The Potential for Decision Support Tools to Improve the Management of Root-Feeding Fly Pests of Vegetables in Western Europe

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Approaches Used to Monitor Pest Infestations
3. Approaches Used to Forecast Pest Infestations
4. Use of Decision Support
4.1. Insecticides
4.2. Physical Control
4.3. Avoidance through the Adjustment of Planting or Harvesting Dates
4.4. Biological Control
4.5. Manipulation of Pest Behaviour
5. Climate Change
6. Benefits Provided by the Use of Decision Support Tools
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellis, P.R. The identification and exploitation of resistance in carrots and wild umbelliferae to the carrot fly, Psila rosae (f.). Integr. Pest Manag. Rev. 1999, 4, 259–268. [Google Scholar] [CrossRef]
- Ticheler, J.; Loosjes, M.; Noorlander, J. Sterile-insect technique for control of the onion maggot, Delia antiqua. Integr. Control. Insect Pests Neth. 1980, 93–97. [Google Scholar]
- Leger, C.; Riga, E. Evaluation of marigolds and entomopathogenic nematodes for control of the cabbage maggotdelia radicum. J. Sustain. Agric. 2009, 33, 128–141. [Google Scholar] [CrossRef]
- Davidson, G.; Chandler, D. Laboratory Evaluation of Entomopathogenic Fungi Against Larvae and Adults of Onion Maggot (Diptera: Anthomyiidae). J. Econ. Entomol. 2005, 98, 1848–1855. [Google Scholar] [CrossRef] [PubMed]
- Chandler, D.; Davidson, G. Evaluation of Entomopathogenic Fungus Metarhizium anisopliae Against Soil-Dwelling Stages of Cabbage Maggot (Diptera: Anthomyiidae) in Glasshouse and Field Experiments and Effect of Fungicides on Fungal Activity. J. Econ. Entomol. 2005, 98, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Myrand, V.; Buffet, J.P.; Guertin, C. Susceptibility of Cabbage Maggot Larvae (Diptera: Anthomyiidae) to Hypocreales Entomopathogenic Fungi. J. Econ. Entomol. 2015, 108, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Herbst, M.; Hommes, M. Influence of soil moisture and type on entomopathogenic nematode and predatory mite performance in cabbage root fly control. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Hamburg, Germany, 4–7 October 2015. [Google Scholar]
- Vernon, R.S.; McGregor, R.R. Exclusion fences reduce colonization of carrots by the carrot rust fly, Psila rosae (Diptera: Psilidae). J. Entomol. Soc. Br. Columbia 1999, 96, 103–109. [Google Scholar]
- Bomford, M.K.; Vernon, R.S.; Pats, P. Importance of collection overhangs on the efficacy of exclusion fences for managing cabbage flies (Diptera: Anthomyiidae). Environ. Entomol. 2000, 29, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Siekmann, G.; Hommes, M. Exclusion fences against cabbage root fly and carrot fly. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Ljubljana, Slovenia, 3–7 October 2005. [Google Scholar]
- Witkowska, E.; Moorhouse, E.R.; Jukes, A.; Elliott, M.S.; Collier, R.H. Implementing Integrated Pest Management in commercial crops of radish (Raphanus sativus). Crop Prot. 2018, 114, 148–154. [Google Scholar] [CrossRef]
- Finch, S.; Collier, R. Integrated pest management in field vegetable crops in northern Europe—With focus on two key pests. Crop. Prot. 2000, 19, 817–824. [Google Scholar] [CrossRef]
- Finch, S.; Billiald, H.; Collier, R.H. Companion planting—Do aromatic plants disrupt host-plant finding by the cabbage root fly and the onion fly more effectively than non-aromatic plants? Entomol. Exp. Appl. 2003, 109, 183–195. [Google Scholar] [CrossRef]
- Rousse, P.; Fournet, S.; Porteneuve, C.; Brunel, E. Trap cropping to control Delia radicum populations in cruciferous crops: First results and future applications. Entomol. Exp. Appl. 2003, 109, 133–138. [Google Scholar] [CrossRef]
