japan. Entomol. Sci. 2010, 13, 60–67. [Google Scholar] [CrossRef]
- Wang, X.G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest. Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Beers, E.H.; Van Steenwyk, R.A.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Burrack, H.J.; Barrantes, L.D.; Beers, E.H.; Dreves, A.J.; Hamby, K.A.; Haviland, D.R.; Isaacs, R.; Richardson, T.A.; Shearer, P.W.; et al. Evaluation of monitoring traps for Drosophila suzukii (Diptera: Drosophilidae) in North America. J. Econ. Entomol. 2012, 105, 1350–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briem, F.; Dominic, A.R.; Golla, B.; Hoffmann, C.; Englert, C.; Herz, A.; Vogt, H. Explorative data analysis of Drosophila suzukii trap catches from a seven-year monitoring program in Southwest Germany. Insects 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R Package geepack for Generalized Estimating Equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Fine, J.P. Estimating equations for association structures. Stat. Med. 2004, 23, 859–880. [Google Scholar] [CrossRef]
- Yan, J. Geepack: Yet another package for Generalized Estimating Equations”. R-News 2002, 2, 12–14. [Google Scholar]
- Wollmann, J.; Schlesener, D.C.H.; Ferreira, M.S.; Kruger, A.P.; Bernardi, D.; Garcia, J.A.B.; Nunes, A.M.; Garcia, M.S.; Garcia, F.R.M. Population dynamics of Drosophila suzukii (Diptera: Drosophilidae) in berry crops in Southern Brazil. Neotrop. Entomol. 2019, 48, 699–705. [Google Scholar] [CrossRef]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef]
- Kanzawa, T. Studies on Drosophila suzukii Mats. Rev. Appl. Entomol. 1939, 29, 49. [Google Scholar]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal tolerances of the spotted-wing Drosophila Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinjo, H.; Kunimi, Y.; Nakai, M. Effects of temperature on the reproduction and development of Drosophila suzukii (Diptera: Drosophilidae). Appl. Entomol. Zool. 2014, 49, 297–304. [Google Scholar] [CrossRef]
- Mazzetto, F.; Pansa, M.G.; Ingegno, B.L.; Tavella, L.; Alma, A. Monitoring of the exotic fly Drosophila suzukii in stone, pome and soft fruit orchards in NW Italy. J. Asia-Pac. Entomol. 2015, 18, 321–329. [Google Scholar] [CrossRef]
- Harris, D.W.; Hamby, K.A.; Wilson, H.E.; Zalom, F.G. Seasonal monitoring of Drosophila suzukii (Diptera: Drosophilidae) in a mixed fruit production system. J. Asia-Pac. Entomol. 2014, 17, 857–864. [Google Scholar] [CrossRef]
- Kacar, G.; Wang, X.G.; Stewart, T.J.; Daane, K.M. Overwintering survival of Drosophila suzukii (Diptera: Drosophilidae) and the effect of food on adult survival in California’s San Joaquin Valley. Environ. Entomol. 2016, 45, 763–771. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest. Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Pelton, E.; Gratton, C.; Isaacs, R.; Van Timmeren, S.; Blanton, A.; Guédot, C. Earlier activity of Drosophila suzukii in high woodland landscapes but relative abundance is unaffected. J. Pest. Sci. 2016, 89, 725–733. [Google Scholar] [CrossRef]
