Coprophagous Hydrophilid Beetles (Coleoptera, Hydrophilidae, Sphaeridiinae) Distribution in the Polish Carpathians

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Sampling Method (Collection of Dung Beetles)
2.3. Nomenclature and Systematics
2.4. Data Analyses
3. Results
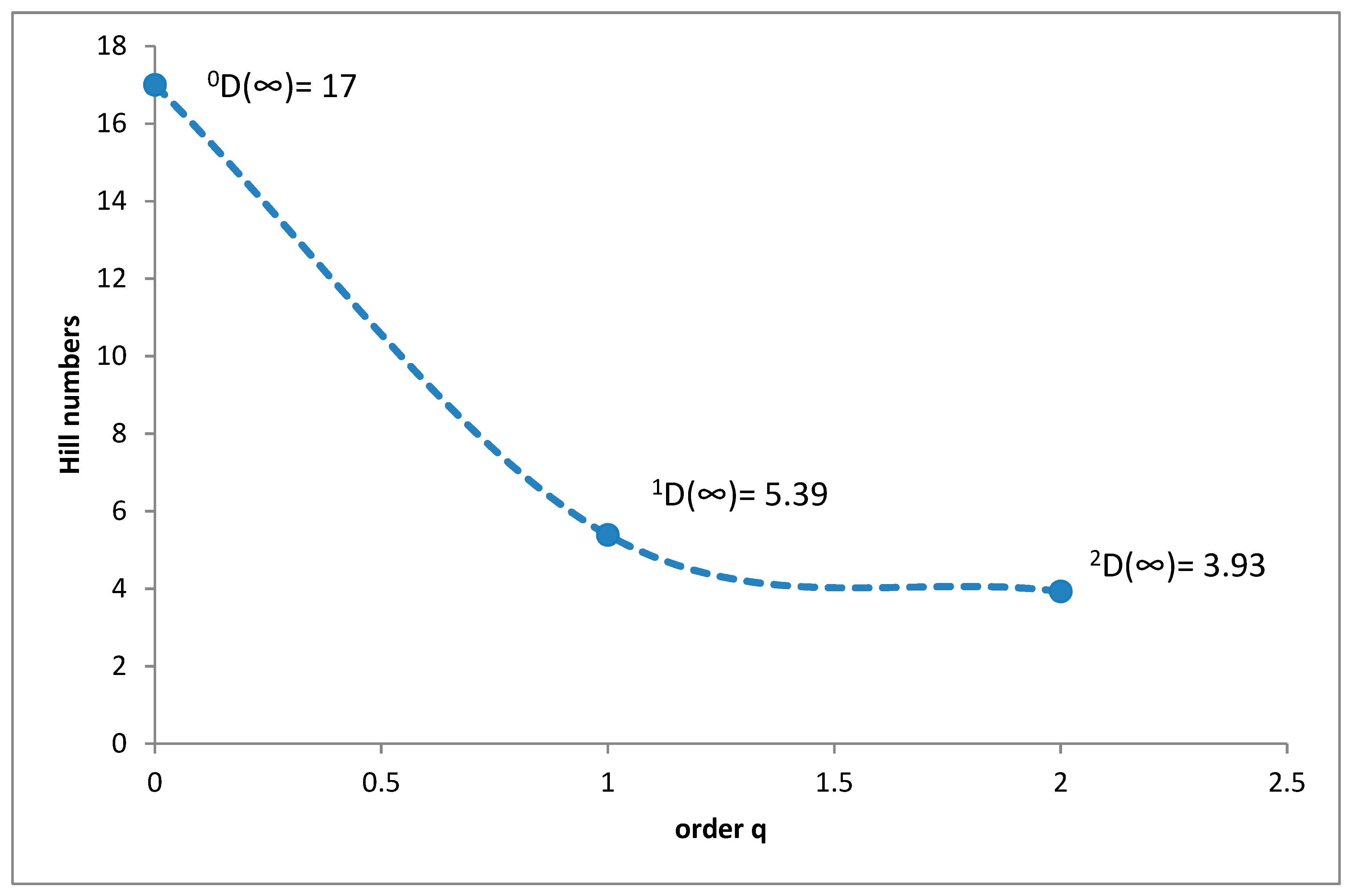
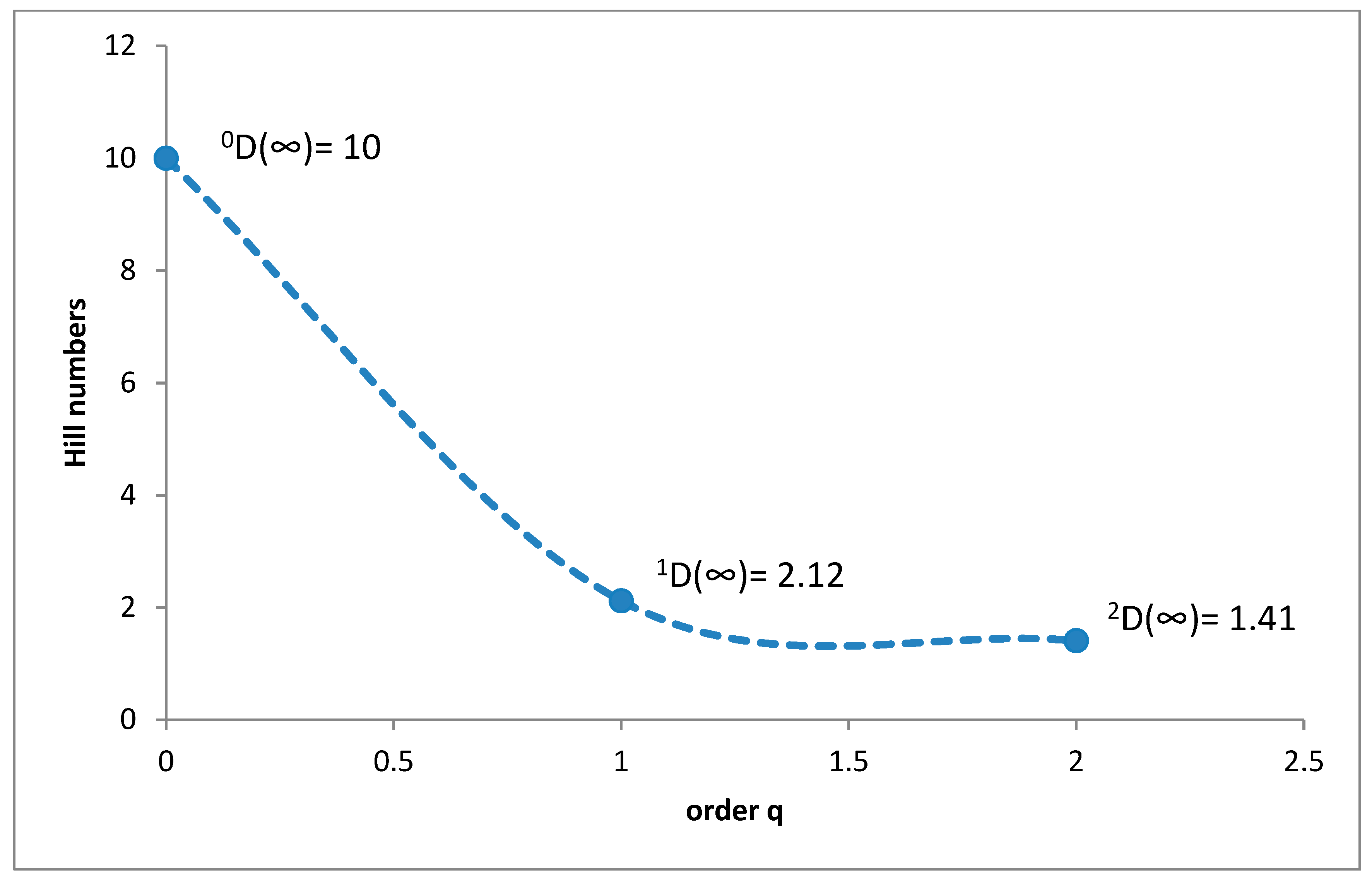
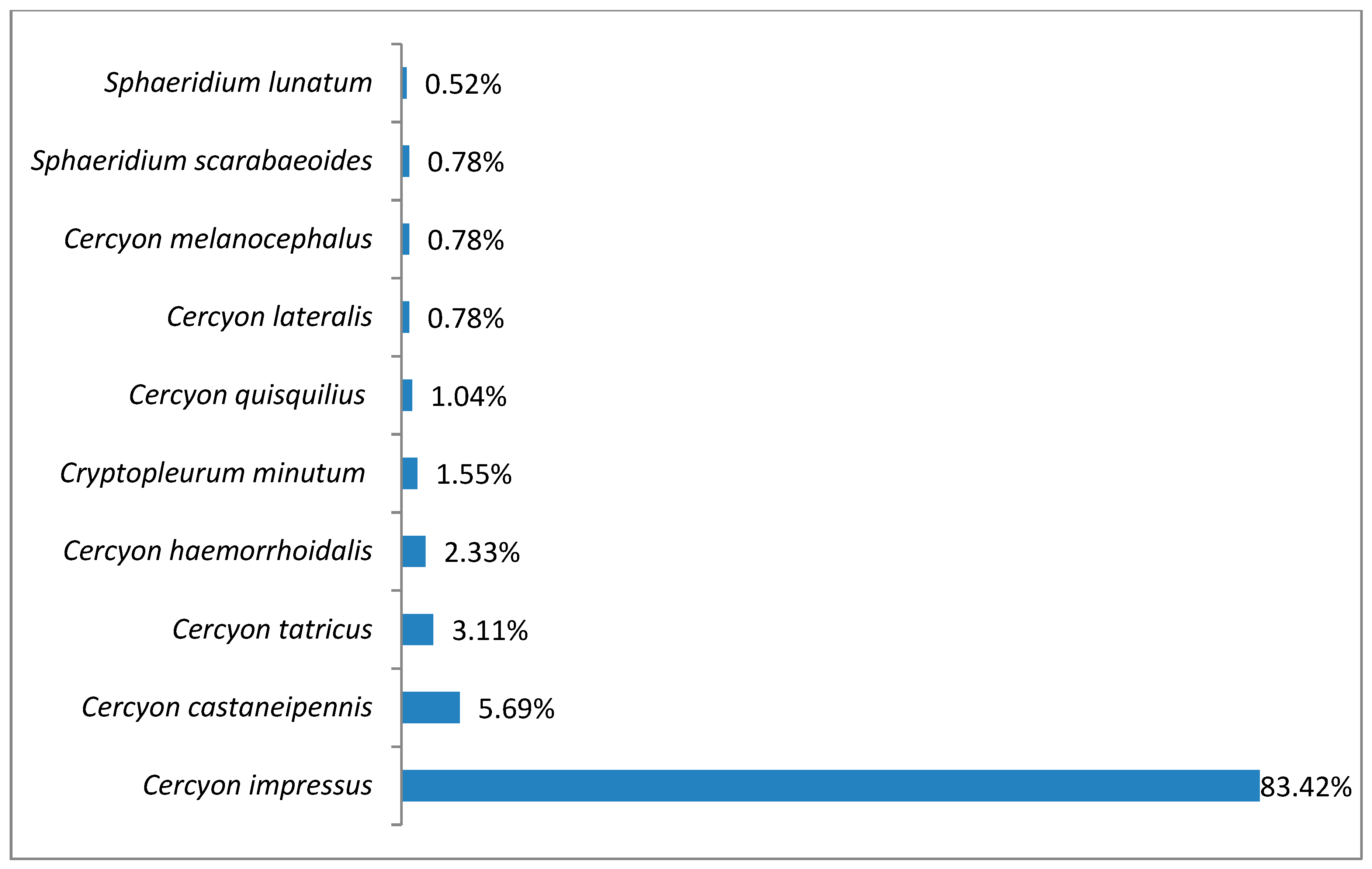
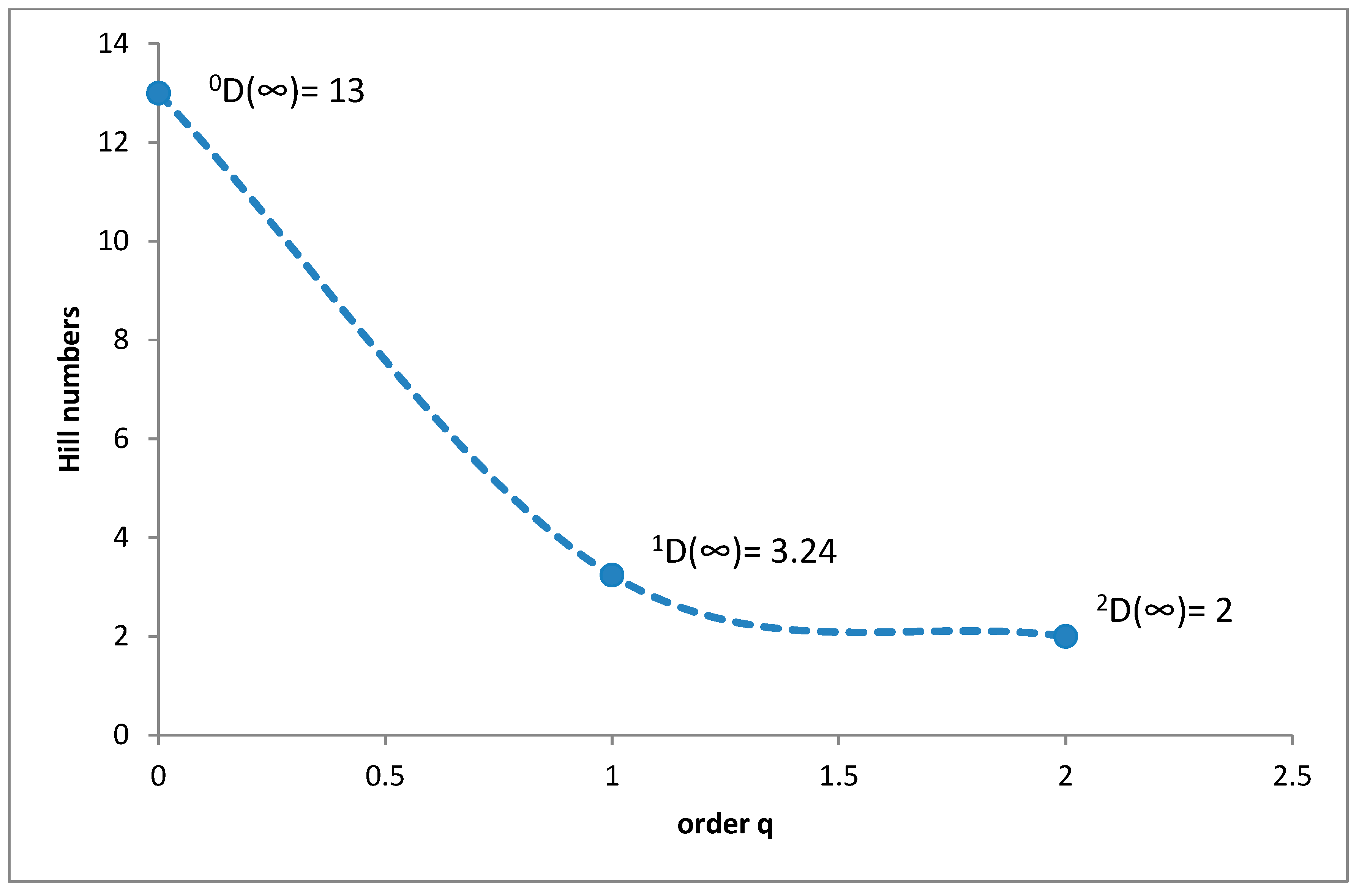
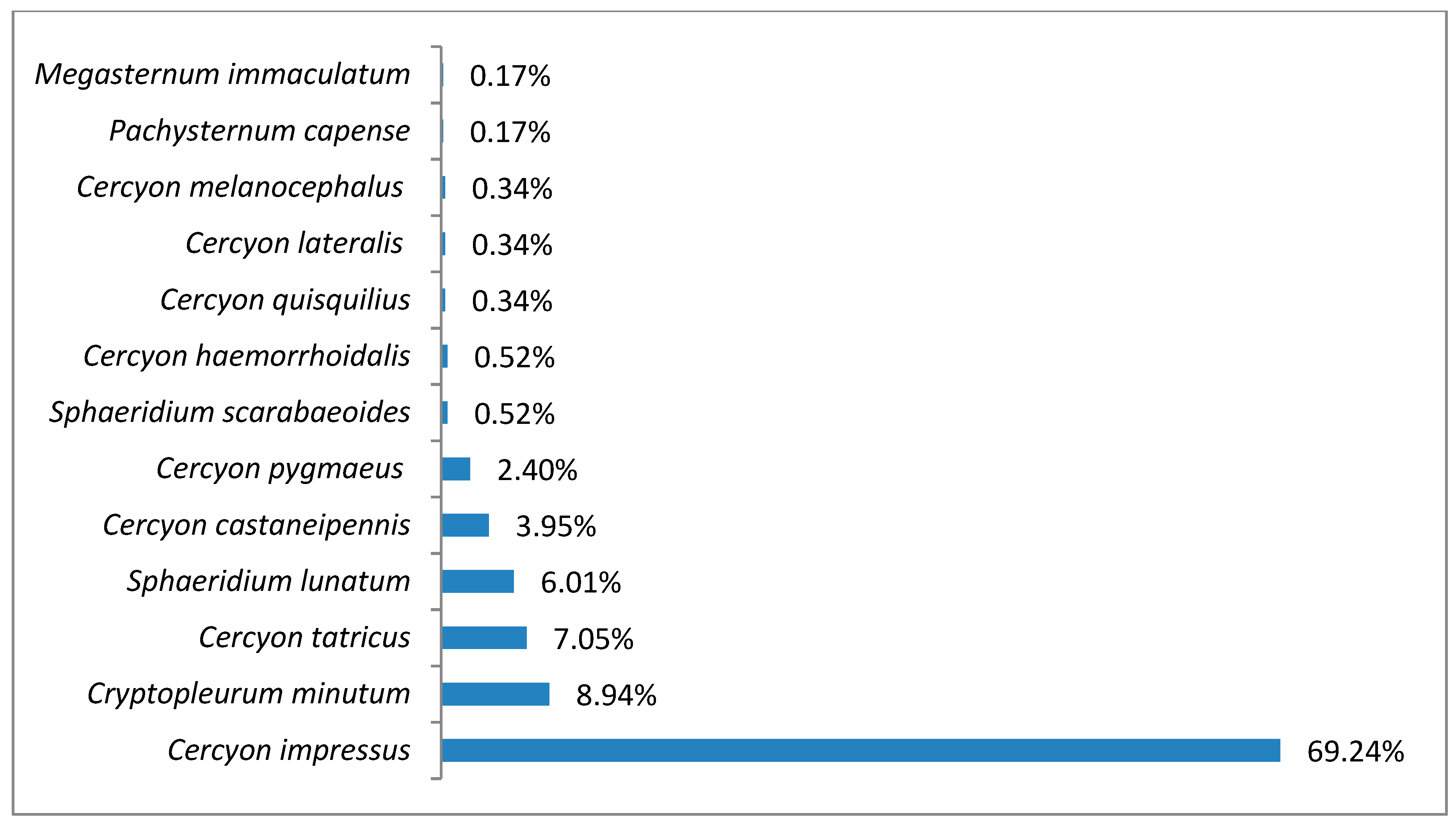
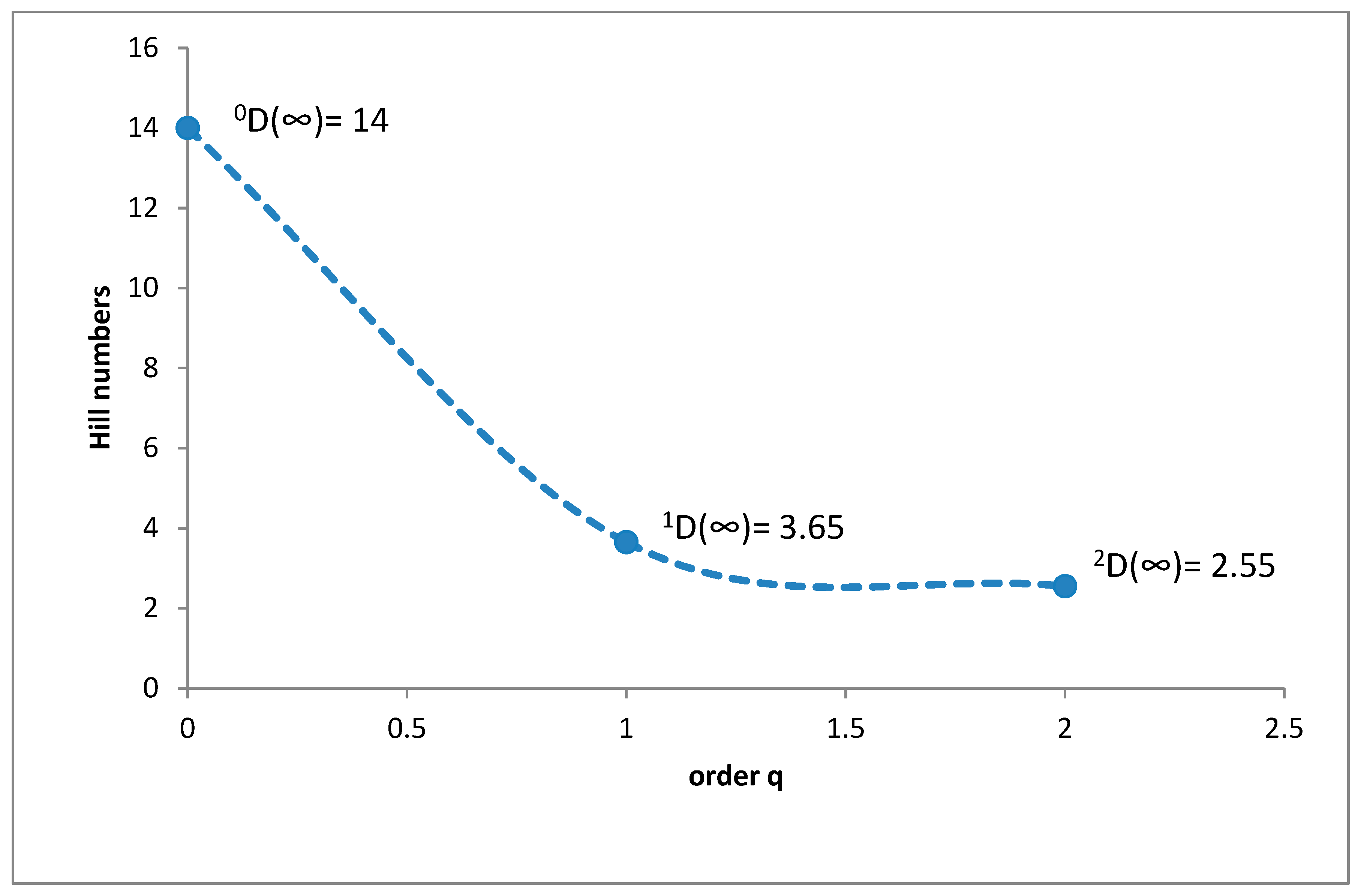
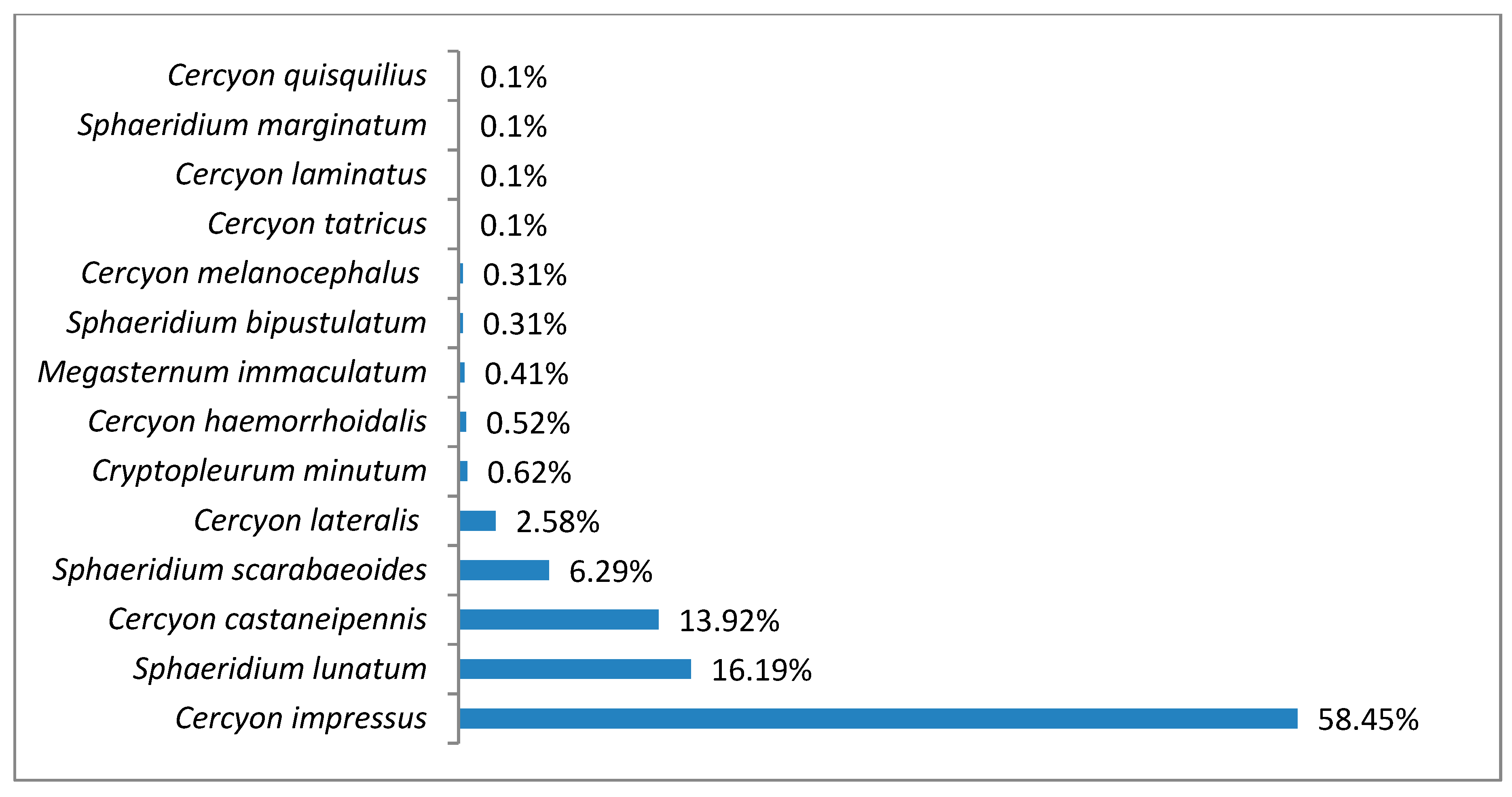
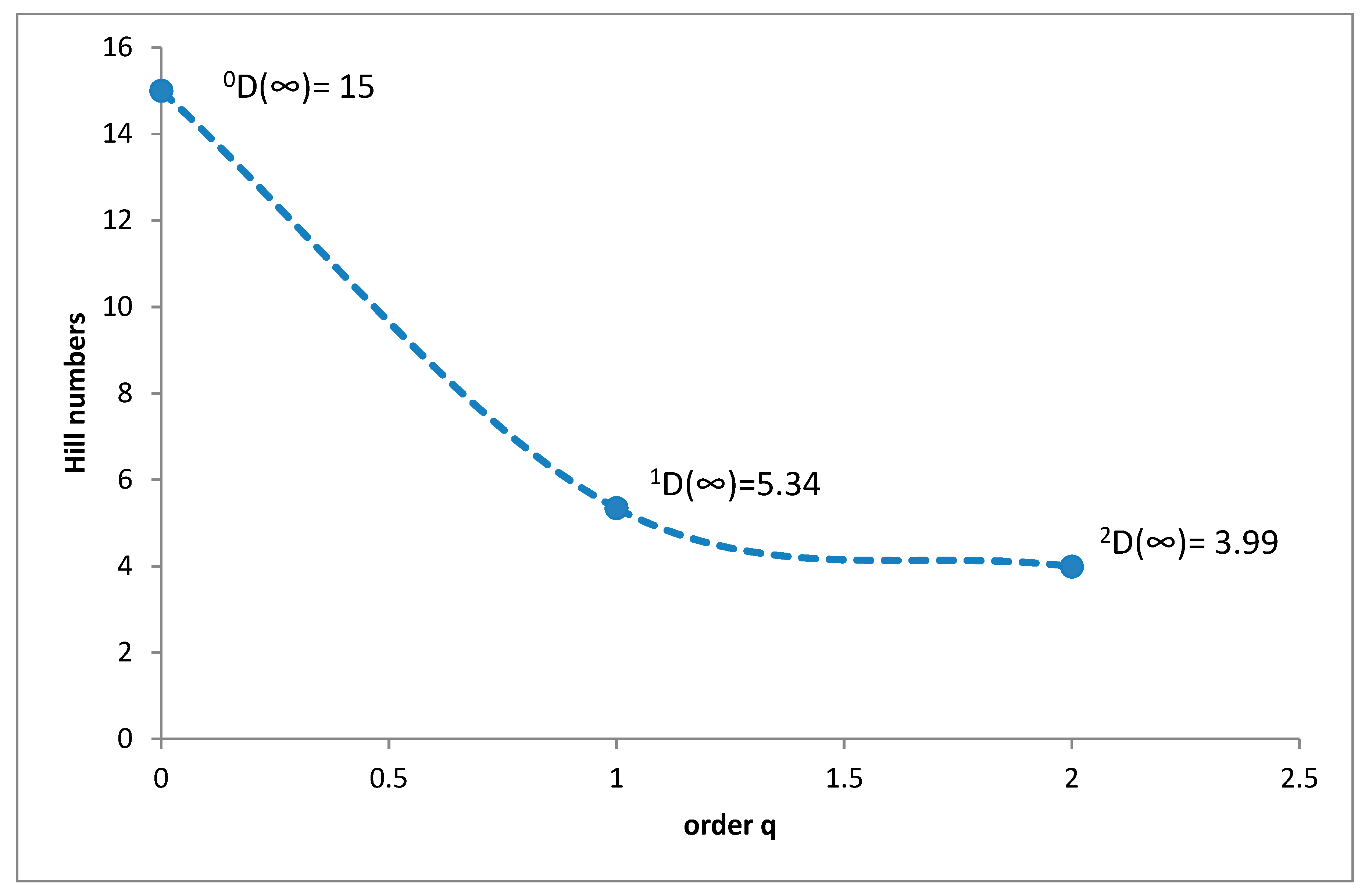
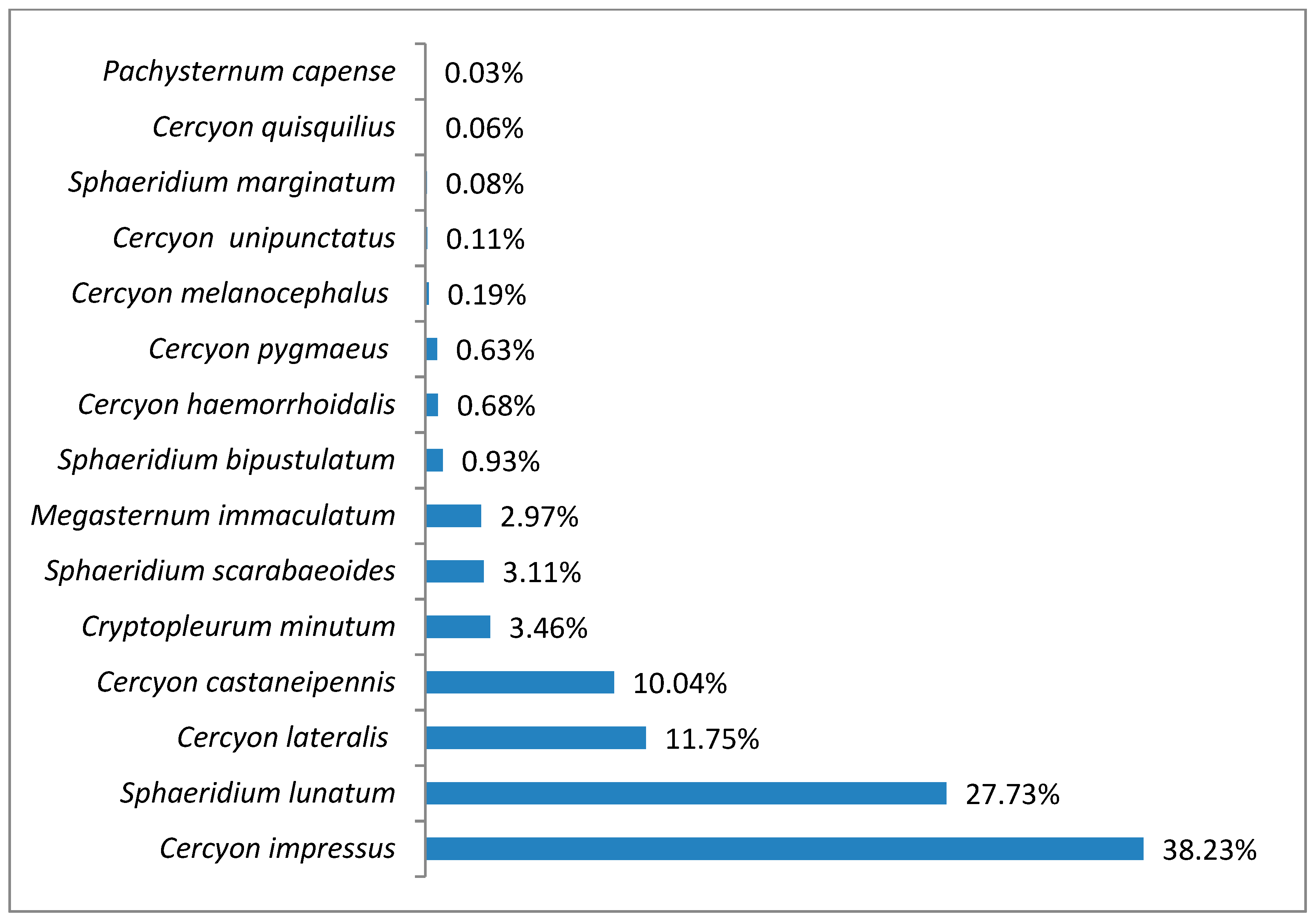
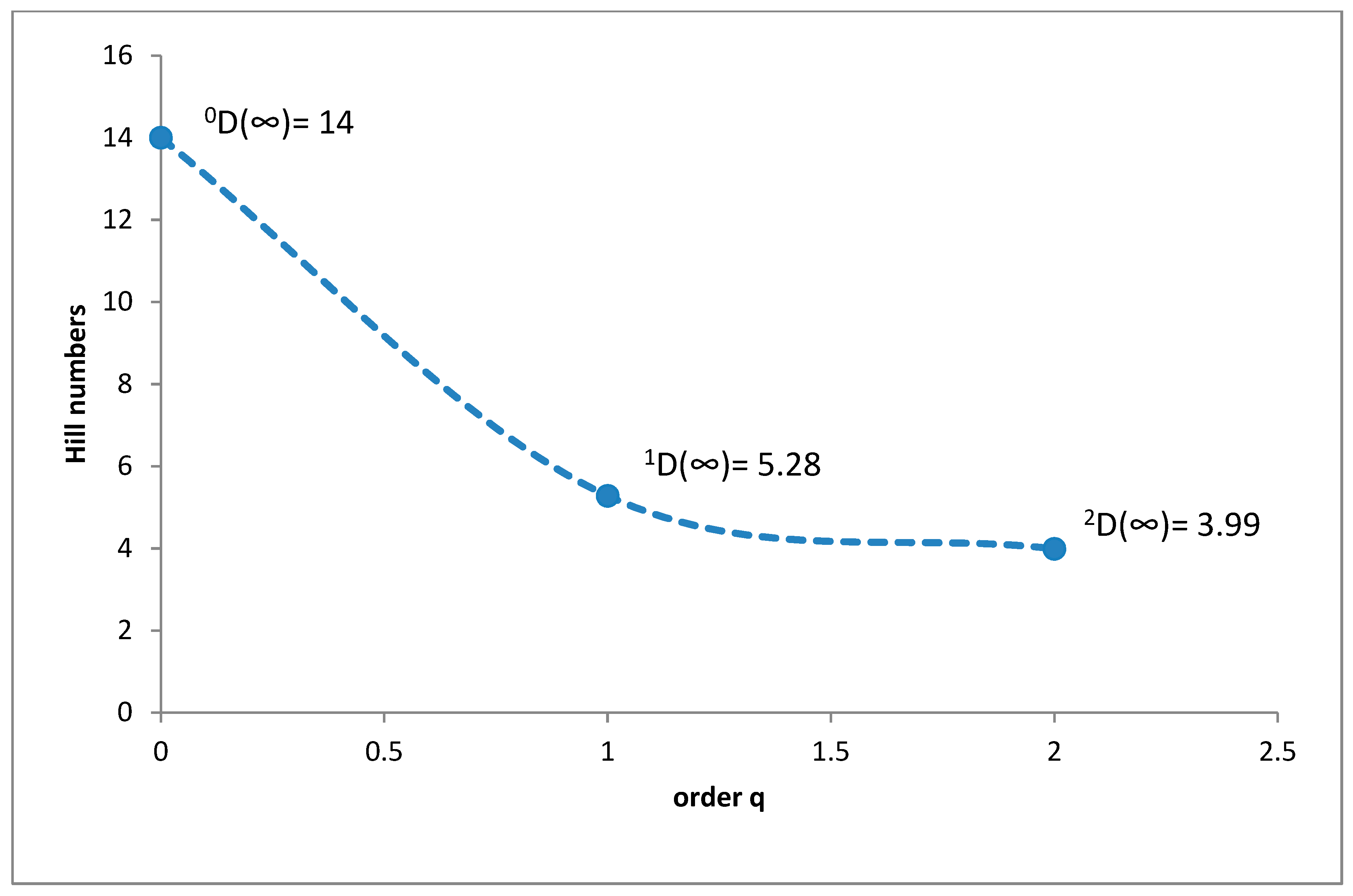
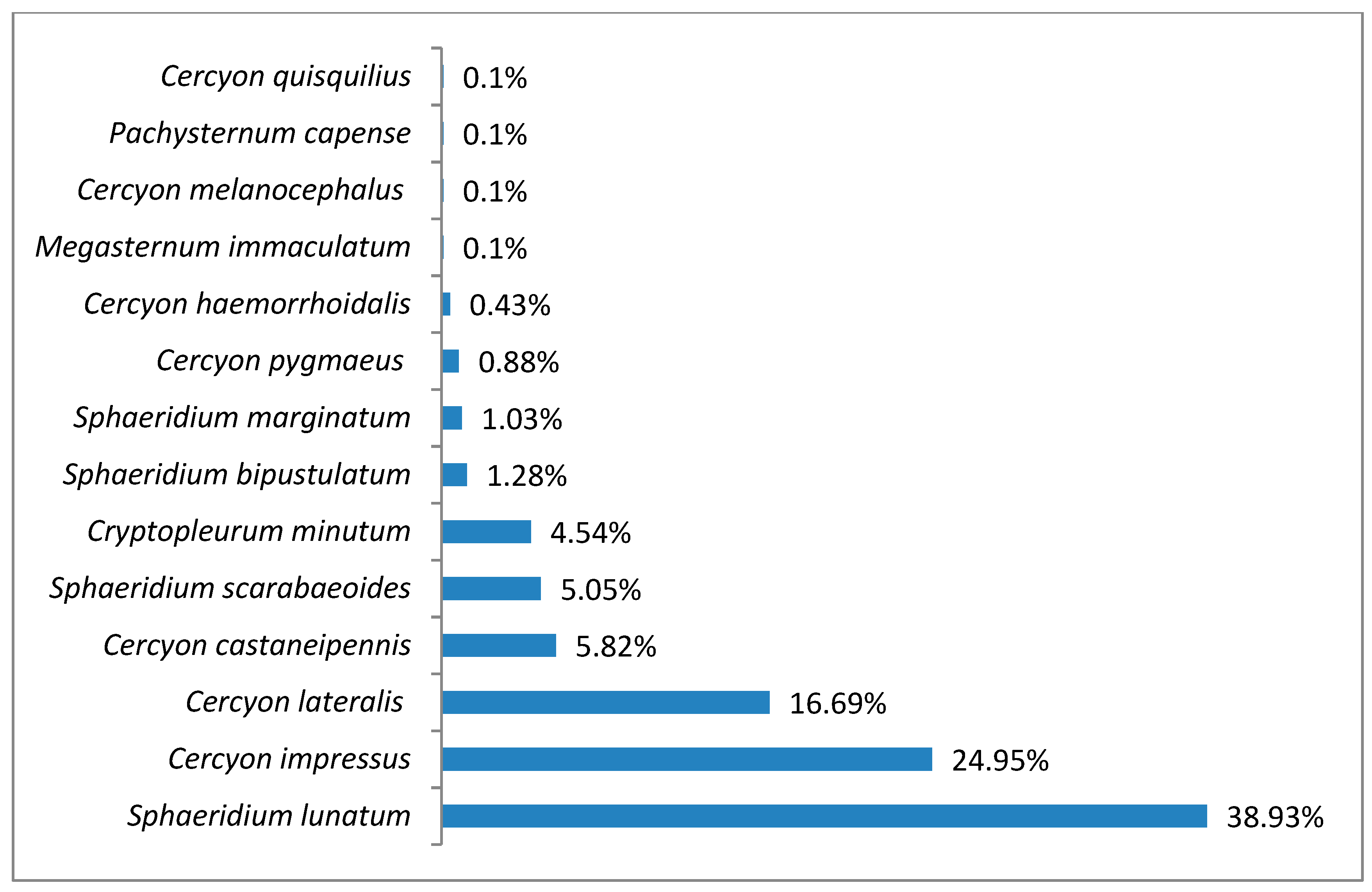
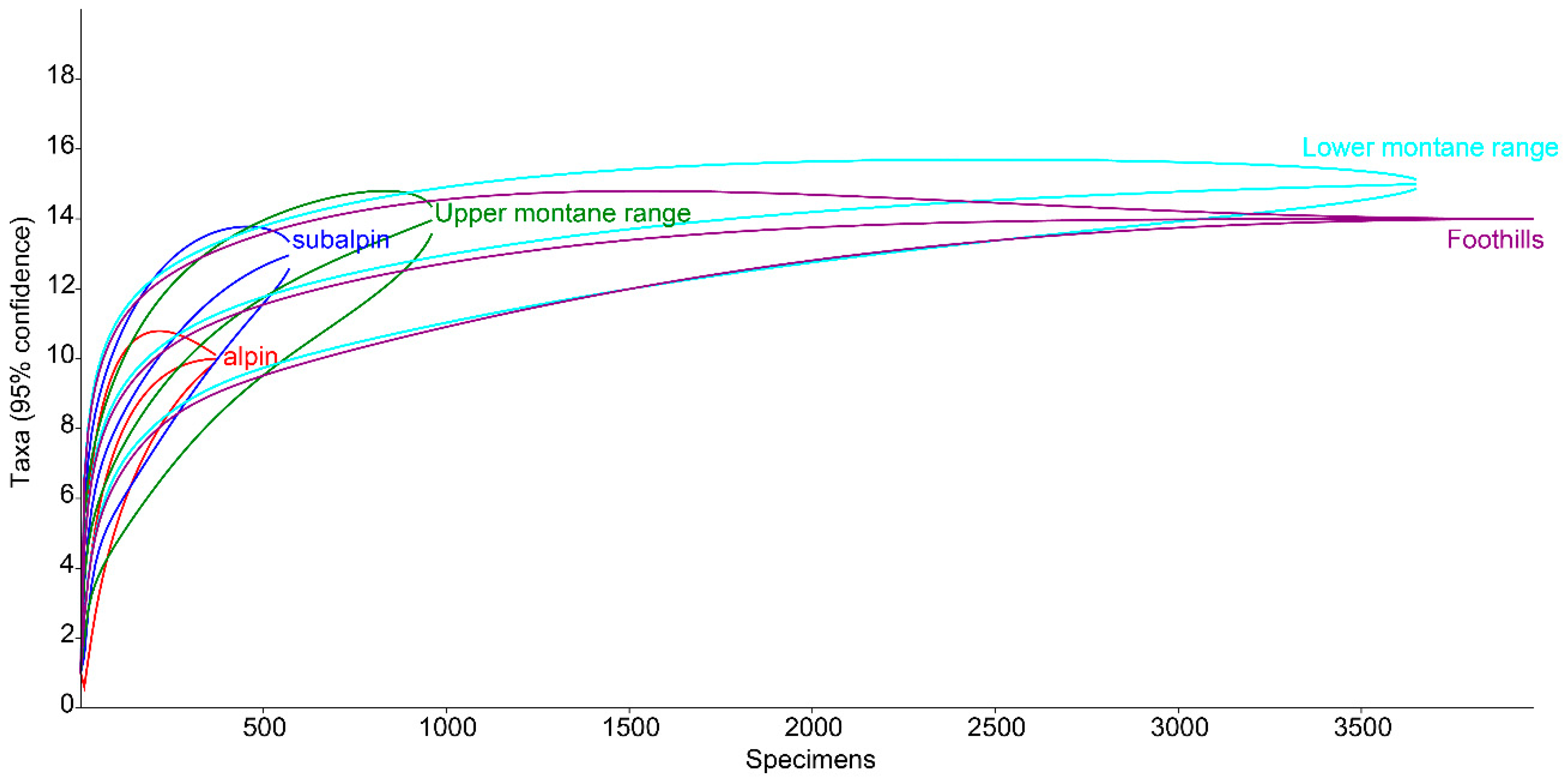
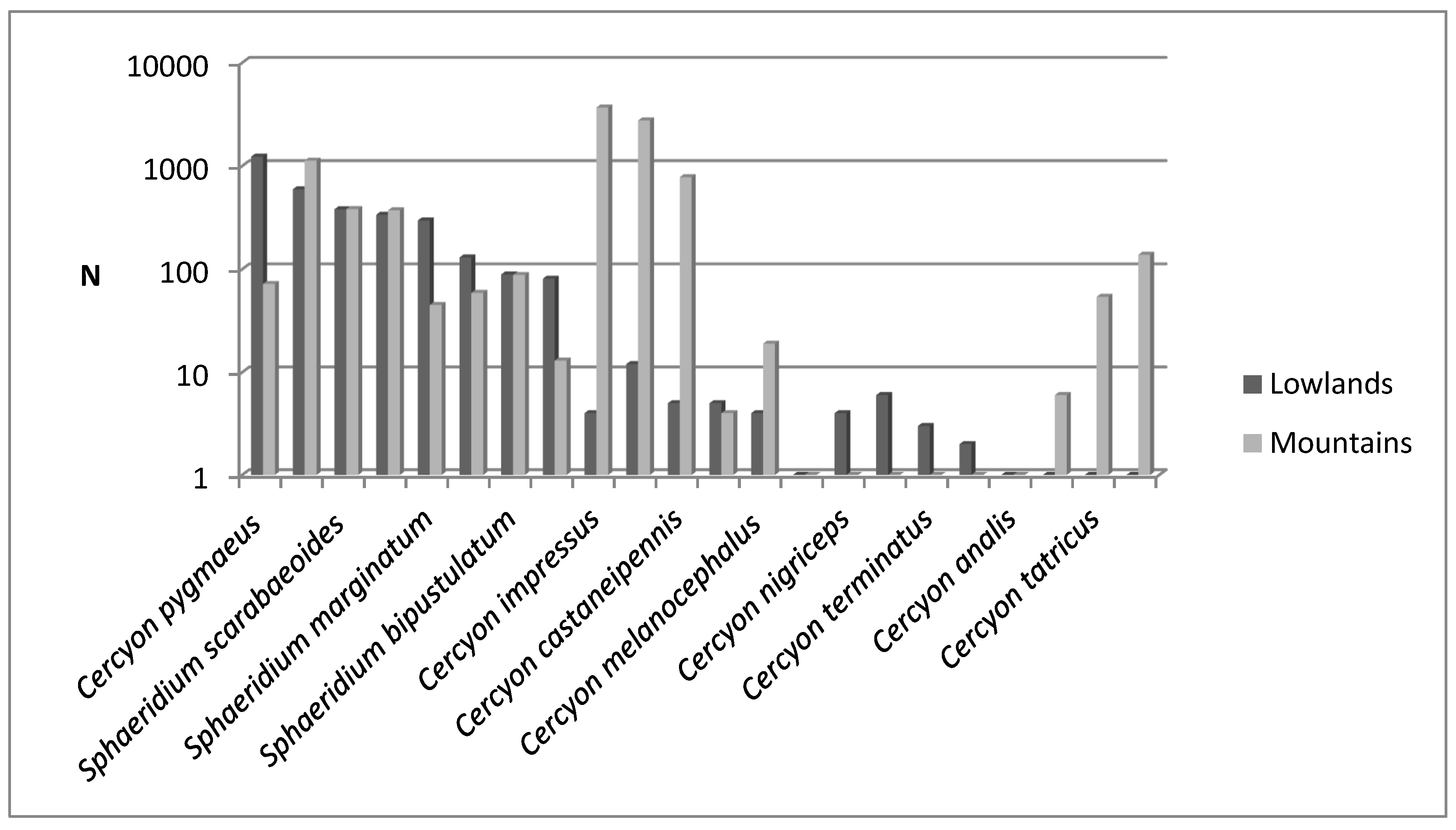
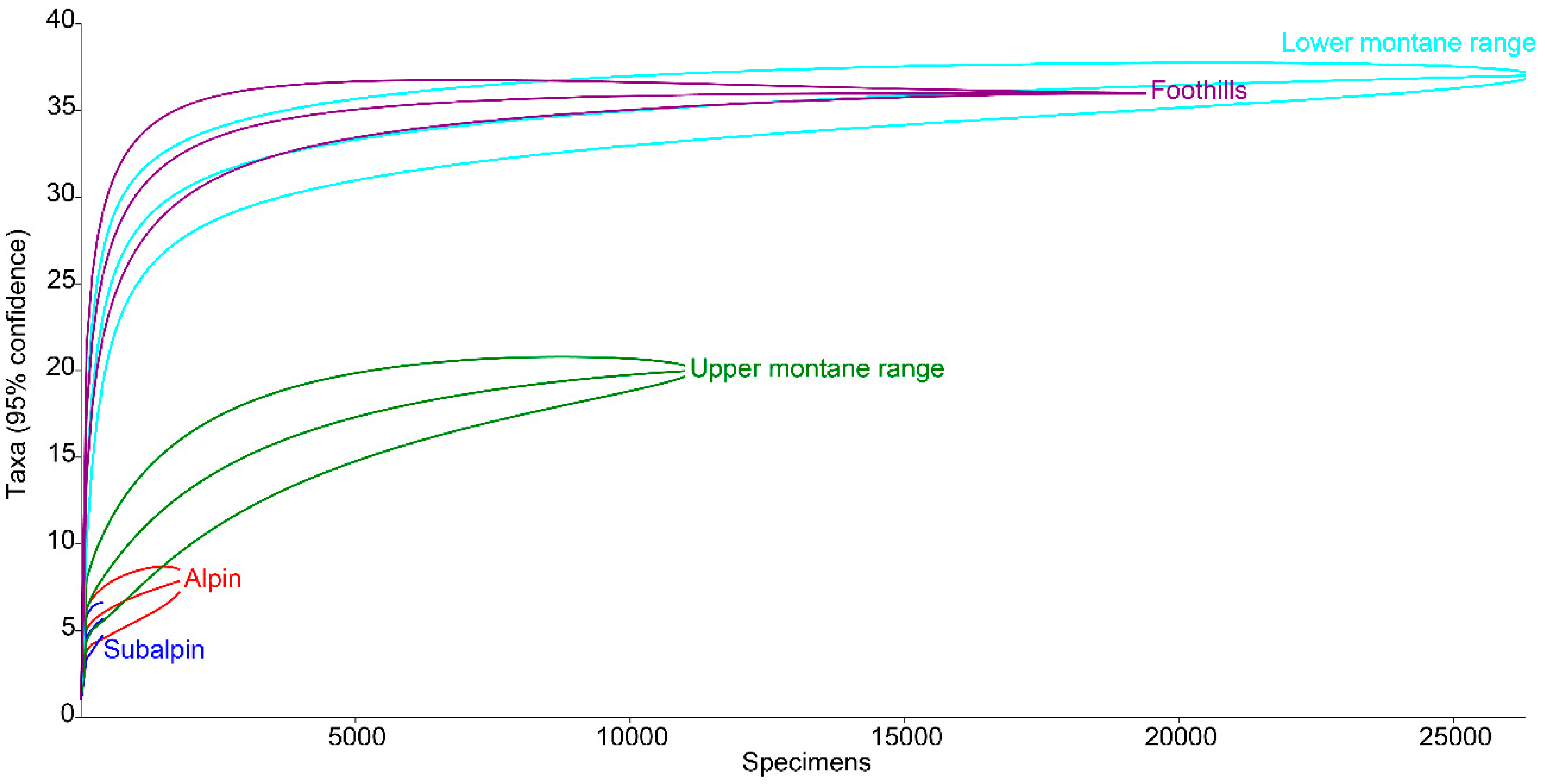
3.1. Species Diversity and Dominance Structures of Coprophagous Hydrophilid Beetle Assemblages in the Polish Carpathians
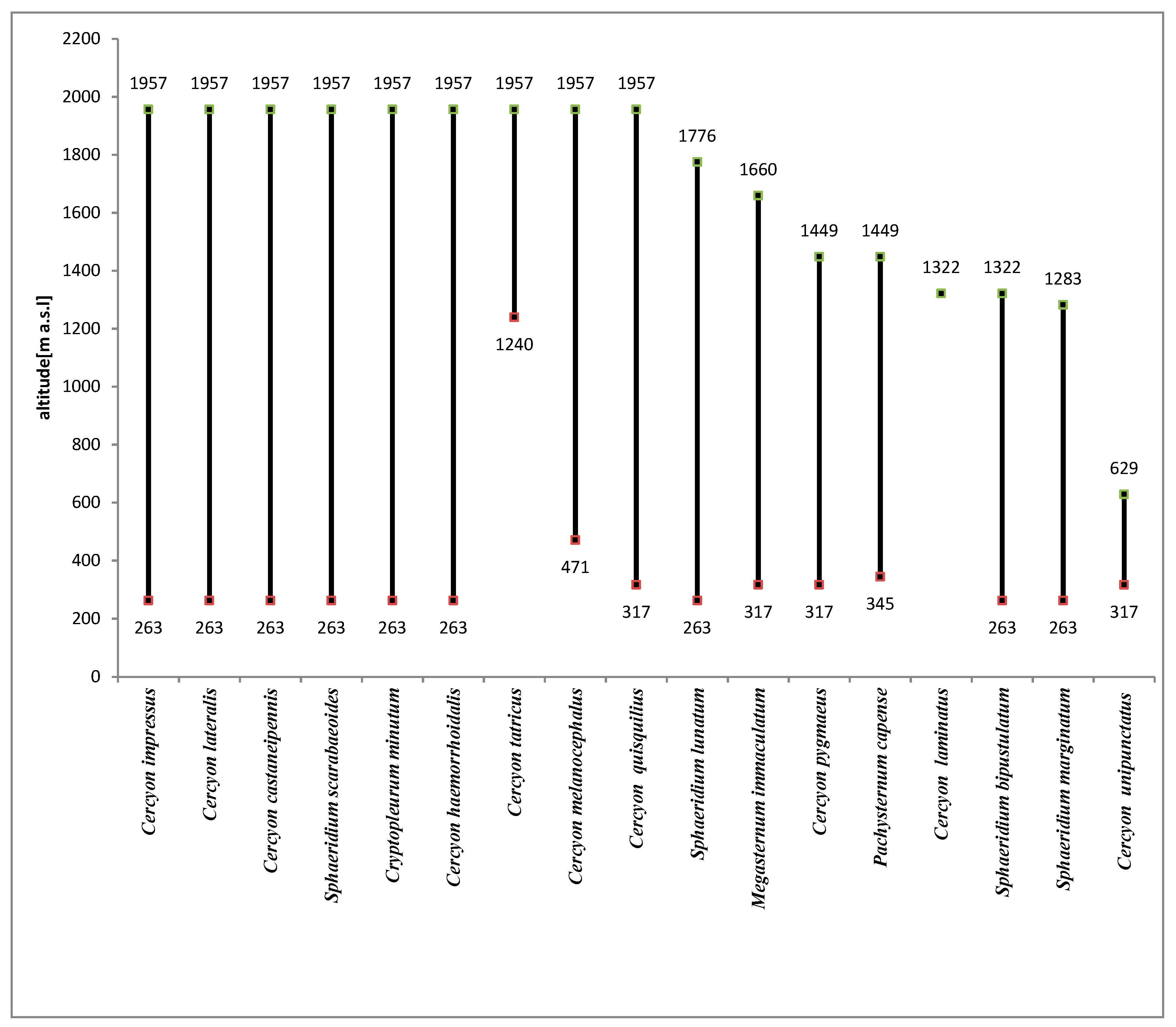
3.2. Altitudinal Distribution of Species in the Polish Carpathians
3.3. Overview of Collected Species
Cercyon (Cercyon) castaneipennis (Vorst, 2009)This recently described species has been recorded in Belarus, the Canary Islands, Russia, the Czech Republic, Latvia, the Netherlands, Poland, Slovakia and Sweden [34]. In the past, it was not distinguished from C. obsoletus (Gyllenhal, 1808), also known in Poland, although it was described as the color aberration C. obsoletus ab. rubridorsis by Reitter in the early twentieth century [35]. For this reason, its distribution in Poland is not yet well known. Some of the old data on C. obsoletus undoubtedly refer to this species. It has already been recorded in ten regions: the Baltic Coast, the Masurian Lake District, the Wielkopolska-Kujawska Lowland, the Mazovian Lowland, Białowieża Forest, Upper Silesia, the Kraków-Wieluń Upland, the Malopolska Upland, the Western Beskids and the Bieszczady Mountains [6,14,15,36,37]. It lives in the excrement of large herbivores (cows, horses and others) in diverse habitats [13].