- George, D.; Port, G.; Collier, R. Selection of trap crop and companion plants for the management of pest insects in field vegetables. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Ljubljana, Slovenia, 3–7 October 2005. [Google Scholar]
- Ferry, A.; Le Tron, S.; Dugravot, S.; Cortesero, A.M. Field evaluation of the combined deterrent and attractive effects of dimethyl disulfide on Delia radicum and its natural enemies. Biol. Control 2009, 49, 219–226. [Google Scholar] [CrossRef]
- Lamy, F.C.; Poinsot, D.; Cortesero, A.-M.; Dugravot, S. Artificially applied plant volatile organic compounds modify the behavior of a pest with no adverse effect on its natural enemies in the field: Improving the push–pull strategy against a major Brassicaceae pest. J. Pest Sci. 2017, 90, 611–621. [Google Scholar] [CrossRef]
- Lamy, F.; Dugravot, S.; Cortesero, A.M.; Chaminade, V.; Faloya, V.; Poinsot, D. One more step toward a push-pull strategy combining both a trap crop and plant volatile organic compounds against the cabbage root fly Delia radicum. Environ. Sci. Pollut. Res. 2017, 25, 29868–29879. [Google Scholar] [CrossRef] [PubMed]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Damos, P.T. Modular structure of web-based decision support systems for integrated pest management. A review. Agron. Sustain. Dev. 2015, 35, 1347–1372. [Google Scholar] [CrossRef] [Green Version]
- Finch, S.; Collier, R.H.; Skinner, G. Local population differences in emergence of cabbage root flies from south-west Lancashire: implications for pest forecasting and population divergence. Ecol. Entomol. 1986, 11, 139–145. [Google Scholar] [CrossRef]
- Johansen, T.J.; Meadow, R. Diapause development in early and late emerging phenotypes of Delia floralis. Insect Sci. 2013, 21, 103–113. [Google Scholar] [CrossRef]
- Finch, S.; Skinner, G. Some factors affecting the efficiency of water-traps for capturing cabbage root flies. Ann. Appl. Boil. 1974, 77, 213–226. [Google Scholar] [CrossRef]
- Finch, S. Improving the selectivity of water traps for monitoring populations of the cabbage root fly. Ann. Appl. Boil. 1992, 120, 1–7. [Google Scholar] [CrossRef]
- Finch, S.; Skinner, G.; Freeman, G.H. The distribution and analysis of cabbage root fly egg populations. Ann. Appl. Boil. 1975, 79, 1–18. [Google Scholar] [CrossRef]
- Bligaard, J.; Meadow, R.; Nielsen, O.; Percy-Smith, A. Evaluation of felt traps to estimate egg numbers of cabbage root fly, Delia radicum, and turnip root fly, Delia floralis in commercial crops. Ѐntomol. Exp. Appl. 1999, 90, 141–148. [Google Scholar] [CrossRef]
- Libby, J.L.; Chapman, R.K.; Wyman, J.A. Aspects of cabbage maggot management in Wisconsin. Proc. North Cent. Branch Entomol. Soc. Am. 1975, 30, 77–78. [Google Scholar]
- Vernon, R.S.; Broatch, J.S. Responsiveness of Delia spp. (Diptera: Anthomyiidae) to colored sticky traps in flowering and rosette stage canola. Can. Entomol. 1996, 128, 1077–1085. [Google Scholar] [CrossRef]
- Kuhar, T.P.; Hutchison, W.D.; Whalen, J.; Riley, D.G.; Meneley, J.C.; Doughty, H.B.; Burkness, E.C.; Wold-Burkness, S.J. Field evaluation of a novel lure for trapping seedcorn maggot adults. Plant Health Prog. 2006, 7, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.A.; Scatcherd, J.E. Bean seed fly (Delia platura, Delia florilega) and onion fly (Delia antiqua) incidence in England and an evaluation of chemical and biological control options. Ann. Appl. Biol. 2007, 151, 259–267. [Google Scholar] [CrossRef]
