Wing Geometric Morphometrics as a Tool for the Identification of Culex Subgenus Mosquitoes of Culex (Diptera: Culicidae)


 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Sampling and Identification
2.2. Data Acquisition and Morphometrics Analysis
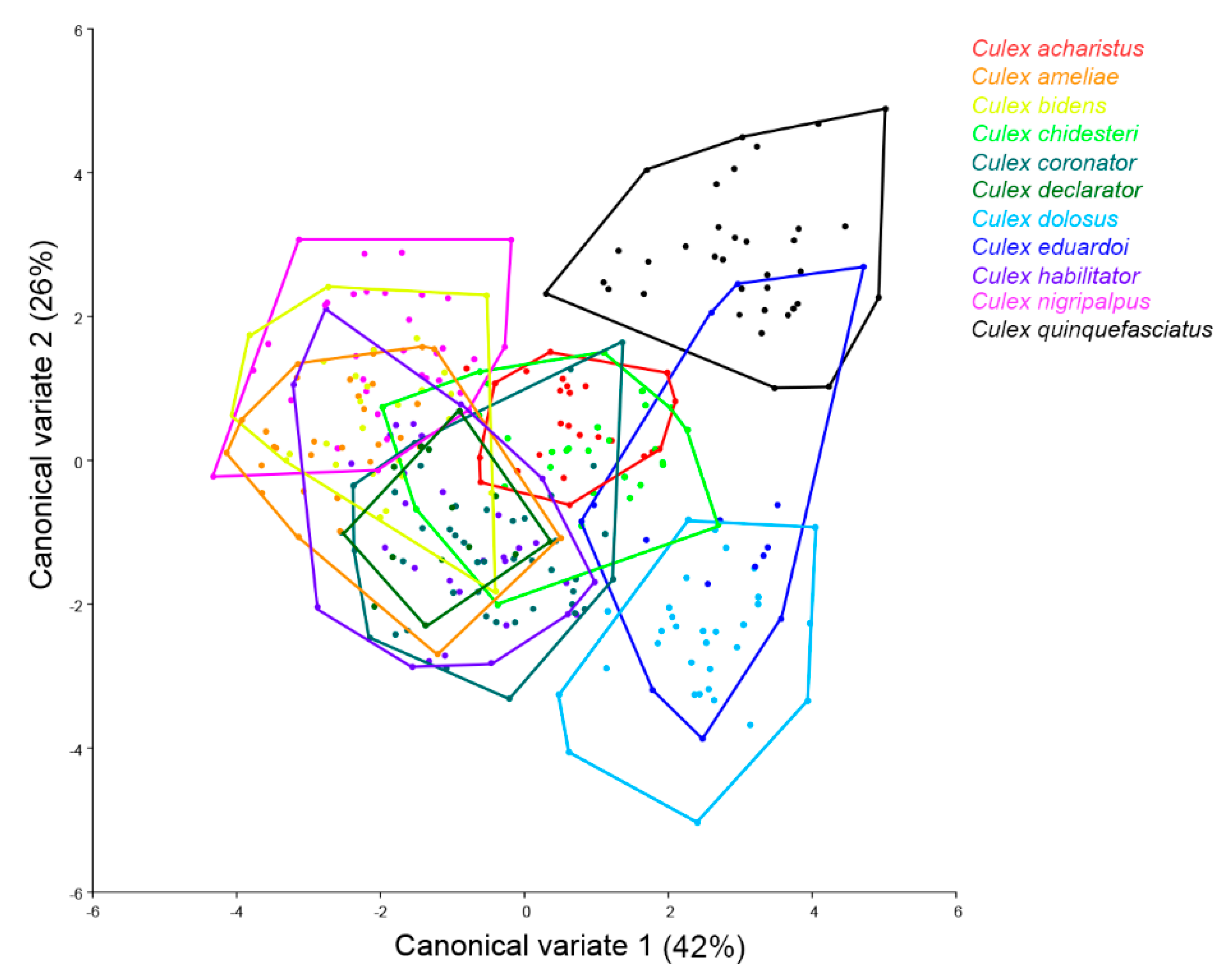
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shapiro-Mendoza, C.K.; Rice, M.E.; Galang, R.R.; Fulton, A.C.; VanMaldeghem, K.; Prado, M.V.; Ellis, E.; Anesi, M.S.; Simeone, R.M.; Petersen, E.E.; et al. Pregnancy Outcomes After Maternal Zika Virus Infection During Pregnancy—U.S. Territories, 1 January 2016–25 April 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Delaney, A.; Mai, C.; Smoots, A.; Cragan, J.; Ellington, S.; Langlois, P.; Breidenbach, R.; Fornoff, J.; Dunn, J.; Yazdy, M.; et al. Population-based surveillance of birth defects potentially related to Zika virus infection—15 States and U.S. Territories, 2016. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital Signs: Trends in reported vectorborne disease cases—United States and Territories, 2004–2016. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Handbook on Integrated Vector Management; WHO: Geneva Switzerland, 2012. [Google Scholar]
- Lizzi, K.M.; Qualls, W.A.; Brown, S.C.; Beier, J.C. Expanding Integrated Vector Management to promote healthy environments. Trends Parasitol. 2014, 30, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbach, R.E. Classification within the cosmopolitan genus Culex (Diptera: Culicidae): The foundation for molecular systematics and phylogenetic research. Acta Trop. 2011, 120, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária no Brasil; Cadernos de Saúde Pública, Editora FIOCRUZ: Rio de Janeiro, Brazil, 1994. [Google Scholar] [CrossRef] [Green Version]
- Darsie, R.F., Jr.; Morris, C.D. Keys to the Adult Females and Fourth Instar Larvae of the Mosquitoes of Florida (Diptera, Culicidae), 1st ed.; Technical Bulletin of the Florida Mosquito Control Association; Florida Mosquito Control Association, Inc.: Fort Myers, FL, USA, 2000. [Google Scholar]
- Forattini, O.P. Culicidologia Médica, Vol. 2: Identificação, Biologia, Epidemiologia; Edusp: São Paulo, Brazil, 2002. [Google Scholar]
- Murugan, K.; Vadivalagan, C.; Karthika, P.; Panneerselvam, C.; Paulpandi, M.; Subramaniam, J.; Wei, H.; Aziz, A.T.; Alsalhi, M.S.; Devanesan, S.; et al. DNA barcoding and molecular evolution of mosquito vectors of medical and veterinary importance. Parasitol. Res. 2016, 115, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Calle, D.A.; Quiñones, M.L.; Erazo, H.F.; Jaramillo, N. Morphometric discrimination of females of five species of Anopheles of the subgenus Nyssorhynchus from Southern and Northwest Colombia. Mem. Inst. Oswaldo Cruz 2002, 97, 1191–1195. [Google Scholar] [CrossRef]