Cercyon (Cercyon) haemorrhoidalis (Fabricius, 1775)This very widely distributed Palearctic species was also introduced to the Australian, Oriental, Nearctic and Neotropical regions [34,38]. It inhabits the feces of various herbivorous mammals as well as rotting plant debris, compost piles and carrion. It has also been found in the nests of birds and small rodents [1,38,39,40]. In Poland, it is widespread throughout the country, frequent and in places quite abundant. In the Polish Carpathians, it has been recorded in the Western Beskids, Eastern Beskids and the Bieszczady Mountains [41,42,43,44].It was found at 17 sites in the Polish Carpathians (Table 3). Its vertical range reaches the alpine belt, i.e., a minimum of about 2000 m a.s.l. It is new to the Tatra Mountains.
Cercyon (Cercyon) impressus (Sturm, 1807)This European species was introduced to North America [34]. It lives in all types of decaying plant and animal remains but prefers the excrement of herbivores, especially even-toed ungulates [40]. It is widespread throughout Poland. In the Polish Carpathians, it has been recorded in the Western Beskids, Eastern Beskids, Bieszczady Mountains and Tatra Mountains [36,41,42,43,44,45,46].Found at 45 sites in the Polish Carpathians (Table 3), it is the most abundant species of all those recorded (Table 2). Only within the foothills was it a dominant species (Figure 12), while in all other vegetation/climate belts it was a superdominant (Figure 4, Figure 6, Figure 8 and Figure 10). It was found at all altitude gradients in the study area (Figure 14).
Cercyon (Cercyon) lateralis (Marsham, 1802)This Palearctic species isvery widely distributed in nearly all of Europe and in the Russian part of Asia, as far as Kazakhstan and the Russian Far East. It was introduced to North America, where it became fully acclimated [34,47]. It is found in the excrement of horses, cows, European bison, deer and many other mammals, in rotting plant debris and in rotting fungi. It has also been found in the nests of birds: Turdus philomelos (C. L. Brehm) and Pernis apivorus (Linnaeus) [40,47]. It is widely distributed throughout Poland, where it is a common and abundant species. In the Polish Carpathians, it has been recorded in the Western Beskids, Eastern Beskids, Bieszczady Mountains and Tatra Mountains [36,41,42,43,44,45,46,48].
