Transcriptome Analysis of the Oriental Fruit Fly Bactrocera dorsalis Early Embryos


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Embryo Collection
2.2. RNA Extraction and Library Preparation for Transcriptome Sequencing
2.3. Transcript Sequence Analysis
2.4. Comparison of Differentially Expressed Genes
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. B. Dorsalis Early Embryo Sequencing and Assembly
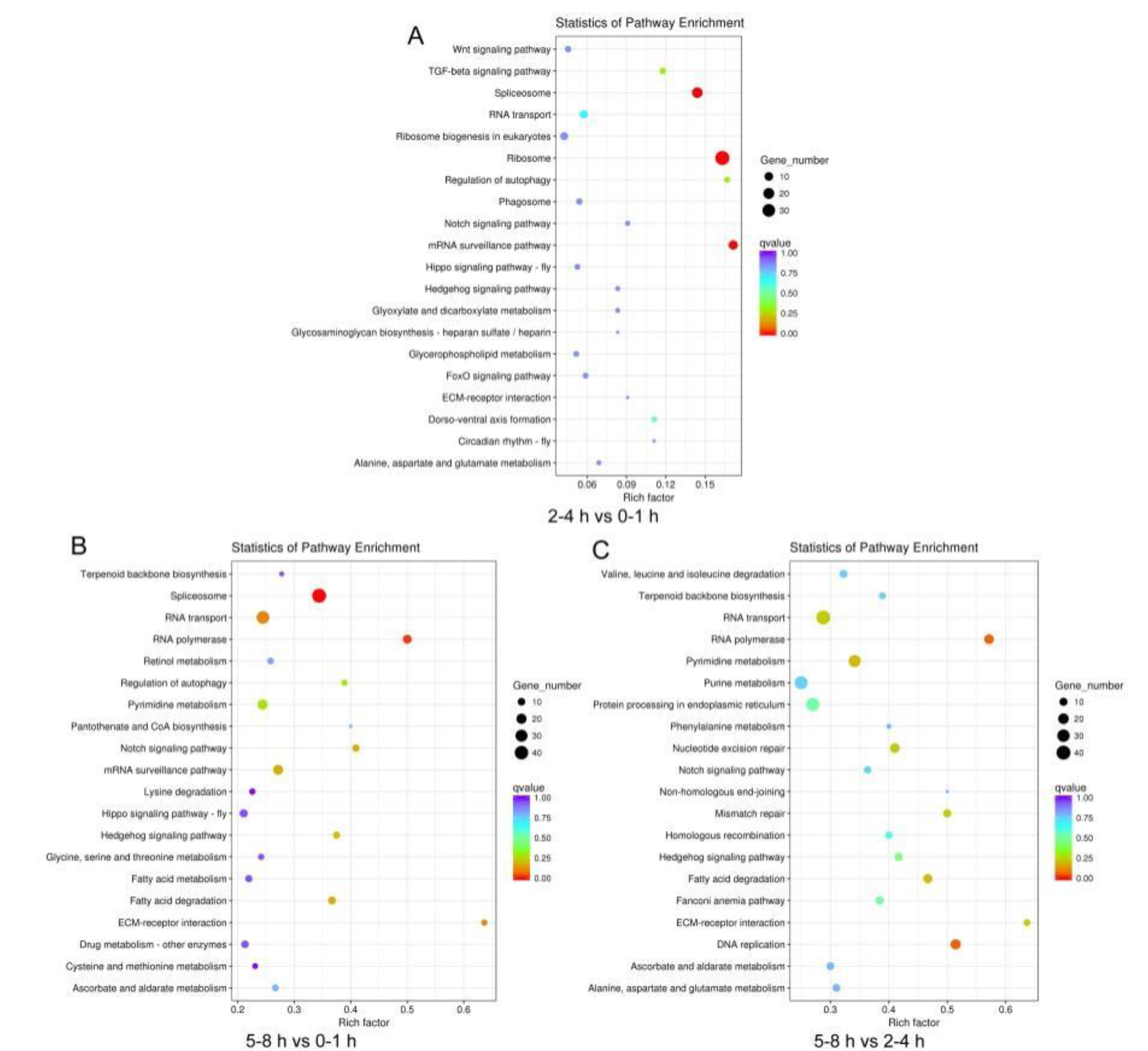
3.2. Comparison of Gene Expression Profiles in Three Sequential Early Embryo Stages
3.3. Unigene Function Annotation and Analysis
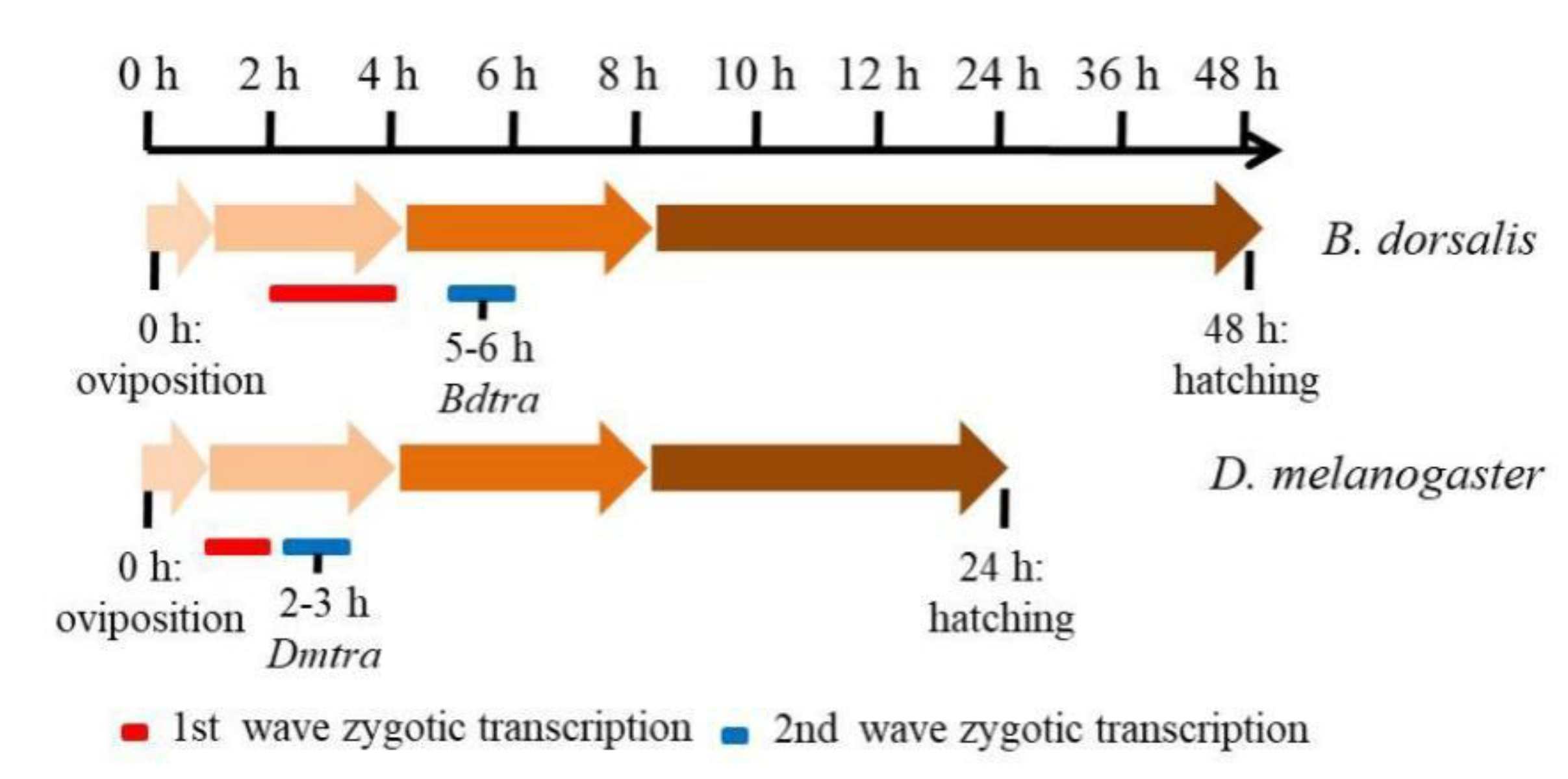
3.4. B. dorsalis Sex Determination and Early Zygotic Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Clarke, A.R.; Armstrong, K.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive Phytophagous Pests Arising through a Recent Tropical Evolutionary Radiation: The Bactrocera Dorsalis Complex of Fruit Flies. Annu. Rev. Èntomol. 2005, 50, 293–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, H. Photographic Guide to Key Control Techniques for Citrus Disease and Insect Pests; Chinese Agricultural Press: Beijing, China, 2012. [Google Scholar]
- Hsu, J.-C.; Feng, H.-T.; Wu, W.-J. Resistance and synergistic effects of insecticides in Bactrocera dorsalis (Diptera: Tephritidae) in Taiwan. J. Econ. Èntomol. 2004, 97, 1682–1688. [Google Scholar] [CrossRef] [PubMed]
- Vontas, J.; Hernández-Crespo, P.; Margaritopoulos, J.T.; Ortego, F.; Feng, H.-T.; Mathiopoulos, K.D.; Hsu, J.-C. Insecticide resistance in Tephritid flies. Pestic. Biochem. Physiol. 2011, 100, 199–205. [Google Scholar] [CrossRef]
- Ant, T.; Koukidou, M.; Rempoulakis, P.; Gong, H.-F.; Economopoulos, A.; Vontas, J.; Alphey, L.S. Control of the olive fruit fly using genetics-enhanced sterile insect technique. BMC Biol. 2012, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.S. Genetic sexing strains in medfly, Ceratitis capitata, sterile insect technique programmes. Genetica 2002, 116, 5–13. [Google Scholar] [CrossRef]
- Dafa’Alla, T.; Fu, G.; Alphey, L.S. Use of a regulatory mechanism of sex determination in pest insect control. J. Genet. 2010, 89, 301–305. [Google Scholar] [CrossRef]
- Koukidou, M.; Alphey, L.S. Practical Applications of Insects’ Sexual Development for Pest Control. Sex. Dev. 2014, 8, 127–136. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Cáceres, C.; Zacharopoulou, A.; Franz, G.; Wimmer, E. Conditional embryonic lethality to improve the sterile insect technique in Ceratitis capitata (Diptera: Tephritidae). BMC Biol. 2009, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Horn, C.; Wimmer, E. A transgene-based, embryo-specific lethality system for insect pest management. Nat. Biotechnol. 2002, 21, 64–70. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Handler, A.M. A transgenic embryonic sexing system for Anastrepha suspensa (Diptera: Tephritidae). Insect Biochem. Mol. Biol. 2012, 42, 790–795. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Handler, A.M. Strategy for enhanced transgenic strain development for embryonic conditional lethality in Anastrepha suspensa. Proc. Natl. Acad. Sci. USA 2012, 109, 9348–9353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concha, C.; Scott, M.J. Sexual Development in Lucilia cuprina (Diptera, Calliphoridae) Is Controlled by the Transformer Gene. Genetics 2009, 182, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hediger, M.; Henggeler, C.; Meier, N.; Perez, R.; Saccone, G.; Bopp, D. Molecular Characterization of the Key Switch F Provides a Basis for Understanding the Rapid Divergence of the Sex-Determining Pathway in the Housefly. Genetics 2009, 184, 155–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos, D.; Koukidou, M.; Savakis, C.; Komitopoulou, K. The transformer gene in Bactrocera oleae: The genetic switch that determines its sex fate. Insect Mol. Biol. 2007, 16, 221–230. [Google Scholar] [CrossRef]
- Pane, A.; Salvemini, M.; Bovi, P.D.; Polito, C.; Saccone, G. The transformer gene in Ceratitis capitata provides a genetic basis for selecting and remembering the sexual fate. Development 2002, 129, 3715–3725. [Google Scholar]
- Peng, W.; Zheng, W.; Handler, A.M.; Zhang, H. The role of the transformer gene in sex determination and reproduction in the tephritid fruit fly, Bactrocera dorsalis (Hendel). Genetica 2015, 143, 717–727. [Google Scholar] [CrossRef]
- Salvemini, M.; Robertson, M.; Aronson, B.; Atkinson, P.; Polito, L.C.; Saccone, G. Ceratitis capitata transformer-2 gene is required to establish and maintain the autoregulation of Cctra, the master gene for female sex determination. Int. J. Dev. Biol. 2009, 53, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Schetelig, M.F.; Milano, A.; Saccone, G.; Handler, A.M. Male only progeny in Anastrepha suspensa by RNAi-induced sex reversion of chromosomal females. Insect Biochem. Mol. Biol. 2012, 42, 51–57. [Google Scholar] [CrossRef]
- Fu, G.; Condon, K.C.; Epton, M.J.; Gong, P.; Jin, L.; Condon, G.C.; Morrison, N.I.; Dafa’Alla, T.H.; Alphey, L.S. Female-specific insect lethality engineered using alternative splicing. Nat. Biotechnol. 2007, 25, 353–357. [Google Scholar] [CrossRef]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.S.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Walker, A.S.; Fu, G.; Harvey-Samuel, T.; Dafa’Alla, T.; Miles, A.; Marubbi, T.; Granville, D.; Humphrey-Jones, N.; O’Connell, S.; et al. Engineered Female-Specific Lethality for Control of Pest Lepidoptera. ACS Synth. Biol. 2013, 2, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Lees, R.; Nimmo, D.; Aw, D.; Jin, L.; Gray, P.; Berendonk, T.U.; White-Cooper, H.; Scaife, S.; Phuc, H.K.; et al. Female-specific flightless phenotype for mosquito control. Proc. Natl. Acad. Sci. USA 