- Börstler, J.; Lühken, R.; Rudolf, M.; Steinke, S.; Melaun, C.; Becker, S.; Garms, R.; Krüger, A. The use of morphometric wing characters to discriminate female Culex pipiens and Culex torrentium. J. Vector Ecol. 2014, 39, 204–212. [Google Scholar] [CrossRef]
- Laurito, M.; Almirón, W.R.; Ludueña-Almeida, F.F. Discrimination of four Culex (Culex) species from the Neotropics based on geometric morphometrics. Zoomorphology 2015, 1611, 447–455. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Christe, R.O.; Multini, L.C.; Vidal, P.O.; Wilk-da-Silva, R.; de Carvalho, G.C.; Marrelli, M.T. Morphometric wing characters as a tool for mosquito identification. PLoS ONE 2016, 11, e0161643. [Google Scholar] [CrossRef]
- Lorenz, C.; Almeida, F.; Almeida-Lopes, F.; Louise, C.; Pereira, S.N.; Petersen, V.; Vidal, P.O.; Virginio, F.; Suesdek, L. Geometric morphometrics in mosquitoes: What has been measured? Infect. Genet. Evol. 2017, 54, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Sousa, A.R.; Ceretti, W.; Urbinatti, P.R.; de Carvalho, G.C.; de Paula, M.B.; Fernandes, A.; Matos, M.O.; Orico, L.D.; Araujo, A.B.; Nardi, M.S.; et al. Mosquito fauna in municipal parks of São Paulo City, Brazil: A preliminary survey. J. Am. Mosq. Control Assoc. 2013, 29, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Junior, W.; Wilke, A.B.B.; Marrelli, M.T. Mosquitoes in urban green spaces: Using an island biogeographic approach to identify drivers of species richness and composition. Sci. Rep. 2017, 7, 17826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagas, C.R.F. Plasmodium spp. em aves Silvestres da Fundação Parque Zoológico de São Paulo: Identificação de Espécies por Microscopia e Sequenciamento do Gene Mitocondrial Citocromo b. Ph.D. Thesis, Instituto de Medicina Tropical de São Paulo, Universidade de São Paulo, São Paulo, Brazil, 2016. [Google Scholar] [CrossRef] [Green Version]
- Sudia, W.D.; Chamberlain, R.W. Battery-operated light trap, an improved model. Mosq. News 1962, 22, 126–129. [Google Scholar]
- Forattini, O.P. Entomologia Médica, vol.2: Culicini: Culex, Aedes e Psorophora; Edusp: São Paulo, Brazil, 1965. [Google Scholar]
- Sabattini, M.S.; Avilés, G.; Monath, T.P. Historical, epidemiological and ecological aspects of arboviruses in Argentina: Togaviridae, Alphavirus. In An Overview of Arbovirology in Brazil and Neighbouring Countries; da Rosa, A.P.A.T., Vasconcelos, P.F.C., da Rosa, J.F.S.T., Eds.; Instituto Evandro Chagas: Belem, Brazil, 1998. [Google Scholar]
- Centers for Disease Control and Prevention. CDC, Arbovirus Catalog. 2018. Available online: https://wwwn.cdc.gov/Arbocat/Default.aspx (accessed on 3 March 2020).