Cercyon (Cercyon) melanocephalus (Linnaeus, 1758)This species with Palearctic range is widely distributed in Europe and has been found in Asia in Lebanon, Russia (Western and Eastern Siberia) and Uzbekistan [34,49]. It lives in the excrement of herbivores, mainly even-toed ungulates [40]. In Poland, it is distributed throughout the country, but it is seen rarely and only as isolated specimens. It is known across the entire arc of the Polish Carpathians except the Pieniny Mountains [36,41,42,43,44,45,46].
Cercyon (Cercyon) pygmaeus (Illiger, 1801)This is a widely distributed Palearctic species, reaching Eastern Siberia. It has been introduced to North America [34]. It mainly inhabits the feces of herbivores, most often even-toed ungulates [40]. In Poland, it is widespread throughout the country. It is one of the most common coprophagous representatives of the genus. In the Polish Carpathians, it has been recorded in the Western Beskids, Eastern Beskids and Bieszczady Mountains [41,42,43,44,45,46].
Cercyon (Cercyon) quisquilius (Linnaeus, 1761)This species is widespread throughout the Palearctic region. It has been introduced to North America, South America and Australia [34,38,50]. It is polysaprophagous, inhabiting mainly the excrement of mammals (cows, horses, sheep and others). It is often found in compost, rotting fungi and plant waste [39,40,47,49,51,52,53]. It has also been caught in the nest of a black stork, Ciconia nigra (Linnaeus, 1758) [54]. It is widespread throughout Poland and has been recorded many times across the entire Polish Carpathians, except for the Tatra Mountains [36,41,42,43,44,55].