- Kinjo, H.; Kunimi, Y.; Ban, T.; Nakai, M. Oviposition efficacy of Drosophila suzukii (Diptera: Drosophilidae) on different cultivars of blueberry. J. Econ. Entomol. 2013, 106, 1767–1771. [Google Scholar] [CrossRef] [Green Version]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Burrack, H.J.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Orchards | Lat. (North) | Long. (East) | Treatment | Number of Traps | Date of Trap Establishment |
|---|---|---|---|---|---|---|
| Lehonia 1 | 1 | 39°19′28.12″ | 23°2′57.53″ | Unmanaged cherries | 10 | 16 March 2018 |
| Wild hosts | 2 | 26 April 2018 | ||||
| 2 | 39°19′27.99″ | 23°2′20.85″ | Cherries | 10 | 26 March 2018 | |
| Wild hosts | 2 | 26 April 2018 | ||||
| 3 | 39°19′37.93″ | 23°1′35.19″ | Cherries | 5 | 30 March 2018 | |
| 4 | 39°19′9.21″ | 23°1′34.53″ | Stone fruit | 2 | 26 April 2018 | |
| Pome fruit | 3 | 26 April 2018 | ||||
| Wild host | 1 | 26 April 2018 | ||||
| Agia Fotini 2 | 1 | 40°43′8.46″ | 22°0′20.63″ | Conventional cherries | 10 | 11 April 2018 |
| Unmanaged cherries | 2 | 11 April 2018 | ||||
| Wild hosts | 2 | 11 April 2018 | ||||
| 2 | 40°42′0.10″ | 22°0′14.80″ | Conventional cherries | 10 | 11 April 2018 | |
| Wild hosts | 2 | 11 April 2018 | ||||
| 3 | 40°42′7.07″ | 22°0′25.64″ | Conventional cherries | 3 | 31 March 2019 | |
| Wild hosts | 2 | 31 March 2019 | ||||
| 4 | 40°41′30.56″ | 21°59′55.08″ | Conventional cherries | 3 | 31 March 2019 | |
| Wild hosts | 2 | 31 March 2019 | ||||
| 5 | 40°41′42.51″ | 22°0′1.00″ | Conventional cherries | 3 | 31 March 2019 | |
| Wild hosts | 2 | 31 March 2019 | ||||
| 6 | 40°42′55.97″ | 22°0′20.79″ | Conventional cherries | 3 | 31 March 2019 | |
| Wild hosts | 2 | 31 March 2019 |
| Location | Year | Ripening Period | Sweet-Cherry Cultivar | Treatment | Cherry Harvest/Sampling Dates | Number of Cherries |
|---|---|---|---|---|---|---|
| Lehonia | 2018 | Early | Sweet early | Conventional | 5 May | 100 |
| Early lory | Unmanaged | 5 May | 100 | |||
| Late | Precoce bernard | Conventional | 14–20 May | 100 | ||
| Precoce bernard | Unmanaged | 14–20 May | 100 | |||
| 2019 | Early | Sweet early | Conventional | 11 May | 200 | |
| Early lory | Unmanaged | 11 May | 100 | |||
| Late | Precoce bernard | Conventional | 1 June | 200 | ||
| Precoce bernard | Unmanaged | 28 May | 100 | |||
| Agia Fotini | 2018 | Early | Prime giant | Conventional | 6 June | 100 |
| Skeena | Unmanaged | 24 June | 100 | |||
| Late | Gesmestorfer | Conventional | 8 July | 100 | ||
| Gesmestorfer | Unmanaged | 16 July | 100 | |||
| 2019 | Early | Prime giant, Skeena | Conventional | 16 & 26 June | 300 | |
| Prime giant | Unmanaged | 16 June | 100 | |||
| Late | Gesmestorfer | Conventional | 18 July | 100 | ||
| Gesmestorfer | Unmanaged | 18 July | 100 |
| Factor | IRR (95% CI) | p Value |
|---|---|---|
| Intercept | 0.64 (0.26, 1.53) | 0.313 |
| Location (ref: Lehonia) | 3.28 (1.84, 5.85) | <0.001 |
| Year (ref: first year) | 0.24 (0.18, 0.33) | <0.001 |
| Season (ref: spring) | ||
| summer | 5.65 (2.28, 14.00) | <0.001 |
| autumn | 9.43 (4.54, 19.60) | <0.001 |
| winter | 5.75 (2.77, 11.90) | <0.001 |
| Mean daily temperature | 1.06 (1.03, 1.09) | <0.001 |
| Factor | Lehonia | Agia Fotini | ||
|---|---|---|---|---|
| IRR (95% CI) | p Value | IRR (95% CI) | p Value | |