2010, 107, 4550–4554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadros, W.; Lipshitz, H.D. The maternal-to-zygotic transition: A play in two acts. Development 2009, 136, 3033–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadros, W.; Goldman, A.L.; Babak, T.; Menzies, F.; Vardy, L.; Orr-Weaver, T.; Hughes, T.R.; Westwood, J.T.; Smibert, C.A.; Lipshitz, H.D. SMAUG Is a Major Regulator of Maternal mRNA Destabilization in Drosophila and Its Translation Is Activated by the PAN GU Kinase. Dev. Cell 2007, 12, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Bushati, N.; Stark, A.; Brennecke, J.; Cohen, S.M. Temporal Reciprocity of miRNAs and Their Targets during the Maternal-to-Zygotic Transition in Drosophila. Curr. Biol. 2008, 18, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Benoit, B.; He, C.H.; Zhang, F.; Votruba, S.M.; Tadros, W.; Westwood, J.T.; Smibert, C.A.; Lipshitz, H.D.; Theurkauf, W.E. An essential role for the RNA-binding protein Smaug during the Drosophila maternal-to-zygotic transition. Development 2009, 136, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.R.T. The TAGteam DNA motif controls the timing of Drosophila pre-blastoderm transcription. Development 2006, 133, 1967–1977. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.-L.; Nien, C.-Y.; Liu, H.-Y.; Metzstein, M.M.; Kirov, N.; Rushlow, C.A. The zinc-finger protein Zelda is a key activator of the early zygotic genome in Drosophila. Nature 2008, 456, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Erickson, J.W.; Cline, T.W. A bZIP protein, sisterless-a, collaborates with bHLH transcription factors early in Drosophila development to determine sex. Genome Res. 1993, 7, 1688–1702. [Google Scholar] [CrossRef] [Green Version]
- Lecuit, T.; Samanta, R.; Wieschaus, E. slam Encodes a Developmental Regulator of Polarized Membrane Growth during Cleavage of the Drosophila Embryo. Dev. Cell 2002, 2, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Stein, J.A.; Broihier, H.T.; Moore, L.A.; Lehmann, R. Slow as Molasses is required for polarized membrane growth and germ cell migration in Drosophila. Development 2002, 129, 3925–3934. [Google Scholar] [PubMed]
- Schejter, E.D.; Wieschaus, E. Bottleneck acts as a regulator of the microfilament network governing cellularization of the Drosophila embryo. Cell 1993, 75, 373–385. [Google Scholar] [CrossRef]
- Rose, L.S.; Wieschaus, E. The Drosophila cellularization gene nullo produces a blastoderm-specific transcript whose levels respond to the nucleocytoplasmic ratio. Genes Dev. 1992, 6, 1255–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhulst, E.C.; Van De Zande, L.; Beukeboom, L.W. Insect sex determination: It all evolves around transformer. Curr. Opin. Genet. Dev. 2010, 20, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, M.F.; Eirin-Lopez, J.M.; Stefani, R.N.; Perondini, A.L.P.; Selivon, D.; Sánchez, L. The gene doublesex of Anastrepha fruit flies (Diptera, Tephritidae) and its evolution in insects. Dev. Genes Evol. 2007, 217, 725–731. [Google Scholar] [CrossRef]