- Otto, M. Populationsökologische Untersuchungen zur Spargelfliege (Platyparea poeciloptera) und Zwiebelfliege (Delia antiqua) unter Besonderer Berücksichtigung des Einsatzes von Simulationsmodellen im Integrierten Pflanzenschutz [Population Ecologic Studies on the Asparagus Fly and the Onion Fly with Particular Focus on the Use of Simulation Models in Integrated Plant Protection]. Ph.D. Thesis, Universität Bayreuth, Bayreuth, Germany, 2002. [Google Scholar]
- Vernon, R.S. A spectral zone of color preference for the onion fly, Delia antiqua (diptera: Anthomyiidae), with reference to the reflective intensity of traps. Can. Entomol. 1986, 118, 849–856. [Google Scholar] [CrossRef]
- Judd, O.J.R.; Whitfield, G.H. Visual and olfactory behaviour of Delia antiqua (Diptera: Anthomyiidae) in relation to time of day and ovarian development. Eur. J. Entomol. 1997, 94, 199–209. [Google Scholar]
- Thöming, G.; Schjøll, A.F.; Johansen, T.J. Developing tools for monitoring and forecasting of onion fly Delia antiqua in Norway. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Arenenberg, Switzerland, 2–6 October 2017. [Google Scholar]
- Collier, R.H.; Finch, S. Some factors affecting the efficiency of sticky board traps for capturing the carrot fly, Psila rosae (Diptera, Psilidae). Bull. Entomol. Res. 1990, 80, 153–158. [Google Scholar] [CrossRef]
- Finch, S.; Collier, R.H. Effects of the angle of inclination of traps on the numbers of large Diptera caught on sticky boards in certain vegetable crops. Entomol. Exp. Appl. 1989, 52, 23–27. [Google Scholar] [CrossRef]
- Collier, R. Review of carrot fly control in Northern Europe—2009. EPPO Bull. 2009, 39, 116–120. [Google Scholar] [CrossRef]
- Bligaard, J. Damage thresholds for cabbage root fly [Delia radicum (L.)] in cauliflower assessed from pot experiments. Acta Agric. Scand. Sect. B Soil Plant Sci. 1999, 49, 57–64. [Google Scholar] [CrossRef]
- Maack, G. Schadwirkung der Kleinen Kohlfliege (Phorbia Brassicae Bouché) und Möglichkeiten zur Reduzierung des Insektizidaufwandes bei der Bekämpfung [Damage of the Cabbage Root Fly and Possibilities to Reduce the Amount of Insecticides for Control]. Ph.D. Thesis, Technische Universität Berlin, Berlin, Germany, 1977. [Google Scholar]
- Eckenrode, C.; Chapman, R.K. Effect of various temperatures upon rate of development of the cabbage maggot under artificial conditions. Ann. Entomol. Soc. Am. 1971, 64, 1079–1083. [Google Scholar] [CrossRef]
- Eckenrode, C.J.; Chapman, R.K. Seasonal adult cabbage maggot populations in the field in relation to thermal-unit accumulations. Ann. Entomol. Soc. Am. 1972, 65, 151–156. [Google Scholar] [CrossRef]
- Vincent, C.; Stewart, R. Évaluation de deux types de pièges pour le dépistage des adultes de la mouche du chou, Hylemya (Delia) brassicae (Wiedemann)(Diptera: Anthomyiidae). Ann. Entomol. Soc. Que 1981, 26, 41–50. [Google Scholar]
- Funderburk, J.; Higley, L.; Pedigo, L. Seedcorn maggot (Diptera: Anthomyiidae) phenology in central Iowa and examination of a thermal-unit system to predict development under field conditions. Environ. Entomol. 1984, 13, 105–109. [Google Scholar] [CrossRef]
- Sanborn, S.; Wyman, J.; Chapman, R. Threshold temperature and heat unit summations for seedcorn maggot development under controlled conditions. Ann. Entomol. Soc. Am. 1982, 75, 103–106. [Google Scholar] [CrossRef]
- Liu, H.; McEwen, F.; Ritcey, G. Forecasting events in the life cycle of the onion maggot, Hylemya antiqua (Diptera: Anthomyiidae): Application to control schemes. Environ. Entomol. 1982, 11, 751–755. [Google Scholar] [CrossRef]
- UC IPM Research and IPM. Models Insects, Mites, Diseases, Plants and Beneficials. Available online: http://ipm.ucanr.edu/MODELS/index.html (accessed on 5 May 2020).