| Intercept | 25.30 (16.10, 39.90) | <0.001 | 0.17 (0.11, 0.28) | <0.001 |
| Treatment (ref: unmanaged) | ||||
| conventional | 0.39 (0.18, 0.87) | 0.021 | 0.62 (0.40, 0.97) | 0.034 |
| wild hosts | 0.44 (0.13, 1.52) | 0.196 | 1.38 (0.63, 3.02) | 0.427 |
| mixed organic | 0.07 (0.05, 0.11) | <0.001 | - | - |
| Year (ref: first year) | 0.99 (0.75, 1.33) | 0.992 | 0.17 (0.14, 0.20) | <0.001 |
| Season (ref: spring) | ||||
| summer | 0.17 (0.12, 0.23) | <0.001 | 60.70 (48.00, 76.70) | <0.001 |
| autumn | 3.17 (2.08, 4.84) | <0.001 | 115.00 (92.00, 143.00) | <0.001 |
| winter | 1.97 (1.50, 2.57) | <0.001 | 18.50 (13.10, 26.10) | <0.001 |
| Mean Temperature | 0.91 (0.90, 0.93) | <0.001 | 1.12 (1.09, 1.14) | <0.001 |
| Treatment * Season 1 | ||||
| conventional * summer | 82.00 (16.90, 398.00) | <0.001 | 0.73 (0.56, 0.95) | 0.017 |
| wild hosts * summer | 0.69 (0.18, 2.67) | 0.588 | 0.20 (0.10, 0.41) | <0.001 |
| mixed organic * summer | 2.25 (1.29, 3.91) | <0.001 | - | - |
| conventional * autumn | 1.68 (0.81, 3.48) | 0.168 | 1.42 (1.06, 1.91) | 0.020 |
| wild hosts * autumn | 0.40 (0.16, 0.99) | 0.048 | 0.41 (0.17, 0.98) | 0.045 |
| mixed organic * autumn | 3.65 (2.13, 6.25) | <0.001 | - | - |
| conventional * winter | 1.30 (0.62, 2.70) | 0.490 | 1.69 (1.14, 2.52) | 0.009 |
| wild hosts * winter | 1.66 (0.98, 2.82) | 0.059 | 1.79 (1.10, 2.93) | 0.019 |
| mixed organic * winter | 2.42 (1.64, 3.59) | <0.001 | - | - |
| Dependent Variable | Factor | OR (95% CI) | p Value |
|---|---|---|---|
| Cherry infestation | Intercept | 0.11 (0.08, 0.16) | <0.001 |
| Location (ref: Ag. Fotini) | 1.60 (1.07, 2.40) | 0.023 | |
| Year (ref: second year) | 2.70 (1.80, 4.05) | <0.001 | |
| Treatment (ref: unmanaged) | 6.08 (4.79, 7.73) | <0.001 | |
| Cherry cultivar (ref: late) | 0.09 (0.06, 0.15) | <0.001 | |
| Location (Lehonia) * Year (first) 1 | 0.56 (0.34, 0.93) | 0.024 | |
| Location (Lehonia) * Cherry cultivar (early) | 19.61 (11.26, 34.15) | <0.001 | |
| Dependent Variable | Factor | IRR (95% CI) | p value |
| Emerging adults | Intercept | 2.53 (2.23, 2.87) | <0.001 |
| Location (ref: Ag. Fotini) | 0.94 (0.83, 1.06) | 0.324 | |
| Year (ref: second year) | 1.27 (1.15, 1.41) | <0.001 | |
| Treatment (ref: unmanaged) | 0.94 (0.84, 1.06) | 0.309 | |
| Cherry cultivar (ref: late) | 1.19 (1.00, 1.25) | 0.050 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papanastasiou, S.A.; Rodovitis, V.G.; Bataka, E.P.; Verykouki, E.; Papadopoulos, N.T. Population Dynamics of Drosophila suzukii in Coastal and Mainland Sweet Cherry Orchards of Greece. Insects 2020, 11, 621. https://doi.org/10.3390/insects11090621
Papanastasiou SA, Rodovitis VG, Bataka EP, Verykouki E, Papadopoulos NT. Population Dynamics of Drosophila suzukii in Coastal and Mainland Sweet Cherry Orchards of Greece. Insects. 2020; 11(9):621. https://doi.org/10.3390/insects11090621
Chicago/Turabian StylePapanastasiou, Stella A., Vasilis G. Rodovitis, Evmorfia P. Bataka, Eleni Verykouki, and Nikos T. Papadopoulos. 2020. "Population Dynamics of Drosophila suzukii in Coastal and Mainland Sweet Cherry Orchards of Greece" Insects 11, no. 9: 621. https://doi.org/10.3390/insects11090621