- Ruiz, M.F.; Milano, A.; Salvemini, M.; Eirin-Lopez, J.M.; Perondini, A.L.P.; Selivon, D.; Polito, C.; Saccone, G.; Sánchez, L. The Gene Transformer of Anastrepha Fruit Flies (Diptera, Tephritidae) and Its Evolution in Insects. PLoS ONE 2007, 2, e1239. [Google Scholar] [CrossRef] [Green Version]
- Sarno, F.; Ruiz, M.F.; Eirin-Lopez, J.M.; Perondini, A.; Selivon, D.; Sánchez, L. The gene transformer-2 of Anastrepha fruit flies (Diptera, Tephritidae) and its evolution in insects. BMC Evol. Biol. 2010, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Salz, H.; Erickson, J.W. Sex determination in Drosophila the view from the top. Fly 2010, 4, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Penalva, L.O.F.; Sánchez, L. RNA Binding Protein Sex-Lethal (Sxl) and Control of Drosophila Sex Determination and Dosage Compensation. Microbiol. Mol. Biol. Rev. 2003, 67, 343–359. [Google Scholar] [CrossRef] [Green Version]
- Erickson, J.W.; Quintero, J.J. Indirect effects of ploidy suggest X chromosome dose, not the X:A ratio, signals sex in Drosophila. PLoS Biol. 2007, 5, e332. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.S.; Cline, T.W. Drosophila switch gene Sex-lethal can bypass its switch-gene target transformer to regulate aspects of female behavior. Proc. Natl. Acad. Sci. USA 2013, 110, E4474–E4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashiyama, K.; Hayashi, Y.; Kobayashi, S. Drosophila Sex lethal Gene Initiates Female Development in Germline Progenitors. Science 2011, 333, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Lagos, D.; Ruiz, M.F.; Sánchez, L.; Komitopoulou, K. Isolation and characterization of the Bactrocera oleae genes orthologous to the sex determining Sex-lethal and doublesex genes of Drosophila melanogaster. Gene 2005, 348, 111–121. [Google Scholar] [CrossRef]
- Saccone, G.; Peluso, I.; Artiaco, D.; Giordano, E.; Bopp, D.; Polito, L.C. The Ceratitis capitata homologue of the Drosophila sex-determining gene sex-lethal is structurally conserved, but not sex-specifically regulated. Development 1998, 125, 1495–1500. [Google Scholar] [PubMed]
- Meise, M.; Hilfiker-Kleiner, D.; Dübendorfer, A.; Brunner, C.; Nöthiger, R.; Bopp, D. Sex-lethal, the master sex-determining gene in Drosophila, is not sex-specifically regulated in Musca domestica. Development 1998, 125, 1487–1494. [Google Scholar] [PubMed]
- Hall, A.B.; Basu, S.; Jiang, X.; Qi, Y.; Timoshevskiy, V.A.; Biedler, J.K.; Sharakhova, M.V.; Elahi, R.; Anderson, M.A.E.; Chen, X.-G.; et al. A male-determining factor in the mosquito Aedes aegypti. Science 2015, 348, 1268–1270. [Google Scholar] [CrossRef] [Green Version]