- Phelps, K.; Collier, R.H.; Reader, R.J.; Finch, S. Monte Carlo simulation method for forecasting the timing of pest insect attacks. Crop Prot. 1993, 12, 335–342. [Google Scholar] [CrossRef]
- Hommes, M.; Gebelein, D. Simulation models for the cabbage root fly and the carrot fly. Integrated Control in Field Vegetable Crops. In Proceedings of the IOBC WPRS Bulletin, Working Group “Integrated Protection in Field Vegetables”, Guitté, France, 6–8 November 1995. [Google Scholar]
- Johansen, T.J.; Meadow, R. Emergence patterns of Norwegian brassica root fly populations. In Proceedings of the IOBC/WPRS Bulletin, Working Group “Integrated Protection in Field Vegetables”, Deinze, Belgium, 12–16 October 2003. [Google Scholar]
- Johansen, T.J.; Meadow, R. Population differences in emergence of brassica root flies (Diptera: Anthomyiidae). Environ. Entomol. 2006, 35, 1161–1165. [Google Scholar] [CrossRef]
- VIPS, Varsling Innen PlanteSkadegjørere. Available online: www.vips-landbruk.no (accessed on 5 May 2020).
- SEGES. Available online: https://www.landbrugsinfo.dk/Planteavl/Plantevaern/Skadedyr/Skadedyrsarter/ Sider/PL_Kaalflue_varsling.aspx (accessed on 5 May 2020).
- Syngenta, AHDB Pest Bulletin. Available online: https://www.syngenta.co.uk/ahdb-pest-bulletin (accessed on 4 June 2020).
- Information System for Integrated Plant Production (ISIP). Available online: https://isip.de/isip/servlet/isip-de (accessed on 5 May 2020).
- Bulletins de Santé du Végétal (BSV). Available online: http://draaf.bretagne.agriculture.gouv.fr/Bulletins-de-Sante-du-Vegetal-BSV (accessed on 5 May 2020).
- European Commission, Neonicotinoids. Available online: https://ec.europa.eu/food/plant/pesticides/ approval_active_substances/approval_renewal/neonicotinoids_en (accessed on 5 May 2020).
- Collier, R.; Finch, S. A review of research to address carrot fly (Psila rosae) control in the UK. EPPO Bull. 2009, 39, 121–127. [Google Scholar] [CrossRef]
- Collier, R. Optimising Carrot Fly Control Using Pyrethroids and Coragen®. AHDB Project FV 414. Unpublished Data 2014. Available online: https://ahdb.org.uk/fv-414-carrots-optimising-carrot-fly-control-using-pyrethroids-and-coragen (accessed on 10 June 2020).
- Markkula, I.; Ojanen, H.; Tiilikkala, K. Forecasting and monitoring of the carrot fly (Psila rosae) in Finland. Pests Dis. 1998, 2, 319–705. [Google Scholar]
- Johansen, T.J. Monitoring and control of the carrot fly (Psila rosae fabr.) in northern Norway. Acta Agric. Scand. Sect. B Soil Plant Sci. 1999, 49, 158–166. [Google Scholar] [CrossRef]
- Folkedal Schjøll, A.; (Norwegian Institute of Bioeconomy Research (NIBIO), Division of Biotechnology and Plant Health, Ås, Norway). Personal communication, 2007.
