Differential Foraging of Indigenous and Exotic Honeybee (Apis mellifera L.) Races on Nectar-Rich Flow in a Subtropical Ecosystem


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Area
2.2. Tested Honeybees
2.3. Weather Data
2.4. Outgoing Workers and Incoming, Pollen-Loaded Foragers
2.5. Data Processing and Statistical Analysis
3. Results
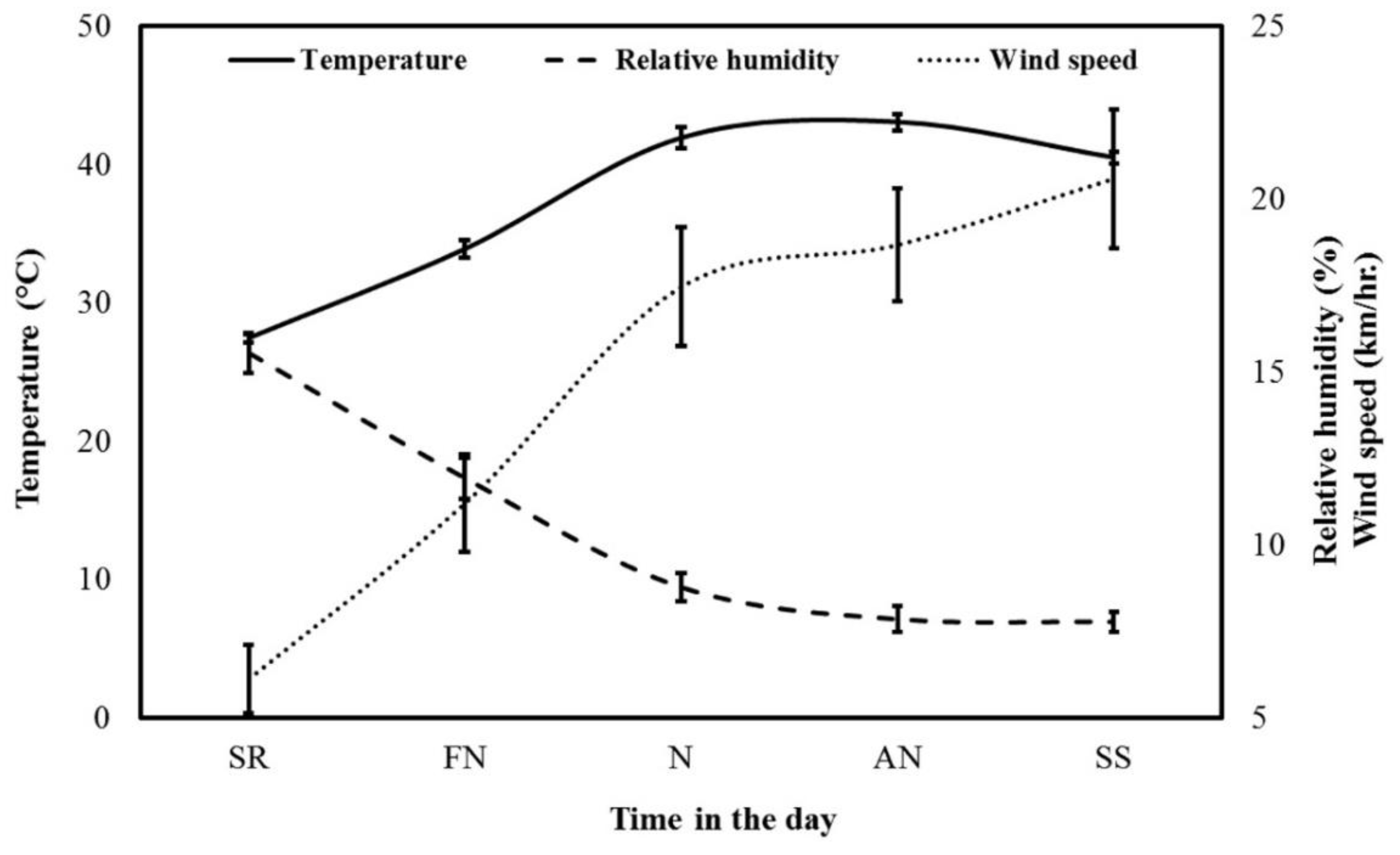
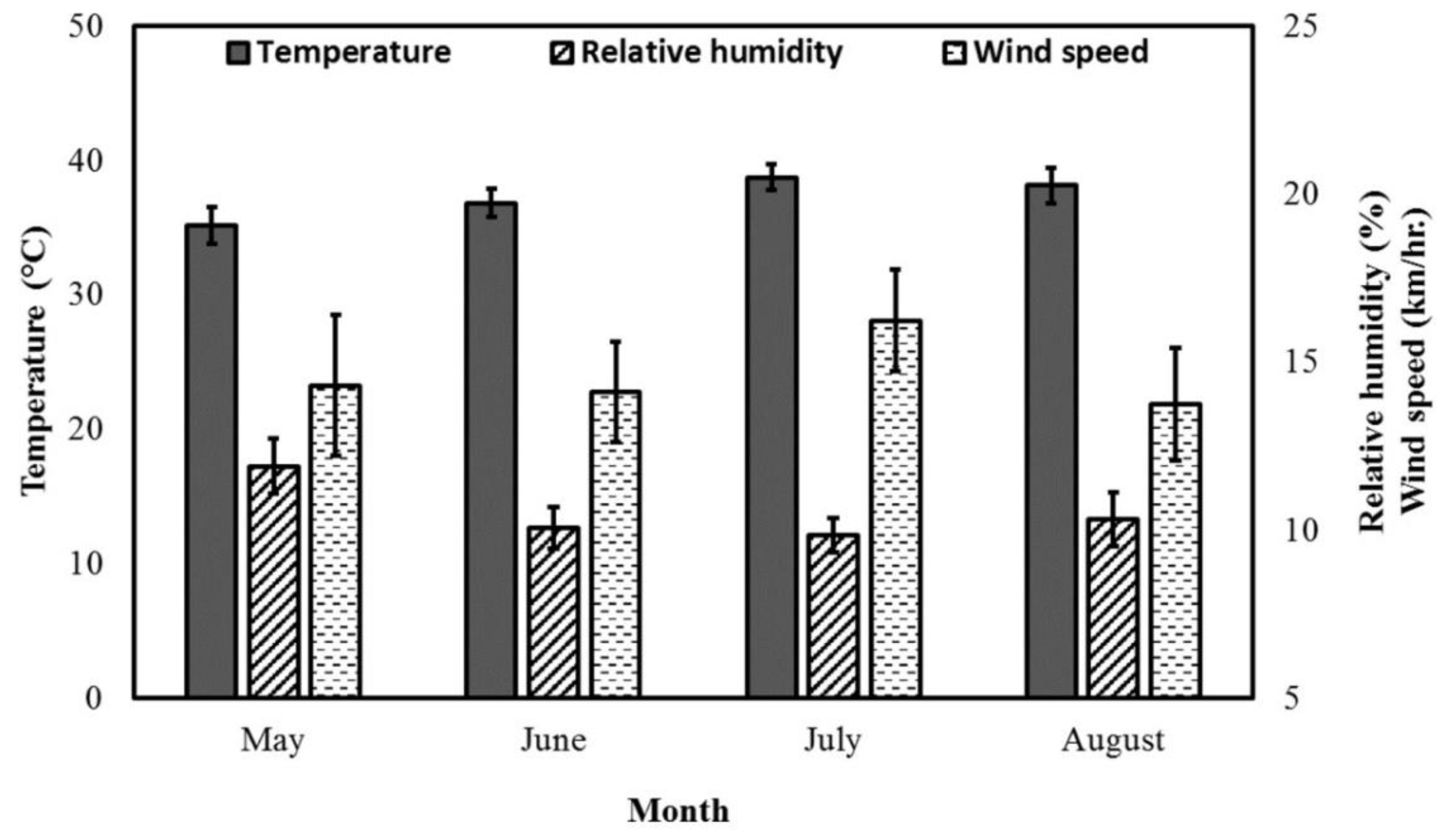
3.1. Weather Data
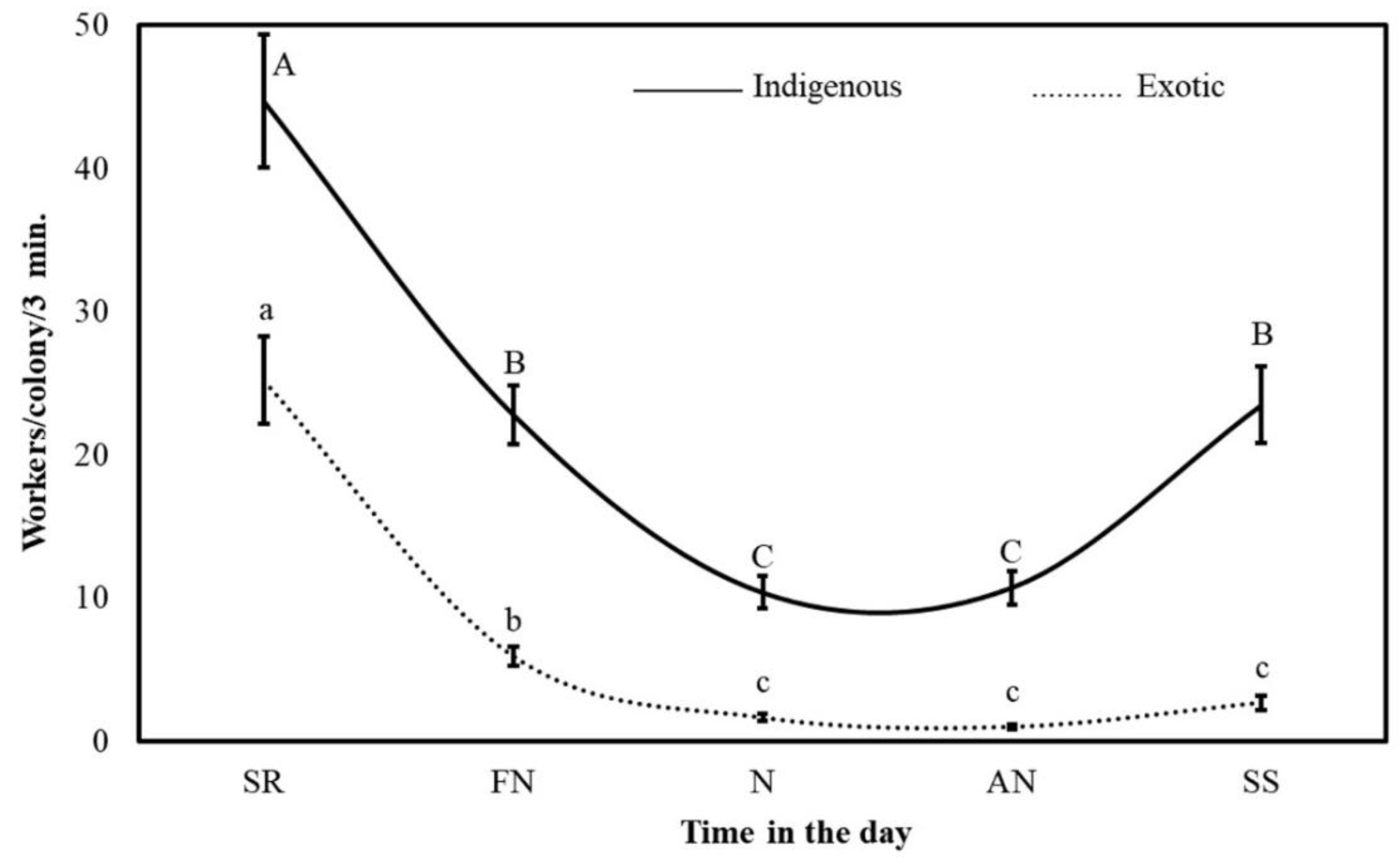
3.2. Foraging Activities
3.3. Pollen-Gathering Activities
3.4. Correlation between Foraging, Pollen Gathering and Weather Factors
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Medina Cuéllar, S.E.; Tirado González, D.N.; Portillo Vázquez, M.; Tirado Estrada, G.; Medina Flores, C.A.; Venegas Venegas, J.A.; Ramos Parra, M. Multifractal detrended fluctuation analysis to characterize honey bee production in semi-arid ecosystems. Interciencia 2018, 43, 498–504. [Google Scholar]