- Krzywinska, E.; Dennison, N.J.; Lycett, G.J.; Krzywinski, J. A maleness gene in the malaria mosquito Anopheles gambiae. Science 2016, 353, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Heinze, S.D.; Wu, Y.; Kohlbrenner, T.; Morilla, I.; Brunner, C.; Wimmer, E.; Van De Zande, L.; Robinson, M.D.; Beukeboom, L.W.; et al. Male sex in houseflies is determined byMdmd, a paralog of the generic splice factor geneCWC22. Science 2017, 356, 642–645. [Google Scholar] [CrossRef]
- Criscione, F.; Qi, Y.; Tu, Z. GUY1 confers complete female lethality and is a strong candidate for a male-determining factor in Anopheles stephensi. eLife 2016, 5, 219. [Google Scholar] [CrossRef]
- Meccariello, A.; Salvemini, M.; Primo, P.; Hall, A.B.; Koskinioti, P.; Dalíková, M.; Gravina, A.; Gucciardino, M.A.; Forlenza, F.; Gregoriou, M.-E.; et al. Maleness-on-the-Y (MoY) orchestrates male sex determination in major agricultural fruit fly pests. Science 2019, 365, 1457–1460. [Google Scholar] [CrossRef]
- Peng, W.; Tariq, K.; Zheng, W.-W.; Yu, S.-N.; Zhang, H.-Y. MicroRNA Let-7 targets the ecdysone signaling pathway E75 gene to control larval–pupal development in Bactrocera dorsalis. Insect Sci. 2017, 26, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000research 2013, 2, 188. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gabrieli, P.; Falaguerra, A.; Siciliano, P.; Gomulski, L.M.; Scolari, F.; Zacharopoulou, A.; Franz, G.; Malacrida, A.; Gasperi, G. Sex and the single embryo: Early deveopment in the Mediterranean fruit fly, Ceratitis capitata. BMC Dev. Biol. 2010, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.-H.; Hou, Q.; Dou, W.; Wei, D.-D.; Yue, Y.; Yang, R.-L.; Yu, S.-F.; De Schutter, K.; Smagghe, G.; Wang, J.-J. RNA-seq analysis of gene expression changes during pupariation in Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). BMC Genom. 2018, 19, 693. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Luo, D.; Wu, F.; Wang, J.-L.; Zhang, H. RNA sequencing to characterize transcriptional changes of sexual maturation and mating in the female oriental fruit fly Bactrocera dorsalis. BMC Genom. 2016, 17, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Peng, T.; He, W.; Zhang, H. High-Throughput Sequencing to Reveal Genes Involved in Reproduction and Development in Bactrocera dorsalis (Diptera: Tephritidae). PLoS ONE 2012, 7, e36463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.-M.; Dou, W.; Niu, J.-Z.; Jiang, H.-B.; Yang, W.-J.; Jia, F.-X.; Hu, F.; Cong, L.; Wang, J.-J. Transcriptome Analysis of the Oriental Fruit Fly (Bactrocera dorsalis). PLoS ONE 2011, 6, e29127. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Bachtrog, D. Reversal of an ancient sex chromosome to an autosome in Drosophila. Nature 2013, 499, 332–335. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Kozhina, E.; Mahadevaraju, S.; Yang, D.; Avila, F.W.; Erickson, J.W. Maternal Groucho and bHLH repressors amplify the dose-sensitive X chromosome signal in Drosophila sex determination. Dev. Biol. 2008, 323, 248–260. [Google Scholar] [CrossRef] [Green Version]