- Collier, R.; (Warwick Crop Centre, School of Life Sciences, University of Warwick, Wellesbourne, Warwick, UK). Personal communication, 2019.
- Collier, R.H.; Phelps, K. Using trapping data to decide whether or not to spray against the carrot fly. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Guitté, France, 6–8 November 1995. [Google Scholar]
- Hawkes, C. The diurnal periodicity and cycle of behaviour of the adult cabbage root fly (Erioschia brassicae). Ann. Appl. Biol. 1972, 70, 109–118. [Google Scholar] [CrossRef]
- Meadow, R.; Johansen, T.J. Exclusion fences against brassica root flies (Delia radicum and D. floralis). In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Deinze, Belgium, 12–16 October 2003. [Google Scholar]
- Silver, N.; Hillier, K.; Blatt, S. Management of Delia (Diptera: Anthomyiidae) through selectively timed planting of Phaseolus vulgaris (Fabaceae) in Atlantic Canada. Can. Entomol. 2018, 150, 663–674. [Google Scholar] [CrossRef]
- Nault, B.A.; Werling, B.P.; Straub, R.W.; Nyrop, J.P. Delaying onion planting to control onion maggot (Diptera: Anthomyiidae): Efficacy and underlying mechanisms. J. Econ. Entomol. 2011, 104, 1622–1632. [Google Scholar] [CrossRef]
- Herrmann, F.; Buck, H.; Hommes, M.; Saucke, H. Schlagseparierung als Ansatz zur Prävention von Möhrenfliegenschäden. [Spatial risk avoidance as a strategy to prevent carrot fly damage]. Wiss. Ökologischer Landbau Gießen 2011, 11, 15–18. [Google Scholar]
- Herrmann, F.; Buck, H.; Liebig, N.; Hommes, M.; Saucke, H. Vermeidung und Reduktion von Möhrenfliegenschäden im Ökolandbau [Prevention and reduction of damage by the carrot fly in organic production]. Wiss. Ökologischer Landbau 2009, 10, 11–13. [Google Scholar]
- Johnsson, B. Forecasting the timing of damage by the carrot fly. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Vienna, Austrlia, 28–30 October 1991. [Google Scholar]
- Finch, S.; Collier, R.H. Parasitism of overwintering pupae of cabbage root fly, Delia radicum (l) (Diptera: anthomyiidae), in England and Wales. Bull. Entomol. Res. 1984, 74, 79–86. [Google Scholar] [CrossRef]
- Meyling, N.V.; Navntoft, S.; Philipsen, H.; Thorup-Kristensen, K.; Eilenberg, J. Natural regulation of Delia radicum in organic cabbage production. Agric. Ecosyst. Environ. 2013, 164, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Finch, S.; Elliott, M.S.; Torrance, M.T. Is the parasitoid staphylinid beetle Aleochara bilineata an effective predator of the egg stage of its natural host, the cabbage root fly? In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Chania, Crete, 6–8 October 1997. [Google Scholar]
- Bennison, J.; Lole, M.; Pope, T.; Maulden, K.; Maher, H.; Watling, M. Potential control of cabbage root fly (Delia radicum) with the predatory staphylinid beetle Atheta coriaria. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Dubrovnik, Croatia, 4–7 October 2009. [Google Scholar]
- Herbst, M. Evaluierung Nachhaltiger Komponenten für den Integrierten Pflanzenschutz zur Bekämpfung der Kleinen Kohlfliege (Delia Radicum (L.)) im Kohlanbau [Evaluation of Sustainable Factors for an Integrated Control of the Cabbage Root Fly]. Ph.D. Thesis, Gottfried Wilhelm Leibniz Universität Hannover, Freistaat Sachsen, Germany, 2017. [Google Scholar]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. Biocontrol 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Josso, C.; Le Ralec, A.; Raymond, L.; Saulais, J.; Baudry, J.; Poinsot, D.; Cortesero, A.M. Effects of field and landscape variables on crop colonization and biological control of the cabbage root fly. Delia Radicum Landsc. Ecol. 2013, 28, 1697–1715. [Google Scholar] [CrossRef]