- Dukku, U.H.; Russom, Z.; Domo, A. Diurnal and seasonal flight activity of the honey bee, Apis Mellifera L, and its relationship with temperature, light intensity and relative humidity in the savanna of northern Nigeria. Global J. Sci. Front. Res. Biol. Sci. 2013, 13, 1–7. [Google Scholar]
- Alamu, O.T.; Omoayena, B.O.; Amao, A.O. Flight activity of African honeybee in response to changes in colony size and weather variables during honey flow period in the Northern Guinea Savanna, Nigeria. Int. J. Appl. Res.Technol. 2014, 3, 122–130. [Google Scholar]
- Abou-Shaara, H.F. The foraging behaviour of honey bees, Apis mellifera: A review. Vet. Med.-Czech. 2014, 59, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.B.; Webster, T.C. Apiculture and forestry (bees and trees). Agrofor. Syst. 1995, 29, 313–320. [Google Scholar] [CrossRef]
- Keasar, T.; Shmida, A. An evaluation of Israeli forestry trees and shrubs as potential forage plants for bees. Isr. J. Plant Sci 2009, 57, 49–64. [Google Scholar] [CrossRef]
- Roman, A.; Weryszko, A. Flight activity of honey bee foragers (Apis mellifera L.) under changeable honey flow conditions. Acta. Sci. Pol. Zootechn. 2006, 5, 95–104. [Google Scholar]