- Harrison, D. Sex determination: Controlling the master. Curr. Biol. 2007, 17, R328–R330. [Google Scholar] [CrossRef] [Green Version]
- Salvemini, M.; Arunkumar, K.P.; Nagaraju, J.; Sanges, R.; Petrella, V.; Tomar, A.; Zhang, H.; Zheng, W.; Saccone, G. De Novo Assembly and Transcriptome Analysis of the Mediterranean Fruit Fly Ceratitis capitata Early Embryos. PLoS ONE 2014, 9, e114191. [Google Scholar] [CrossRef] [Green Version]
- Morrow, J.L.; Riegler, M.; Frommer, M.; Shearman, D. Expression patterns of sex-determination genes in single male and female embryos of two Bactrocera fruit fly species during early development. Insect Mol. Biol. 2014, 23, 754–767. [Google Scholar] [CrossRef]
- Jiménez-Guri, E.; Huerta-Cepas, J.; Cozzuto, L.; Wotton, K.R.; Kang, H.; Himmelbauer, H.; Roma, G.; Gabaldon, T.; Jaeger, J. Comparative transcriptomics of early dipteran development. BMC Genom. 2013, 14, 123. [Google Scholar] [CrossRef] [Green Version]
- Tomancak, P.; Beaton, A.; Weiszmann, R.; Kwan, E.; Shu, S.; Lewis, S.E.; Richards, S.; Ashburner, M.; Hartenstein, V.; Celniker, S.E.; et al. Systematic determination of patterns of gene expression during Drosophila embryogenesis. Genome Biol. 2002, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lemke, S.; Antonopoulos, D.A.; Meyer, F.; Domanus, M.H.; Schmidt-Ott, U. BMP signaling components in embryonic transcriptomes of the hover fly Episyrphus balteatus (Syrphidae). BMC Genom. 2011, 12, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomulski, L.M.; Dimopoulos, G.; Xi, Z.; Soares, M.B.; Bonaldo, M.D.F.; Malacrida, A.; Gasperi, G. Gene discovery in an invasive tephritid model pest species, the Mediterranean fruit fly, Ceratitis capitata. BMC Genom. 2008, 9, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gempe, T.; Beye, M. Function and evolution of sex determination mechanisms, genes and pathways in insects. BioEssays 2010, 33, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bopp, D.; Saccone, G.; Beye, M. Sex Determination in Insects: Variations on a Common Theme. Sex. Dev. 2014, 8, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Wu, Q.; Li, J.-W.; Zhang, G.; Wan, F.-H. RNAi-Mediated Knock-Down of transformer and transformer 2 to Generate Male-Only Progeny in the Oriental Fruit Fly, Bactrocera dorsalis (Hendel). PLoS ONE 2015, 10, e0128892. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-L.; Dai, S.-M.; Lu, K.-H.; Chang, C. Female-specific doublesex dsRNA interrupts yolk protein gene expression and reproductive ability in oriental fruit fly, Bactrocera dorsalis (Hendel). Insect Biochem. Mol. Biol. 2008, 38, 155–165. [Google Scholar] [CrossRef]