Cercyon (Cercyon) tatricus (Endródy–Younga 1967)This is a mountain species with a very interesting distribution. Until now, it had been recorded in two areas separated by a great distance—the Carpathians in Europe (Slovakia, Ukraine and Romania) and the Russian Far East (Amur Oblast, Khabarovsk Krai, Primorsky Krai and Kamchatka) [27,34]. It is found in the feces of even-toed ungulates, as well as that of bears [27,40]. It has been recorded in the High Tatras in Slovakia—the closest location to Poland—as well as the Belianske and Low Tatras [40]. It is also known in the Ivano-Frankivsk Oblast of Ukraine [27].
Cercyon (Cercyon) unipunctatus (Linnaeus, 1758)This is a widespread Palearctic species found all over Europe, except for its southernmost parts. In Asia, it is known in Kazakhstan, Russia, Mongolia, northern parts of China and Northern Japan [34,50]. In the mid-nineteenth century, it was introduced to North America, where it also spread rapidly [39]. It inhabits various types of decaying plant remains, mammal excrement (especially that of cows, horses and sheep) and chicken manure. It is sometimes found in decaying plant debris near water and in bird nests [54,56]. In Poland, it is widespread throughout the country, where it is very common and numerous. In the Polish Carpathians, it has been recorded many times in the Western Beskids, Eastern Beskids and the Bieszczady and Pieniny Mountains [41,43,44,55,57,58].
Cercyon (Paracycreon) laminatus (Sharp, 1873)This species was originally widespread in the Eastern Palearctic (Japan, Russian Far East and China). It was introduced to Europe in the mid-twentieth century, where it became fully acclimated [40]. It has now been found in Hawaii, Taiwan, Australia and Chile [38,59]. It lives in various kinds of decaying organic matter and the excrement of various mammals [40]. It is often encountered in Poland. Although it was first recorded in Poland relatively recently, it has already been found in 14 regions. In the Polish Carpathians, it is known in the Eastern Beskids and the Bieszczady and Pieniny Mountains [55,60,61,62].Only one individual was found in the Polish Carpathians (Table 2), on Tarnica in the Bieszczady Mountains (1322 m a.s.l.).