- Pisano, M.B.; Ré, V.E.; Díaz, L.A.; Farías, A.; Stein, M.; Sanchez-Seco, M.P.; Tenorio, A.; Almirón, W.R.; Contigiani, M.S. Enzootic activity of pixuna and Rio Negro viruses (Venezuelan equine encephalitis complex) in a neotropical region of Argentina. Vector Borne Zoonotic Dis. 2010, 10, 199–201. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. CDC, Mosquito Species in which West Nile Virus Has Been Detected. 2017. Available online: https://www.cdc.gov/westnile/resources/pdfs/MosquitoSpecies1999-2016.pdf (accessed on 3 March 2020).
- Vasconcelos, P.F.C.; Travassos-da-Rosa, J.F.S.; Travassos-da-Rosa, A.P.A.; Dégallier, N.; Pinheiro, F.P.; Sá-Filho, G.C. Epidemiologia das encefalites por arbovírus na Amazônia brasileira. Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef]
- Labarthe, N.; Serrão, M.L.; Melo, F.Y.; De Oliveira, S.J.; Lourenço-de Oliveira, R. Potential vectors of Dirofilaria immitis (Leidy, 1856) in Itacoatiara, Oceanic Region of Niterói municipality, State of Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz 1998, 93, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Barrera, R.; MacKay, A.; Amador, M.; Vasquez, J.; Smith, J.; Díaz, A.; Acevedo, V.; Cabán, B.; Hunsperger, E.A.; Muñoz-Jordán, J.L. Mosquito vectors of West Nile virus during an epizootic outbreak in Puerto Rico. J. Med. Entomol. 2010, 47, 1185–1195. [Google Scholar] [CrossRef]
- Mackay, I.M.; Arden, K.E. Mayaro virus: A forest virus primed for a trip to the city? Microbes Infect. 2016, 18, 724–734. [Google Scholar] [CrossRef]
- Rohlf, F.J. The tps series of software. Hystrix 2015, 26, 1–4. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Harbach, R.E.; Kitching, I.J.; Culverwell, C.L.; Dubois, J.; Linton, Y.M. Phylogeny of mosquitoes of tribe culicini (Diptera: Culicidae) based on morphological diversity. Zool. Scr. 2012, 41, 499–514. [Google Scholar] [CrossRef]
- Suesdek, L. Microevolution of medically important mosquitoes—A review. Acta Trop. 2019, 191, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Morais, S.A.; Moratore, C.; Suesdek, L.; Marrelli, M.T. Genetic-morphometric variation in Culex quinquefasciatus from Brazil and La Plata, Argentina. Mem. Inst. Oswaldo Cruz 2010, 105, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Wilke, A.B.B.; Vidal, P.O.; Suesdek, L.; Marrelli, M.T. Population genetics of neotropical Culex quinquefasciatus (Diptera: Culicidae). Parasites Vectors 2014, 7, 468. [Google Scholar]
- Vidal, P.O.; Suesdek, L. Comparison of wing geometry data and genetic data for assessing the population structure of Aedes aegypti. Infect. Genet. Evol. 2012, 12, 591–596. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Apiwathnasorn, C.; Ruangsittichai, J.; Sriwichai, P.; Attrapadung, S.; Samung, Y.; Dujardin, J.P. DNA barcoding and wing morphometrics to distinguish three Aedes vectors in Thailand. Acta Trop. 2016, 159, 1–10. [Google Scholar] [CrossRef]
- Silva, S.O.F.; Fuente, A.L.C.; Mello, C.F.; Alencar, J. Morphological differentiation between seven Brazilian populations of Haemagogus capricornii and Hg. janthinomys (Diptera: Culicidae) using geometric morphometry of the wings. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180106. [Google Scholar] [CrossRef] [Green Version]