- Laohakieat, K.; Aketarawong, N.; Isasawin, S.; Thitamadee, S.; Thanaphum, S. The study of the transformer gene from Bactrocera dorsalis and B. correcta with putative core promoter regions. BMC Genet. 2016, 17, 34. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, L. Sex-determining mechanisms in insects. Int. J. Dev. Biol. 2008, 52, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Salz, H. Sex determination in insects: A binary decision based on alternative splicing. Curr. Opin. Genet. Dev. 2011, 21, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Hoshijima, K.; Inoue, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science 1991, 252, 833–836. [Google Scholar] [CrossRef]
- Hediger, M.; Siegenthaler, C.; Moser, M.; Bopp, D. The transformer2 gene in Musca domestica is required for selecting and maintaining the female pathway of development. Dev. Genes Evol. 2005, 215, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, D.K.; Schubiger, G. Activation of transcription in Drosophila embryos is a gradual process mediated by the nucleocytoplasmic ratio. Genes Dev. 1996, 10, 1131–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetelig, M.F.; Targovska, A.; Meza, J.S.; Bourtzis, K.; Handler, A.M. Tetracycline-suppressible female lethality and sterility in the Mexican fruit fly, Anastrepha ludens. Insect Mol. Biol. 2016, 25, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Linger, R.J.; Scott, M.J. Building early-larval sexing systems for genetic control of the Australian sheep blow fly Lucilia cuprina using two constitutive promoters. Sci. Rep. 2017, 7, 2538. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Williamson, M.E.; Davis, R.J.; Andere, A.A.; Picard, C.J.; Scott, M.J. Improved transgenic sexing strains for genetic control of the Australian sheep blow fly Lucilia cuprina using embryo-specific gene promoters. Mol. Genet. Genom. 2019, 295, 287–298. [Google Scholar] [CrossRef]
- Chen, C.-H.; Huang, H.; Ward, C.M.; Su, J.T.; Schaeffer, L.V.; Guo, M.; Hay, B.A. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 2007, 316, 597–600. [Google Scholar] [CrossRef]
- Kyrou, K.; Hammond, A.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Niu, B.; Ji, D.; Li, M.; Li, K.; James, A.A.; Tan, A.; Huang, Y. Silkworm genetic sexing through W chromosome-linked, targeted gene integration. Proc. Natl. Acad. Sci. USA 2018, 115, 8752–8756. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Reads | 272,036,418 |
|---|---|
| Total number of unigenes | 13,489 |
| Average transcript length (bp) | 2185 |
| Minimum (bp) | 61 |
| Maximum (bp) | 57,167 |
| Number of transcripts >1 Kb | 9809 |
| Number of transcripts >2 Kb | 5227 |
| Number of transcripts >3 Kb | 2873 |
| Number of transcripts >5 Kb | 1066 |
| Number of transcripts >10 Kb | 128 |
| Sample Name | 0–1 h | 2–4 h | 5–8 h |
|---|---|---|---|
| Raw reads | 84,969,396 | 87,332,604 | 99,734,418 |
| Clean reads | 83,787,930 | 85,655,698 | 98,332,254 |
| Clean bases | 12.57 G | 12.85 G | 14.75 G |