Pachysternum capense (Mulsant, 1844)This is a species originating in sub-Saharan Africa, from which it was introduced to Northern Africa, North America, South America, Australia, Europe and many islands including the Canary Islands, Madeira, the Comoros and Mauritius [63]. In Europe, it was first recorded on the basis of specimens caught in Greece in 1997 [24]. It has spread significantly since then and is currently known in France, Italy, Hungary and Romania [34,63,64]. This shows that this African species is fully acclimated in Europe and is rapidly expanding its range.A new species for Polish fauna. In the Polish Carpathians, six individuals were caught at five sites: Ciechań (629 m a.s.l.), Rozstajne (452 m a.s.l.), Stasiówka (395 m a.s.l.) and Zawada (384 m a.s.l.) in the Eastern Beskids and Upłaziańska Kopa (1449 m a.s.l.) in the Tatras (Table 3). The nearest known sites of this species in Romania and Hungary are more than 350 km away [63].
Megasternum immaculatum (Stephens, 1829)M. immaculatum, although described by Stephens almost 200 years ago, was until recently treated as a synonym of M. concinnum (Marsham, 1802) [65]. It was not restored to the status of a separate species until a few years ago [66]. Diagnostic characters provided by the authors, based on the structure of the male copulation apparatus and the upper body color, enable some distinction between the two species. Therefore, previously published data on the occurrence of M. concinnum require verification, since in many cases they probably refer to M. immaculatum. To date, it has been confirmed in the United Kingdom [6,34], Poland [6], Western and Eastern Siberia [67] and Bulgaria [68].Although the authors of the Catalogue of Palaearctic Coleoptera [34] explain in the comments that further research is underway to clarify the taxonomic questions regarding the genus Megasternum (associated with numerous synonyms attributed to M. concinnum), they do not question the presence of two distinct species in Central Europe.The species was found at 12 sites in the Polish Carpathians (Table 3). Its vertical range reached up to about 1660 m a.s.l.