- Vidal, P.O.; Peruzin, M.C.; Suesdek, L. Wing diagnostic characters for Culex quinquefasciatus and Culex nigripalpus (Diptera, Culicidae). Rev. Bras. Entomol. 2011, 55, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, C.; Marques, T.C.; Sallum, M.A.M.; Suesdek, L. Morphometrical diagnosis of the malaria vectors Anopheles cruzii, An. homunculus and An. bellator. Parasites Vectors 2012, 5, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo, O.N.; Dujardin, J.P.; Calle-Londono, D.; Fonseca-González, I. Geometric morphometrics for the taxonomy of 11 species of Anopheles (Nyssorhynchus) mosquitoes. Med. Vet. Entomol. 2015, 29, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Guinn, M.L.O.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes for West Nile Virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Zinser, M.; Ramberg, F.; Willott, E. Culex quinquefasciatus (Diptera: Culicidae) as a potential West Nile virus vector in Tucson, Arizona: Blood meal analysis indicates feeding on both humans and birds. J. Insect Sci. 2004, 4, 1–3. [Google Scholar] [CrossRef]
- Vitek, C.J.; Richards, S.L.; Mores, C.N.; Day, J.F.; Lord, C.C. Arbovirus transmission by Culex nigripalpus in Florida, 2005. J. Med. Entomol. 2008, 45, 483–493. [Google Scholar] [CrossRef]
- Erickson, S.M.; Xi, Z.; Mayhew, G.F.; Ramirez, J.L.; Aliota, M.T.; Christensen, B.M.; Dimopoulos, G. Mosquito infection responses to developing filarial worms. PLoS Negl. Trop. Dis. 2009, 3, e529. [Google Scholar] [CrossRef] [PubMed]
- Ariani, C.V.; Juneja, P.; Smith, S.; Tinsley, M.C.; Jiggins, F.M. Vector competence of Aedes aegypti mosquitoes for filarial nematodes is affected by age and nutrient limitation. Exp. Gerontol. 2015, 61, 47–53. [Google Scholar] [CrossRef]
- Samy, A.M.; Elaagip, A.H.; Kenawy, M.A.; Ayres, C.F.J.; Peterson, A.T.; Soliman, D.E. Climate change influences on the global potential distribution of the mosquito Culex quinquefasciatus, vector of West Nile virus and lymphatic filariasis. PLoS ONE 2016, 11, e0163863. [Google Scholar] [CrossRef]
- Kramer, L.D.; Styer, L.M.; Ebel, G.D. A global perspective on the epidemiology of West Nile Virus. Annu. Rev. Entomol. 2008, 53, 61–81. [Google Scholar] [CrossRef] [Green Version]
- Cano, J.; Rebollo, M.P.; Golding, N.; Pullan, R.L.; Crellen, T.; Soler, A.; Kelly-Hope, L.A.; Lindsay, S.W.; Hay, S.I.; Bockarie, M.J.; et al. The global distribution and transmission limits of lymphatic filariasis: Past and present. Parasites Vectors 2014, 7, 466. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera vectors of avian Haemosporidian parasites: Untangling parasite life cycles and their taxonomy. Biol. Rev. Camb. Philos. Soc. 2012, 87, 928–964. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, C. Avian Malaria Parasites and Other Haemosporidia—Gediminas Valkiunas; CRC Press: Boca Raton, FL, USA, 2004; 932p, ISBN 0-415-30097-5. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Valkiūnas, G.; Guimarães, L.d.O.; Monteiro, E.F.; Guida, F.J.V.; Simões, R.F.; Rodrigues, P.T.; Luna, E.J.d.A.; Kirchgatter, K. Diversity and distribution of avian malaria and related Haemosporidian parasites in captive birds from a Brazilian megalopolis. Malar. J. 2017, 16, 83. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | N * | Collection Site | Collection Year | Epidemiological Importance |
|---|---|---|---|---|
| Culex (Cux.) acharistus Root | 26 | Esquel | 2016/2017 | EEEV [21] |
| Culex (Cux.) ameliae Casal | 30 | FPZSP | 2015 | Unknown |