- Prasad, R.; Snyder, W. Predator interference limits fly egg biological control by a guild of ground-active beetles. Biol. Control 2004, 31, 428–437. [Google Scholar] [CrossRef]
- Prasad, R.; Snyder, W. Polyphagy complicates conservation biological control that targets generalist predators. J. Appl. Ecol. 2006, 43, 343–352. [Google Scholar] [CrossRef]
- Razinger, J.; Lutz, M.; Schroers, H.-J.; Urek, G.; Grunder, J. Evaluation of insect associated and plant growth promoting fungi in the control of cabbage root flies. J. Econ. Ѐntomol. 2014, 107, 1348–1354. [Google Scholar] [CrossRef] [Green Version]
- Cotes, B.; Rännbäck, L.-M.; Björkman, M.; Norli, H.R.; Meyling, N.V.; Rämert, B.; Anderson, P. Habitat selection of a parasitoid mediated by volatiles informing on host and intraguild predator densities. Oecologia 2015, 179, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Rännbäck, L.-M.; Cotes, B.; Anderson, P.; Rämert, B.; Meyling, N.V. Mortality risk from entomopathogenic fungi affects oviposition behavior in the parasitoid wasp Trybliographa rapae. J. Invertebr. Pathol. 2015, 124, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Finch, S.; Collier, R. Host-plant selection by insects - a theory based on ‘appropriate/inappropriate landings’ by pest insects of cruciferous plants. Ѐntomol. Exp. Appl. 2000, 96, 91–102. [Google Scholar] [CrossRef]
- Finch, S.; Collier, R.H. The influence of host and non-host companion plants on the behaviour of pest insects in field crops. Ѐntomol. Exp. Appl. 2011, 142, 87–96. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Dugravot, S.; Mortreuil, A.; Le Ralec, A.; Cortesero, A. Selecting volatiles to protect brassicaceous crops against the cabbage root fly, Delia radicum. Ѐntomol. Exp. Appl. 2012, 144, 69–77. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Dugravot, S.; Danner, H.; Van Dam, N.M.; Cortesero, A.M. Characterizing Volatiles and Attractiveness of Five Brassicaceous Plants with Potential for a ‘Push-Pull’ Strategy Toward the Cabbage Root Fly, Delia radicum. J. Chem. Ecol. 2015, 41, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamy, F.; Bellec, L.; Rusu-Stievenard, A.; Clin, P.; Ricono, C.; Olivier, D.; Mauger, S.; Poinsot, D.; Faloya, V.; Daniel, L.; et al. Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates. Insects 2020, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.R.; Pickett, J.A. Push-pull strategy for insect pest management. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Miller, J.R.; Cowles, R.S. Stimulo-deterrent diversion: A concept and its possible application to onion maggot control. J. Chem. Ecol. 1990, 16, 3197–3212. [Google Scholar] [CrossRef] [PubMed]
- Deguine, J.; Rousse, P.; Atiama-Nurbel, T. Agroecological Crop Protection: Concepts and a Case Study from Reunion. Integr. Pest Manag. Pest Control Curr. Future Tactics 2012, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Climate Change Service. Available online: https://climate.copernicus.eu/ (accessed on 5 June 2020).
- Kovats, R.S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.-F. Europe. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 1267–1326. [Google Scholar]
- Norwegian Environment Agency. Available online: https://www.regjeringen.no/en/dep/kld/organisation/Subordinate-agencies/norwegian-environment-agency/id85642/ (accessed on 5 May 2020).