Cryptopleurum minutum (Fabricius, 1775)This is a widely distributed Palearctic species, absent from North Africa, which was introduced to North America [34]. It lives mainly in animal excrement and is also often found in decaying plant debris [40]. It is common throughout Poland. It has been recorded many times over the entire arc of the Polish Carpathians [36,41,42,43,44].
Sphaeridium lunatum (Fabricius, 1792)This is a widely distributed Palearctic species that was introduced to North America [34]. It lives in the excrement of various herbivores, mainly even-toed ungulates [40]. It is common in Poland and probably distributed throughout the country. In the Polish Carpathians, it was previously recorded in the Western Beskids [69], Eastern Beskids [70] and the Bieszczady Mountains [43,58].It was caught at 38 sites in the Polish Carpathians (Table 3). It is the most numerous species of the genus and the second most numerous of all recorded representatives of Hydrophilidae (Table 2). It was a superdominant in the foothills (Figure 12), while within the lower montane range, upper montane range and subalpine belt, it belonged to the group of dominants (Figure 6, Figure 8 and Figure 10). Its vertical range reached up to 1800 m a.s.l. (Figure 14). It is new to the Tatras.
Sphaeridium scarabaeoides (Linnaeus, 1758)This is a Palearctic species that was introduced to Africa, Australia and North America [27]. It lives in the excrement of various herbivores, mainly even-toed ungulates [35]. In Poland, it is common and frequently encountered everywhere. In the Polish Carpathians, it had previously been recorded in the Western Beskids, Eastern Beskids and the Bieszczady and Tatra Mountains [36,38,39,53].
Sphaeridium bipustulatum (Fabricius, 1781)This is a widely distributed Palearctic species that was introduced to North America [34]. It lives in the excrement of various herbivores, mainly even-toed ungulates [40]. In Poland, it is common and frequently encountered. In the Polish Carpathians, it had previously been recorded in the Western Beskids, Eastern Beskids and the Bieszczady and Tatra Mountains [41,43,44,58]. It was caught at 15 sites within the Polish Carpathians (Table 3). Its vertical range reached up to 1322 m a.s.l. (Figure 14).
Sphaeridium marginatum (Fabricius, 1787)This is a widely distributed Palearctic species, also introduced to North America [34]. Like other European representatives of this genus, it lives in the excrements of various herbivores, mainly even-toed ungulates [40]. For a long time, it was treated as a variant of S. bipustulatum, until Van Berge–Henegouwen [26] demonstrated that it was a separate species. For this reason, its distribution in Poland is not yet well known. It has been recorded in five regions: the Wielkopolska-Kujawska Lowland [5,15,37,71,72,73], Podlasie [74], the Białowieża forest [75], Upper Silesia [6,36] and the Malopolska Upland [76,77].
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansen, M. The Hydrophiloidea (Coleoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1987, 18, 1–254. [Google Scholar]
- Hansen, M. The Hydrophiloid beetles, Phylogeny, Classification and a Revision of the Genera (Coleoptera, Hydrophiloidea). Biol. Skr. 1991, 40, 1–367. [Google Scholar]
- Archangelsky, M. Adaptations of immature stages of Sphaeridiinae (Staphyliniformia, Hydrophiloidea: Hydrophilidae) and state of knowledge of preimaginal Hydrophilidae. Coleopt. Bull. 1999, 53, 64–79. [Google Scholar]
- Fikáček, M. Hydrophilidae Leach, 1815. In Australian Beetles: Archostemata, Myxophaga, Adephaga, Polyphaga; Ślipiński, A., Lawrence, J., Eds.; CSIRO Publishing: Clayton, CA, USA, 2019; Volume 2, pp. 271–337. [Google Scholar]
- Przewoźny, M. Hydrophiloidea. In Fauna Polski—Charakterystyka i Wykaz Gatunków; Bogdanowicz, B., Chudzicka, E., Pilipiuk, I., Skibinska, E., Eds.; MiIZ PAN: Warszawa, Poland, 2004; pp. 149–151, 156–158. [Google Scholar]
- Greń, C.; Szołtys, S.; Grzywocz, J.; Królik, R. Chrząszcze (Coleoptera) Śląska Dolnego i Górnego—dotychczasowy stan poznania oraz nowe dane faunistyczne: Kałużnicowate (Hydrophilidae). Acta Ent. Sil. 2015, 23, 97–120. [Google Scholar]
- Hanski, I.; Koskela, H. Niche relations among dung—Inhabiting beetles. Oecologia 1977, 28, 203–231. [Google Scholar] [CrossRef]
- Hanski, I. The community of coprophagous beetles (Coleoptera, Scarabaeidae and Hydrophilidae) in northern Europe. Ann. Ent. Fenn. 1980, 46, 57–73. [Google Scholar]
- Hanski, I. The three coexisting species of Sphaeridium (Coleoptera, Hydrophilidae). Ann. Ent. Fenn. 1980, 46, 39–48. [Google Scholar]
- Hansen, M. De danske arter af slægten Sphaeridium Fabricius (Coleoptera, Hydrophilidae). Ent Meddr. 1990, 58, 59–64. [Google Scholar]
- Romero-Alcaraz, E.; Sánchez-Pinero, F.; Ávila, J.M. Los Sphaeridiinae (Coleóptera: Hydrophilidae) en una zona del suroeste ibérico. I: Composición faunística y fenología. Boln. Asoc. Esp. Ent. 1997, 21, 221–235. [Google Scholar]
- Hutton, S.A.; Giller, P.S. Intra- and interspecific aggregation of north temperate dung beetles on standardised and natural dung pads: The influence of spatial scale. Ecol. Entomol. 2004, 29, 594–605. [Google Scholar] [CrossRef]
- Bajerlein, D.; Przewoźny, M. Coprophagous hydrophilid beetles (Coleoptera: Hydrophilidae) as carriers of phoretic deutonymphs of Uropoda orbicularis (Acari: Mesostigmata) in Poland. Eur. J. Entomol. 2005, 102, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Vorst, O. Cercyon castaneipennis sp. n., an overlooked species from Europe (Coleoptera: Hydrophilidae). Zootaxa 2009, 2054, 59–68. [Google Scholar] [CrossRef]
- Przewoźny, M.; Bajerlein, D. The community of coprophagous hydrophilid beetles (Coleoptera: Hydrophilidae) in a pasture near Poznań (West Wielkopolska, Poland). Pol. J. Entomol. 2010, 79, 253–260. [Google Scholar]
- Mroczyński, R.; Radoslav, K. Differences between beetle communities colonizing cattle and horse dung. Eur. J. Entomol. 2014, 111, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Mroczyński, R.; Marczak, D. Coprophagous beetles (Coleoptera) found in moose (Alces alces L.) feces in Kampinos National Park. WSN 2017, 86, 376–381. [Google Scholar]
- Šlachta, M.; Frelich, J.; Svoboda, L. Seasonal biomass distribution of dung beetles (Scarabaeidae, Geotrupidae, Hydrophilidae) in mountain pastures of South-West Bohemia. J. Agrobiol. 2008, 25, 163–176. [Google Scholar]
- Šlachta, M.; Frelich, J.; Tonka, T. Application of dung-baited pitfall trapping in monitoring study on diversity of coprophagous beetles (Scarabaeidae, Geotrupidae, Hydrophilidae) in cattle pastures. J. Agrobiol. 2009, 26, 83–99. [Google Scholar]
- Šlachta, M.; Frelich, J.; Tonka, T. Composition of community of coprophagous beetles (Coleoptera: Scarabaeidae, Geotrupidae, Hydrophilidae) in beef cattle pasture in Western Bohemia—An application of dung-baited pitfall trapping in monitoring study on coprophagous fauna. Erica 2009, 16, 97–112. [Google Scholar]
- Šlachta, M. Coprophagous beetle community (Coleoptera: Scarabaeidae, Geotrupidae, Hydrophilidae) in two cattle pastures in South Bohemia. J. Agrobiol. 2013, 30, 21–31. [Google Scholar] [CrossRef]
- Górz, A. Dung Beetles of the Polish Carpathians; Pedagogical University Press: Krakow, Poland, 2019; p. 134. [Google Scholar]
- Przewoźny, M. Catalogue of Palearctic Hydrophiloidea (Coleoptera); Internet Version 2019-01-01. Available online: http://waterbeetles.eu/ (accessed on 17 November 2019).
- Fikáček, M.; Boukal, M. Pachysternum capense, a new genus and species for Europe, and updated key to genera and subgenera of European Sphaeridiinae (Coleoptera: Hydrophilidae). Klapalekiana 2004, 40, 1–12. [Google Scholar]
- Freude, H.; Harde, K.W.; Lohse, G.A. Die Käfer Mitteleuropas; Goecke & Evers: Krefeld, Germany, 1971; p. 365. [Google Scholar]
- Van Berge Henegouwen, A. Sphaeridium marginatum reinstated as a species distinct from S. bipustulatum (Coleoptera: Hydrophilidae). Ent. Ber. Amst. 1989, 49, 168–170. [Google Scholar]
- Fikáček, M. Taxonomic status of Cercyon alpinus, C. exorabilis, C. strandi and C. tatricus and notes on their biology (Coleoptera: Hydrophilidae: Sphaeridiinae). Ann. Naturhist. Mus. Wien 2006, 107B, 145–164. [Google Scholar]
- Hortal, J.; Paulo, A.V.; Borges, P.A.; Gaspar, C. Evaluating the performance of species richness estimators: Sensitivity to sample grain size. J. Anim. Ecol. 2006, 75, 274–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotelli, N.J.; Colwell, R.K. Estimating species richness. In Biological Diversity: Frontiers in Measurement and Assessment; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 39–54. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [Green Version]
- Fikáček, M.; Angus, R.B.; Gentili, E.; Jia, F.; Minoshima, Y.N.; Prokin, A.; Przewoźny, M.; Ryndevich, S.K. Hydrophilidae. In Catalogue of Palaearctic Coleoptera. Hydrophiloidea—Staphylinoidea, Revised and Updated Edition; Löbl, I., Löbl, D., Eds.; Koninklijke Brill NV: Leiden, The Netherlands; Boston, MA, USA, 2015; Volume 2, pp. 37–76. [Google Scholar]
- Reitter, E. Fauna Germanica. Die Käfer des Deutschen Reiches; II. Band. Schr. Dtsch. Lehrerver. Naturk., 24: Stuttgart, Germany, 1909; p. 392. [Google Scholar]
- Greń, C. Chrząszcze z rodzin Hydrochidae i Hydrophilidae (Coleoptera) w zbiorach Muzeum Górnośląskiego w Bytomiu. Acta Ent. Sil. 2011, 19, 55–69. [Google Scholar]
- Bajerlein, D.; Przewoźny, M. When a beetle is too small to carry phoretic mites? A case of hydrophilid beetles (Coleoptera: Hydrophilidae) and Uropoda orbicularis (acari: Mesostigmata). Can. J. Zool. 2012, 90, 368–375. [Google Scholar] [CrossRef]
- Fikáček, M. Occurrence of introduced species of the genus Cercyon (Coleoptera: Hydrophilidae) in the Neotropical Region. Rev. de la Soc. Entomol. Argent. 2009, 68, 351–357. [Google Scholar]