| Total mapped | 73,099,585 (87.24%) | 73,131,953 (85.38%) | 84,393,170 (85.82%) |
| Multiple mapped | 228,708 (0.27%) | 223,940 (0.26%) | 336,793 (0.34%) |
| Uniquely mapped | 72,870,877 (86.97%) | 72,908,013 (85.12%) | 84,056,377 (85.48%) |
| Error rate (%) | 0.02 | 0.02 | 0.02 |
| Q 20 (%) | 97.48 | 96.93 | 97.38 |
| Q 30 (%) | 92.72 | 91.65 | 92.58 |
| GC content (%) | 40.94 | 40.59 | 40.71 |
| Gene Name | Gene ID | Gene Length (bp) | ORF Length (aa) | 0–1 h | 2–4 h | 5–8 h |
|---|---|---|---|---|---|---|
| Bddaughterless (Bdda) | 105221792 | 3380 | 710 | 6.19 | 4.73 | 7.30 |
| Bddeadpan (Bddpn) | 105228067 | 2409 | 576 | 0.01 | 0.59 | 44.00 |
| Bddoublesex (Bddsx) | 105226634 | 3858 | 389 | 0.25 | 0.60 | 0.17 |
| Bdfemale-lethal(2)d (Bdfl(2)d) | 105224548 | 3326 | 641 | 31.72 | 49.32 | 51.90 |
| Bdfruitless (Bdfru) | 105224692 | 3346 | 839 | 0 | 0.03 | 29.84 |
| Bdhopscotch (Bdhop) | 105225518 | 3921 | 1175 | 62.75 | 51.02 | 54.04 |
| Bdscute (Bdsc) | 105227369 | 1122 | 296 | 0 | 0.10 | 57.87 |
| Bdsisterless A (BdsisA) | 105231921 | 1075 | 259 | 72.23 | 164.06 | 524.97 |
| BdSex-lethal (BdSxl) | 105232012 | 1938 | 354 | 57.67 | 119.48 | 169.84 |
| Bdtransformer (Bdtra) | 105232904 | 2577 | 422 | 18.31 | 19.48 | 42.76 |
| Bdtransformer-2 (Bdtra-2) | 105222300 | 1676 | 252 | 97.08 | 121.79 | 91.93 |
| Bdvirilizer (Bdvir) | 105229158 | 5842 | 1873 | 33.11 | 24.47 | 47.36 |
| Bdgroucho (Bdgro) | 105227097 | 2981 | 727 | 53.30 | 99.88 | 197.99 |
| BdMes-4 | 105222825 | 4961 | 1556 | 77.50 | 43.23 | 39.14 |
| Bdmdg4 | 105224476 | 1808 | 509 | 27.18 | 12.89 | 33.83 |
| Bdlongitudinals (Bdlo) | 105223842 | 1964 | 128 | 208.09 | 323.25 | 216.68 |
| BdRho1 | 105230670 | 1828 | 193 | 207.05 | 267.43 | 258.73 |
| Bdspaghetti-squash (Bdsqh) | 105227302 | 1793 | 498 | 43.11 | 5.09 | 13.02 |
| Bdextra-macrochaetae (Bdemc) | 105224429 | 1646 | 236 | 0.12 | 35.34 | 187.76 |
| Bdovarian tumor (Bdotu) | 105222775 | 5302 | 1260 | 133.30 | 57.03 | 17.55 |
| Bdrunt | 105226934 | 3210 | 496 | 0 | 69.26 | 45.79 |
| BdZelda (BdZld) | 105232172 | 7823 | 1598 | 63.76 | 147.98 | 120.75 |
| Bdnullo | Novel00216 | 782 | 211 | 0 | 68.06 | 2.15 |
| Bdserendipity a (Bdsrya) | 105222678 | 2520 | 663 | 32.58 | 47.88 | 10.82 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, W.; Yu, S.; Handler, A.M.; Zhang, H. Transcriptome Analysis of the Oriental Fruit Fly Bactrocera dorsalis Early Embryos. Insects 2020, 11, 323. https://doi.org/10.3390/insects11050323
Peng W, Yu S, Handler AM, Zhang H. Transcriptome Analysis of the Oriental Fruit Fly Bactrocera dorsalis Early Embryos. Insects. 2020; 11(5):323. https://doi.org/10.3390/insects11050323
Chicago/Turabian StylePeng, Wei, Shuning Yu, Alfred M. Handler, and Hongyu Zhang. 2020. "Transcriptome Analysis of the Oriental Fruit Fly Bactrocera dorsalis Early Embryos" Insects 11, no. 5: 323. https://doi.org/10.3390/insects11050323