- Met Office. UK Climate Projections: Headline Findings Crown Copyright 2019; Met Office: London, UK, 2019. [Google Scholar]
- Ning, S.; Wei, J.; Feng, J. Predicting the current potential and future world wide distribution of the onion maggot, Delia antiqua using maximum entropy ecological niche modeling. PLoS ONE 2017, 12, e0171190. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.H.; Finch, S.; Phelps, K.; Thompson, A.R. Possible impact of global warming on cabbage root fly (Delia radicum) activity in the uk. Ann. Appl. Biol. 1991, 118, 261–271. [Google Scholar] [CrossRef]
- Sauer, C. Possible impacts of climate change on carrot fly’s population dynamics in Switzerland. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Arenenberg, Switzerland, 2–6 October 2017. [Google Scholar]
- Villeneuve, F.; Latour, F. A few biological specificities of the carrot root fly (Psila rosae Fabre) for a more accurate forecasting in Carrot (Daucus carota L.) production. In Proceedings of the International Symposium on carrot and other Apiaceae, Angers, France, 17–19 September 2014. [Google Scholar]
- Ferguson, A.W.; Skellern, M.P.; Johnen, A.; Von Richthofen, J.S.; Watts, N.P.; Bardsley, E.; Murray, D.A.; Cook, S.M. The potential of decision support systems to improve risk assessment for pollen beetle management in winter oilseed rape. Pest Manag. Sci. 2016, 72, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Collier, R.; Elliott, M.; Wilson, D.; Teverson, D.; Cowgill, S. Phenology and abundance of pest insects of vegetable and salad crops in Britain: Decision support for growers. In Proceedings of the Working Group “Integrated Protection in Field Vegetables”, Stratford-upon-Avon, UK, 13–16 October 2020. [Google Scholar]




{kind=link}
{kind=link}
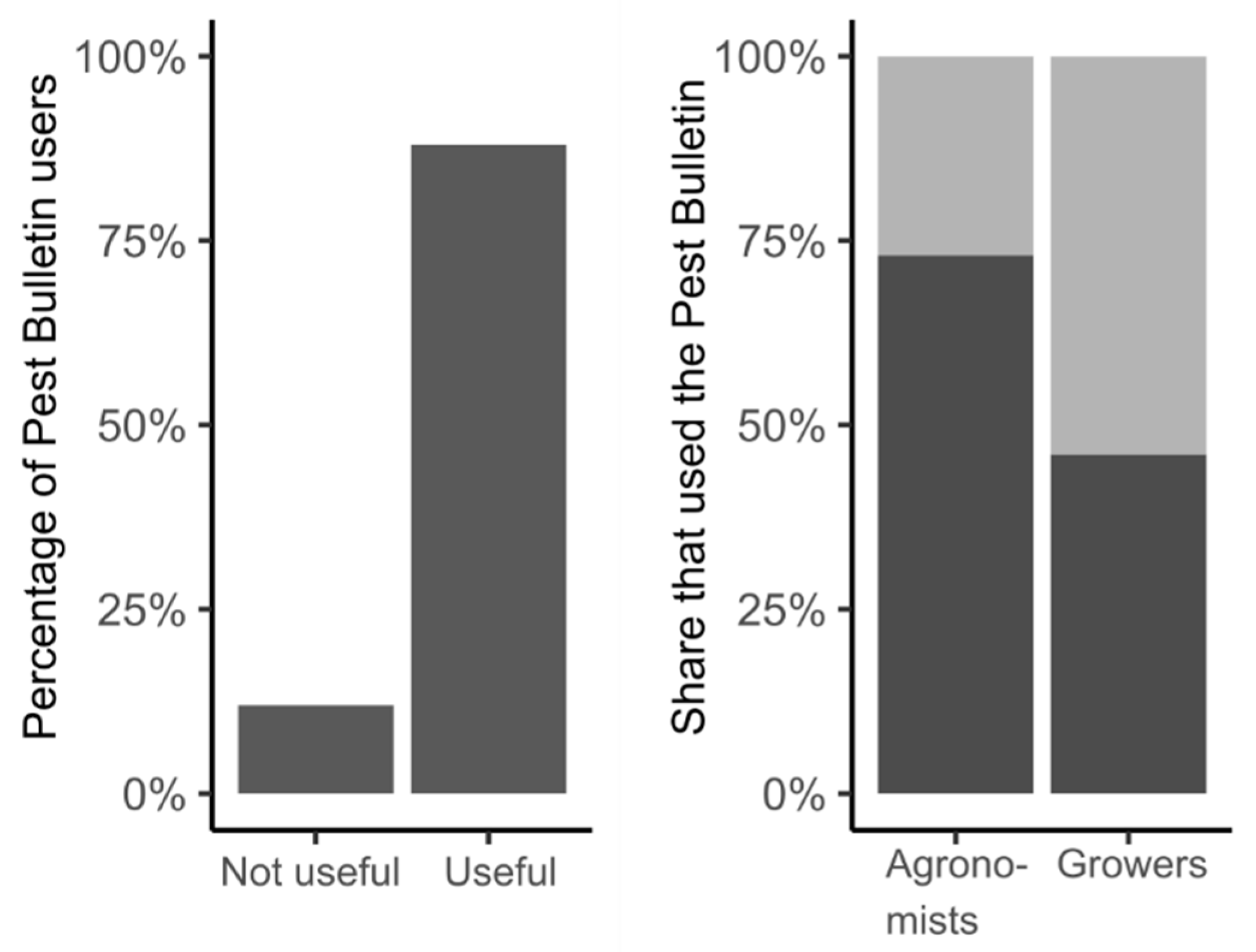{kind=link}
| Pest Insect | Common Name | Plant Family Affected | Monitoring Systems | Forecasting Systems | Thresholds Available |