- Smetana, A. Revision of the subfamily Sphaeridiinae of America north of Mexico (Coleoptera: Hydrophilidae). Mem. Entomol. Soc. Can. 1978, 105, 1–292. [Google Scholar] [CrossRef]
- Boukal, D.S.; Boukal, M.; Fikáček, M.; Hájek, J.; Klečka, J.; Skalický, S.; Šťasný, J.; Trávníček, D. Catalogue of water beetles of the Czech Republic. Klapalekiana 2007, 43, 1–289. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Adephaga prócz Carabidae, Myxophaga, Polyphaga: Hydrophiloidea; Katalog Fauny Polski; PWN: Warszawa, Poland, 1976; pp. 1–307. [Google Scholar]
- Borowiec, L.; Kania, J. Chrząszcze (Coleoptera) nowe i rzadkie w faunie Bieszczadów. Wiad. Entomol. 1995, 14, 153–157. [Google Scholar]
- Pawłowski, J.; Petryszak, B.; Kubisz, D.; Szwałko, P. Chrząszcze (Coleoptera) Bieszczadów Zachodnich; Monografie Bieszczadzkie BPN: Ustrzyki Dolne, Poland, 2000; Volume 8, pp. 9–143. [Google Scholar]
- Kubisz, D.; Szafraniec, S. Chrząszcze (Coleoptera) masywu Babiej Góry. In Monografia Fauny Babiej Góry; Wołoszyn, B.W., Wołoszyn, D., Celary, W., Eds.; Komitet Ochrony Przyrody PAN: Kraków, Poland, 2003; pp. 163–221. [Google Scholar]
- Greń, C. Chrząszcze wodne (Coleoptera: Dytiscidae, Haliplidae, Hydrophilidae, Elmidae) obszaru źródliskowego Wisły w rezerwacie przyrody „Barania Góra“ (Beskid Śląski). Acta Ent. Sil. 2009, 17, 41–52. [Google Scholar]
- Wojas, T. Materiały do poznania chrząszczy (Insecta: Coleoptera) torfowisk i młak Tatr Polskich. Parki Nar. Rez. Przyr. 2010, 29, 49–75. [Google Scholar]
- Ryndevich, S.K. Review of species of the genus Cercyon Leach, 1817 of Russia and adjacent regions. I. Subgenus Cercyon (s. str.) Leach, 1817. Cercyon lateralis—Group (Coleoptera: Hydrophilidae). Annales Universitatis Mariae Curie-Sklodowska (Section C) 2004, 59, 1–13. [Google Scholar]
- Kubisz, D.; Szafraniec, S. Interesujące gatunki chrząszczy stwierdzone w masywie Babiej Góry, Beskid Zachodni (Coleoptera). Acta Ent. Sil. 2001, 7, 43–48. [Google Scholar]
- Ryndevich, S.K.; Fikáček, M. Faunistic and Zoogeographic Notes on Hydrophiloid Beetles from the Palaearctic Region (Coleoptera: Hydrophilidae). Vestn. BarDU Seriya Biol. Nauk. Selskokhozyaistvennye Nauk. 2013, 1, 32–37. [Google Scholar]
- Ryndevich, S.K.; Jia, F.; Fikáček, M. A review of the Asian species of the Cercyon unipunctatus group (Coleoptera: Hydrophilidae: Sphaeridiinae). Entomol. Musei Natl. Pragae 2017, 57, 535–576. [Google Scholar] [CrossRef] [Green Version]
- Smetana, A. Review of the family Hydrophilidae of Canada and Alaska (Coleoptera). Mem. Entomol. Soc. Can. 1988, 142, 3–316. [Google Scholar] [CrossRef]
- Nikitsky, N.B.; Osipov, I.N.; Chemeris, M.V.; Semenov, V.B.; Gusakov, A.A. The beetles of Prioksko-terrasny biosphere reserve—Xylobiontes, mycetobiontes, and Scarabaeidae (with review of the Moscow region fauna of the groups). Arch. Zool. Mus. Mosc. State Univ. 1996, 36, 1–197. [Google Scholar]
- Ryndevich, S.K. Ecological classification on the basis of ecological preferences for the genus Cercyon Leach, 1817 (Coleoptera: Hydrophilidae) of the Palaearctic region. In Materials of the Third All-Russian Symposium on Amphibiotic and Aquatic Insects; Publishing and Printing Center of Voronezh State University: Voronezh, Russia, 2007; pp. 281–284. [Google Scholar]
- Lundyshev, D.S.; Ryndevich, S.K. Water beetles (Coleoptera: Dytiscidae, Noteridae, Helophoridae, Spercheidae, Hydrophilidae, Hydraenidae, Dryopidae) and hydrophilids (Hydrophilidae) in birds nests of Belarus. Vesnik Grodzenskaga Dzyarzhaunaga Universiteta Imya Yanki Kupaly. Seryya 2. Matematyka, Fizika. Infarmatyka, Vylichalnaya Tekhnika i Kiravanne. Biyalogiya 2010, 3, 107–116. [Google Scholar]
- Rossa, R. Materiały do poznania chrząszczy (Coleoptera) Pienińskiego Parku Narodowego. Wiad. Entomol. 1999, 17, 193–194. [Google Scholar]
- Ryndevich, S.K.; Lundyshev, D.S. Beetles of birds nests (Coleoptera: Noteridae, Dytiscidae, Helophoridae, Hydrophilidae & Dryopidae). Latissimus 2005, 20, 17–19. [Google Scholar]
- Galewski, K. Chrząszcze (Coleoptera) Rodzina: Kałużnicowate (Hydrophilidae). Fauna Słodkowodna Polski; PWN: Warszawa, Poland, 1990; p. 261. [Google Scholar]
- Kubisz, D.; Szwałko, P.; Wojas, T. Materials to the fauna of Coleoptera of the Western Bieszczady Mts. (Polisch Eastern Carpathians). Roczn. Muz. górnośl. (Przyr.) 1998, 15, 5–15. [Google Scholar]
- Ryndevich, S.K. Review of species of the genus Cercyon Leach, 1817 of Russia and adjacent regions. IV. The subgenera Paracycreon Orchymont, 1924 and Dicytocercyon Ganglbauer, 1904 (Coleoptera: Hydrophilidae). Zoosystematica Ross. 2008, 17, 89–97. [Google Scholar]
- Kubisz, D.; Szwałko, P. Nowe stanowiska Cercyon laminatus Sharp, Berosus bispina Reiche et Saulcy (Coleoptera, Hydrophilidae). Wiad. Entomol. 1991, 10, 128. [Google Scholar]
- Greń, C. Nowe stanowiska polskich Hydrophilidae (Coleoptera). Acta Ent. Sil. 2003, 9–10, 86. [Google Scholar]
- Przewoźny, M. Nowe stanowiska kałużnic (Coleoptera: Hydrophiloidea) w Polsce. Wiad. Entomol. 2004, 23, 69–80. [Google Scholar]
- Lőkkös, A.; Boukal, M.; Fikáček, M. The hydrophilid beetle Pachysternum capense (Mulsant, 1844) (Coleoptera: Hydrophilidae) became a world-wide adventive species: A summary of distribution with new records from Europe, Australia, and South America. North-West. J. Zool. 2014, 10, 333–336. [Google Scholar]
- Toledo, M.; Rocchi, S. Reperti inediti di Coleotteri acquatici in Italia (Coleoptera: Hydroscaphidae, Gyrinidae, Dytiscidae, Helophoridae, Hydrophilidae, Hydraenidae, Limnichidae, Erirhinidae). Onychium 2017, 13, 63–74. [Google Scholar]
- Hansen, M. Hydrophilidae. In Catalogue of Palaearctic Coleoptera. Hydrophiloidea—Histeroidea—Staphylinoidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2004; Volume 2, pp. 44–68. [Google Scholar]
- Foster, G.N.; Bilton, D.T.; Friday, L.E. Keys to adults of the water beetles of Britain and Ireland; Volume 4 part 5b; RES Handbooks for the Identification of British Insects: St Albans, UK, 2014; p. 126. [Google Scholar]
- Ryndevich, S.K. New faunistic records of hydrophilid beetles (Coleoptera: Hydrophiloidea: Hydrophilidae) from Eurasia. BarSU Herald. Ser. Biol. Sci. Gen. Biol. Agric. Sci. Agron. 2017, 5, 65–70. [Google Scholar]
- Greń, C.; Lubecki, K. Water beetles (Coleoptera: Adephaga, Hydrophiloidea, Byrrhoidea) in Bulgaria: New records. Ann. Up. Sil. Mus. Bytom Entomol. 2018, 26, 1–20. [Google Scholar]
- Wachtl, F. Spis chrząszczów z dorzecza Soły i Koszarawy. Spraw. Kom. Fizyogr. 1870, 4, 246–262. [Google Scholar]
- Trella, T. Wykaz chrząszczów okolic Przemyśla. Hydrophilidae, Lucanidae, Scarabaeidae. Pol. Pismo Ent. 1937, 14–15, 262–265. [Google Scholar]
- Myrdzik, K. Matriały do fauny chrząszczy lądowych województwa Poznańskiego. Pr. Kom. Mt.-Przyr. PTPN B Poznań 1933, 6, 173–194. [Google Scholar]
- Przewoźny, M. Kałużnice (Coleoptera: Hydrophilidae) w zbiorze Kazimierza Myrdzika. Bad. Fizjogr. Pol. Zach. Ser. C Poznań 2004, 50, 67–69. [Google Scholar]
- Ruta, R.; Orzechowski, R.; Aleksandrowicz, O.; Borowski, J.; Buchholz, L.; Komosiński, K.; Lubecki, K. Przewoźny M. Chrząszcze (Insecta: Coleoptera) Gryżyńskiego Parku Krajobrazowego. Przegl. Przyr. 2016, 27, 28–62. [Google Scholar]
- Przewoźny, M.; Lasoń, A. Nowe dla Podlasia i Wyżyny Lubelskiej kałużnice (Coleoptera: Hyfrophilidae). Wiad. Entomol. 2003, 22, 60–61. [Google Scholar]
- Borowiec, L.; Kania, J.; Wanat, M. Chrząszcze (Coleoptera) nowe dla Puszczy Białowieskiej. Wiad. Entomol. 1992, 11, 133–141. [Google Scholar]
- Przewoźny, M.; Miłkowski, M. Kałużnice (Coleoptera: Hydrophiloidea) i Hydraenidae (Coleoptera: Staphylinoidea) nowe dla Wyżyny Małopolskiej. Wiad. Entomol. 2004, 23, 157–162. [Google Scholar]
- Jaskuła, R.; Przewoźny, M.; Melke, A. Chrząszcze (Coleoptera). In Owady (Insecta) Spalskiego Parku Krajobrazowego Część I; Jaskuła, R., Tończyk, G., Eds.; Mazowiecko-Świętokrzyskie Towarzystwo Ornitologiczne: Spała, Poland, 2009; pp. 27–59. [Google Scholar]
- Weiner, J. Życie i Ewolucja Biosfery; PWN: Warszawa, Poland, 1999; p. 591. [Google Scholar]
- Brown, J. Species diversity. In Analytical Biogeography: An Integrated Approach the Stud" of Animal and Plant Distributions; Myers, A.A., Giller, P.S., Eds.; Chapman and Hall: London, UK, 1988; pp. 57–89. [Google Scholar]
- McCoy, D.E. The Distribution of Insects along Elevational Gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Pawłowski, J. Cenne bezkręgowce naziemne Magurskiego Parku Narodowego i terenów ościennych. In Magurski Park Narodowy—Monografia przyrodnicza; Górecki, A., Zemanek, B., Eds.; MPN: Krempna-Kraków, Poland, 2009; pp. 132–146. [Google Scholar]
- Wainwright, C.E.; Stepanian, P.M.; Reynolds, D.R.; Reynolds, A.R. The movement of small insects in the convective boundary layer: Linking patterns to processes. Sci. Rep. 2017, 7, 5438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlicz, M. Klimat Tatr. In Tatrzański Park Narodowy; Szafer, W., Ed.; PAN: Kraków, Poland, 1962; pp. 15–71. [Google Scholar]
- Fenton, M.B. Feeding ecology of insectivorous bats. Bios 1974, 45, 3–15. [Google Scholar]
- Piksa, K.; Brzuskowski, T.; Cichocki, J.; Gubała, W. Species diversity of bats Chiroptera in the Tatra National Park during the summer activity period. Chrońmy Przyr. Ojcz. 2017, 73, 121–134. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
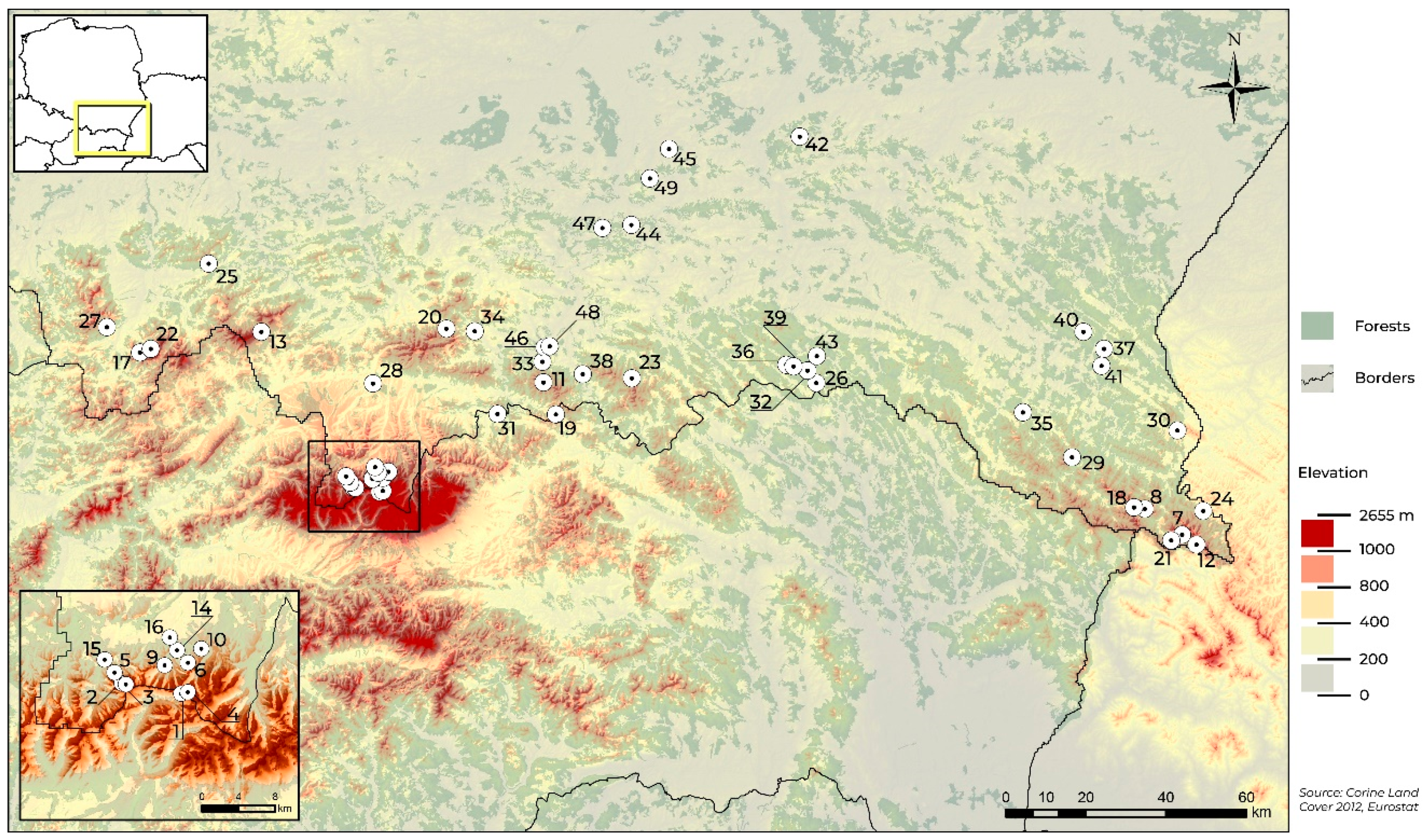
| No. on Map | Site | Elevation (m a.s.l.) | Geographical Coordinate System in DD (Decimal Degrees) | Vegetation Belt | Type of Plant Association |
|---|---|---|---|---|---|
| 1 | Kasprowy Wierch | 1957 | N 49.22850° E 019.98712° | Alpine | Trifido-Distichetum |
| 2 | Twarda Galeria | 1776 | N 49.23967° E 019.89746° | Alpine | Trifido-Distichetum |
| 3 | Kocioł Mułowy | 1707 | N 49.23848° E 019.90410° | Alpine | Trifido-Distichetum |