| Culex (Cux.) bidens Dyar | 27 | FPZSP | 2015 | EEEV [21] |
| Culex (Cux.) chidesteri Dyar | 30 | Municipal Parks of São Paulo City ** | 2010/2011 | Unknown |
| Culex (Cux.) coronator Dyar and Knab | 43 | FPZSP | 2015 | SLEV [22], VEEV [23], WNV [24] |
| Culex (Cux.) declarator Dyar and Knab | 13 | FPZSP | 2015 | SLEV [25], Dirofilaria immitis [26] |
| Culex (Cux.) dolosus (Lynch Arribálzaga) | 32 | Municipal Parks of São Paulo City ** | 2010/2011 | Unknown |
| Culex (Cux.) eduardoi Casal and Garcia | 15 | Municipal Parks of São Paulo City ** | 2010/2011 | Unknown |
| Culex (Cux.) habilitator Dyar and Knab | 32 | FPZSP | 2015 | WNV [27] |
| Culex (Cux.) nigripalpus Theobald | 34 | Municipal Parks of São Paulo City ** | 2010/2011 | EEEV [22], EVEV [22], KEYV [22], ROCV [22], SLEV [22], TENV [22], VEEV [22], WNV [24] |
| Culex (Cux.) quinquefasciatus Say | 57 | Municipal Parks of São Paulo City ** | 2010/2011 | CHIKV [22], EEEV [22], MAYV [28], OROV [22], ROCV [22], SLEV [22], VEEV [22], WNV [22], ZIKV [22], Wuchereria bancrofti [9], Dirofilaria immitis [26] |
| Group 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Culex acharistus | Culex ameliae | Culex bidens | Culex chidesteri | Culex coronator | Culex declarator | Culex dolosus | Culex eduardoi | Culex habilitator | Culex nigripalpus | Culex quinquefasciatus | ||
| Group 1 | Culex acharistus | - | 90 | 89 | 89 | 83 | 46 | 97 | 73 | 91 | 88 | 91 |
| Culex ameliae | 88 | - | 44 | 83 | 74 | 77 | 100 | 100 | 66 | 74 | 91 | |
| Culex bidens | 84 | 57 | - | 79 | 81 | 46 | 100 | 87 | 69 | 62 | 94 | |
| Culex chidesteri | 92 | 87 | 96 | - | 93 | 62 | 91 | 80 | 94 | 91 | 89 | |
| Culex coronator | 88 | 83 | 67 | 72 | - | 62 | 84 | 67 | 47 | 88 | 91 | |
| Culex declarator | 64 | 80 | 48 | 59 | 81 | - | 94 | 73 | 59 | 82 | 94 | |
| Culex dolosus | 100 | 93 | 93 | 90 | 91 | 77 | - | 53 | 97 | 100 | 94 | |
| Culex eduardoi | 72 | 97 | 81 | 86 | 91 | 92 | 63 | - | 81 | 94 | 71 | |
| Culex habilitator | 96 | 73 | 67 | 86 | 58 | 46 | 88 | 67 | - | 85 | 94 | |
| Culex nigripalpus | 100 | 77 | 63 | 79 | 81 | 85 | 100 | 93 | 78 | - | 97 | |
| Culex quinquefasciatus | 92 | 100 | 100 | 100 | 91 | 100 | 100 | 60 | 100 | 97 | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, R.F.; Wilke, A.B.B.; Chagas, C.R.F.; Menezes, R.M.T.d.; Suesdek, L.; Multini, L.C.; Silva, F.S.; Grech, M.G.; Marrelli, M.T.; Kirchgatter, K. Wing Geometric Morphometrics as a Tool for the Identification of Culex Subgenus Mosquitoes of Culex (Diptera: Culicidae). Insects 2020, 11, 567. https://doi.org/10.3390/insects11090567
Simões RF, Wilke ABB, Chagas CRF, Menezes RMTd, Suesdek L, Multini LC, Silva FS, Grech MG, Marrelli MT, Kirchgatter K. Wing Geometric Morphometrics as a Tool for the Identification of Culex Subgenus Mosquitoes of Culex (Diptera: Culicidae). Insects. 2020; 11(9):567. https://doi.org/10.3390/insects11090567
Chicago/Turabian StyleSimões, Roseli França, André Barretto Bruno Wilke, Carolina Romeiro Fernandes Chagas, Regiane Maria Tironi de Menezes, Lincoln Suesdek, Laura Cristina Multini, Fabiana Santos Silva, Marta Gladys Grech, Mauro Toledo Marrelli, and Karin Kirchgatter. 2020. "Wing Geometric Morphometrics as a Tool for the Identification of Culex Subgenus Mosquitoes of Culex (Diptera: Culicidae)" Insects 11, no. 9: 567. https://doi.org/10.3390/insects11090567