|---|---|---|---|---|---|
| Delia radicum | Cabbage root fly | Brassicaceae | Traps using vision and olfaction, egg sampling | Degree-day models, simulation models | Norway, France |
| Delia floralis | Turnip fly | Brassicaceae | Traps using vision, egg sampling | Warnings are disseminated based on egg counts which are related to damage thresholds | Norway |
| Delia platura | Bean seed fly | Various | Traps using vision and olfaction | Degree-day models | No |
| Delia florilega | Root fly | Various | Traps using vision and olfaction | No | No |
| Delia antiqua | Onion fly | Alliaceae | Traps using vision | Degree-day models, simulation model | No |
| Chamaepsila rosae | Carrot fly | Asteraceae | Traps using vision | Degree-day models, simulation models | Several countries |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collier, R.; Mazzi, D.; Folkedal Schjøll, A.; Schorpp, Q.; Thöming, G.; Johansen, T.J.; Meadow, R.; Meyling, N.V.; Cortesero, A.-M.; Vogler, U.; et al. The Potential for Decision Support Tools to Improve the Management of Root-Feeding Fly Pests of Vegetables in Western Europe. Insects 2020, 11, 369. https://doi.org/10.3390/insects11060369
Collier R, Mazzi D, Folkedal Schjøll A, Schorpp Q, Thöming G, Johansen TJ, Meadow R, Meyling NV, Cortesero A-M, Vogler U, et al. The Potential for Decision Support Tools to Improve the Management of Root-Feeding Fly Pests of Vegetables in Western Europe. Insects. 2020; 11(6):369. https://doi.org/10.3390/insects11060369
Chicago/Turabian StyleCollier, Rosemary, Dominique Mazzi, Annette Folkedal Schjøll, Quentin Schorpp, Gunda Thöming, Tor J. Johansen, Richard Meadow, Nicolai V. Meyling, Anne-Marie Cortesero, Ute Vogler, and et al. 2020. "The Potential for Decision Support Tools to Improve the Management of Root-Feeding Fly Pests of Vegetables in Western Europe" Insects 11, no. 6: 369. https://doi.org/10.3390/insects11060369