| 4 | Hala Gąsienicowa | 1660 | N 49.23026° E 019.99769° | Subalpine | Trifido-Distichetum |
| 5 | Upłaziańska Kopa | 1449 | N 49.25033° E 019.88700° | Subalpine | Gladiolo-Agrostietum |
| 6 | Skupniów Upłaz | 1390 | N 49.25930° E 019.99849° | Upper montane range | Gladiolo-Agrostietum |
| 7 | Tarnica | 1322 | N 49.07629° E 022.72537° | Subalpine meadows (Poloninas) | Poo-Deschampsietum |
| 8 | Połonina Caryńska | 1283 | N 49.13834° E 022.60265° | Subalpine meadows (Poloninas) | Poo-Deschampsietum |
| 9 | Hala Kondratowa | 1240 | N 49.25715° E 019.96276° | Upper montane range | Gladiolo-Agrostietum |
| 10 | Polana Kopieniec | 1236 | N 49.27299° E 020.01854° | Upper montane range | Gladiolo-Agrostietum |
| 11 | Przehyba | 1133 | N 49.46750° E 020.55543° | Upper montane range | Plagiothecio-Piceetum |
| 12 | Przełęcz Bukowska | 1117 | N 49.05225° E 022.77288° | Upper montane range | Poo-Deschampsietum |
| 13 | Przełęcz Krowiarki | 1058 | N 45.58989° E 019.58641° | Lower montane range | Plagiothecio-Piceetum |
| 14 | Kuźnice | 1000 | N 49.27168° E 019.98218° | Lower montane range | Arrhenatheretum alatioris |
| 15 | Dolina Kościeliska | 980 | N 49.26355° E 019.87252° | Lower montane range | Gladiolo-Agrostietum |
| 16 | Zakopane TPN | 902 | N 49.28448° E 019.97127° | Lower montane range | Gladiolo-Agrostietum |
| 17 | Hala Boracza | 863 | N 49.54587° E 019.16575° | Lower montane range | Gladiolo-Agrostietum |
| 18 | Brzegi Górne | 773 | N 49.14267° E 022.56499° | Lower montane range | Gladiolo-Agrostietum |
| 19 | Biała Woda | 771 | N 49.39515° E 020.59550° | Lower montane range | Anthylii-Trifolietum montani |
| 20 | Lubomierz | 768 | N 49.59272° E 020.22380° | Lower montane range | Anthylii-Trifolietum montani |
| 21 | Wołosate | 761 | N 49.06482° E 022.68687° | Lower montane range | Gladiolo-Agrostietum |
| 22 | Żabnica | 736 | N 49.55321° E 019.20479° | Lower montane range | Gladiolo-Agrostietum |
| 23 | Uhryń | 713 | N 49.47237° E 020.86013° | Lower montane range | Gladiolo-Agrostietum |
| 24 | Tarnawa | 677 | N 49.12730° E 022.80056° | Lower montane range | Arrhenatheretum alatioris |
| 25 | Kocoń | 630 | N 49.74408° E 019.40473° | Lower montane range | Gladiolo-Agrostietum |
| 26 | Ciechań | 629 | N 49.45070° E 021.49384° | Lower montane range | Carlino-Dianthetum |
| 27 | Kamesznica | 626 | N 49.60177° E 019.05208° | Lower montane range | Gladiolo-Agrostietum |
| 28 | Ludźmierz | 605 | N 49.47160° E 019.96879° | Lower montane range | Arrhenatheretum alatioris |
| 29 | Łopienka | 592 | N 49.26173° E 022.36151° | Lower montane range | Gladiolo-Agrostietum |
| 30 | Michniowiec | 591 | N 49.30973° E 022.72639° | Lower montane range | Gladiolo-Agrostietum |
| 31 | Sromowce Niżne | 548 | N 49.39893° E 020.39525° | Lower montane range | Anthylii-Trifolietum montani |
| 32 | Żydowskie | 499 | N 49.47831° E 021.46578° | Foothills | Arrhenatheretum alatioris |
| 34 | Gaboń | 488 | N 49.51427° E 020.55303° | Foothills | Arrhenatheretum alatioris |
| 34 | Kamienica | 475 | N 49.58584° E 020.32247° | Foothills | Arrhenatheretum alatioris |
| 35 | Kalnica | 471 | N 49.36662° E 022.19975° | Foothills | Arrhenatheretum alatioris |
| 36 | Nieznajowa | 462 | N 49.49251° E 021.39219° | Foothills | Arrhenatheretum alatioris |
| 37 | Serednica | 454 | N 49.50097° E 022.48839° | Foothills | Arrhenatheretum alatioris |
| 38 | Rytro | 452 | N 49.48491° E 020.69088° | Foothills | Arrhenatheretum alatioris |
| 39 | Rozstajne | 452 | N 49.48896° E 021.41798° | Foothills | Arrhenatheretum medioeuropaeum |
| 40 | Paszowa | 439 | N 49.54168° E 022.41953° | Foothills | Arrhenatheretum medioeuropaeum |
| 41 | Kąty | 407 | N 49.55333° E 021.51637° | Foothills | Arrhenatheretum alatioris |
| 42 | Stefkowa | 406 | N 49.46311° E 022.47645° | Foothills | Arrhenatheretum medioeuropaeum |
| 43 | Stasiówka | 395 | N 50.00342° E 021.46434° | Foothills | Arrhenatheretum alatioris |
| 44 | Krempna | 386 | N 49.51071° E 021.49821° | Foothills | Arrhenatheretum alatioris |
| 45 | Polichty | 362 | N 49.81676° E 020.87115° | Foothills | Arrhenatheretum medioeuropaeum |
| 46 | Zawada | 354 | N 49.98436° E 021.00930° | Foothills | Arrhenatheretum medioeuropaeum |
| 47 | Naszacowice | 342 | N 49.54670° E 020.56232° | Foothills | Arrhenatheretum medioeuropaeum |
| 48 | Gołkowice Dolne | 317 | N 49.54876° E 020.57999° | Foothills | Arrhenatheretum medioeuropaeum |
| 49 | Pleśna | 263 | N 49.92034° E 020.94001° | Foothills | Arrhenatheretum medioeuropaeum |
| No | Species | N | % | Class of Domination |
|---|---|---|---|---|
| 1 | Cercyon (Cercyon) impressus (Sturm, 1807) | 3688 | 38.46% | Superdominant |
| 2 | Sphaeridium lunatum Fabricius, 1792 | 2762 | 28.80% | Dominant |
| 3 | Cercyon (Cercyon) lateralis (Marsham, 1802) | 1126 | 11.74% | |
| 4 | Cercyon (Cercyon) castaneipennis Vorst, 2009 | 780 | 8.13% | |
| 5 | Sphaeridium scarabaeoides (Linnaeus, 1758) | 382 | 3.98% | Subdominant |
| 6 | Cryptopleurum minutum (Fabricius, 1775) | 372 | 3.88% | |
| 7 | Megasternum immaculatum (Stephens, 1829) | 118 | 1.23% | |
| 8 | Sphaeridium bipustulatum Fabricius, 1781 | 88 | 0.92% | Subrecedent |
| 9 | Cercyon (Cercyon) pygmaeus (Illiger, 1801) | 72 | 0.75% | |
| 10 | Cercyon (Cercyon) haemorrhoidalis (Fabricius, 1775) | 59 | 0.62% | |
| 11 | Cercyon (Cercyon) tatricus Endródy-Younga 1967 | 54 | 0.56% | |
| 12 | Sphaeridium marginatum Fabricius 1787 | 45 | 0.47% | |
| 13 | Cercyon (Cercyon) melanocephalus (Linnaeus, 1758) | 19 | 0.20% | |
| 14 | Cercyon (Cercyon) quisquilius (Linnaeus, 1761) | 13 | 0.14% | |
| 15 | Pachysternum capense (Mulsant, 1844) | 6 | 0.06% | |
| 16 | Cercyon (Cercyon) unipunctatus (Linnaeus, 1758) | 4 | 0.04% | |
| 17 | Cercyon (Paracycreon) laminatus Sharp, 1873 | 1 | 0.01% | |
| Total | 9589 | 100.00% |
| No | Site | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | Number of Species at Site |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Kasprowy Wierch | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 2 | Twarda Galeria | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 3 | Kocioł Mułowy | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 4 | Hala Gąsienicowa | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 5 | Upłaziańska Kopa | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 12 | |||||
| 6 | Skupniów Upłaz | ● | ● | ● | 3 | ||||||||||||||
| 7 | Tarnica | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 10 | |||||||
| 8 | Połonina Caryńska | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 9 | Hala Kondratowa | ● | ● | 2 | |||||||||||||||
| 10 | Polana Kopieniec | ● | ● | ● | 3 | ||||||||||||||
| 11 | Przehyba | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 12 | Przełęcz Bukowska | ● | ● | ● | ● | ● | 5 | ||||||||||||
| 13 | Przełęcz Krowiarki | ● | 1 | ||||||||||||||||
| 14 | Kuźnice | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 15 | Zakopane | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 16 | Dolina Kościeliska | ● | ● | ● | 3 | ||||||||||||||
| 17 | Hala Boracza | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 18 | Brzegi Górne | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 10 | |||||||
| 19 | Biała Woda | ● | ● | ● | 3 | ||||||||||||||
| 20 | Lubomierz | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 21 | Wołosate | ● | ● | 2 | |||||||||||||||
| 22 | Żabnica | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 23 | Uhryń | ● | ● | ● | ● | ● | 5 | ||||||||||||
| 24 | Tarnawa | ● | ● | ● | ● | 4 | |||||||||||||
| 25 | Kocoń | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 26 | Ciechań | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 14 | |||
| 27 | Kamesznica | ● | ● | ● | 3 | ||||||||||||||
| 28 | Ludźmierz | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 29 | Łopienka | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 30 | Michniowiec | ● | ● | ● | ● | 4 | |||||||||||||
| 31 | Sromowce Niżne | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 32 | Żydowskie | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 33 | Gaboń | ● | ● | ● | ● | ● | ● | ● | ● | 8 | |||||||||
| 34 | Kamienica | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 35 | Kalnica | ● | ● | ● | ● | ● | ● | ● | ● | 8 | |||||||||
| 36 | Nieznajowa | ● | ● | ● | ● | ● | ● | ● | ● | 8 | |||||||||
| 37 | Serednica | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 38 | Rytro | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 39 | Rozstajne | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 10 | |||||||
| 40 | Paszowa | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 41 | Kąty | ● | ● | 2 | |||||||||||||||
| 42 | Stefkowa | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 43 | Stasiówka | ● | ● | ● | ● | ● | ● | 6 | |||||||||||
| 44 | Krempna | ● | ● | ● | ● | ● | ● | ● | ● | ● | 9 | ||||||||
| 45 | Polichty | ● | ● | ● | ● | ● | 5 | ||||||||||||
| 46 | Zawada | ● | ● | ● | ● | ● | 5 | ||||||||||||
| 47 | Naszacowice | ● | ● | ● | ● | ● | ● | ● | 7 | ||||||||||
| 48 | Gołkowice Dolne | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 13 | ||||
| 49 | Pleśna | ● | ● | ● | ● | ● | ● | ● | 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greń, C.; Górz, A. Coprophagous Hydrophilid Beetles (Coleoptera, Hydrophilidae, Sphaeridiinae) Distribution in the Polish Carpathians. Insects 2020, 11, 355. https://doi.org/10.3390/insects11060355
Greń C, Górz A. Coprophagous Hydrophilid Beetles (Coleoptera, Hydrophilidae, Sphaeridiinae) Distribution in the Polish Carpathians. Insects. 2020; 11(6):355. https://doi.org/10.3390/insects11060355
Chicago/Turabian StyleGreń, Czesław, and Andrzej Górz. 2020. "Coprophagous Hydrophilid Beetles (Coleoptera, Hydrophilidae, Sphaeridiinae) Distribution in the Polish Carpathians" Insects 11, no. 6: 355. https://doi.org/10.3390/insects11060355