- Danka, R.G.; Beaman, L.D. Flight activity of USDA-ARS Russian honey bees (Hymenoptera: Apidae) during pollination of lowbush blueberries in Maine. J. Econ. Entomol. 2007, 100, 267–272. [Google Scholar] [CrossRef]
- Blazyte-Cereskiene, L.; Vaitkeviciene, G.; Venskutonyte, S.; Buda, V. Honey bee foraging in spring oilseed rape crops under high ambient temperature conditions. Zemdirbyste 2010, 97, 61–70. [Google Scholar]
- Tan, K.; Yang, S.; Wang, Z.-W.; Radloff, S.E.; Oldroyd, B.P. Differences in foraging and broodnest temperature in the honey bees Apis cerana and A. mellifera. Apidologie 2012, 43, 618–623. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.; Alqarni, A.S.; Iqbal, J.; Owayss, A.A.; Raweh, H.S.; Smith, B.H. Effect of season and behavioral activity on the hypopharyngeal glands of three honey bee Apis mellifera L. races under stressful climatic conditions of central Saudi Arabia. J. Hymenoptera Res. 2019, 68, 85–101. [Google Scholar] [CrossRef]
- Joshi, N.C.; Joshi, P.C. Foraging behaviour of Apis Spp. on apple flowers in a subtropical environment. N. Y. Sci. J. 2010, 3, 71–76. [Google Scholar]
- Omoloye, A.A.; Akinsol, P.A. Foraging sources and effects of selected plant characters and weather variables on the visitation intensity of honeybee, Apis mellifera adansonii (Hymenoptera: Apidae) in the southwest Nigeria. J. Apic. Sci. 2006, 50, 39–48. [Google Scholar]
- Alqarni, A.S. Tolerance of summer temperature in imported and indigenous honeybee Apis mellifera L. races in central Saudi Arabia. Saudi J. Biol. Sci. 2006, 13, 123–127. [Google Scholar]
- Ali, M.A.M. Comparative study for evaluating two honey bee races, Apis mellifera jemenitica (indigenous race) and Apis mellifera carnica (carniolan race) in brood production, population development and foraging activity under the environmental conditions of the central region of the Kingdom of Saudi Arabia. Ann. Agric. Sci. 2011, 56, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalifa, A.S.; Al-Arify, I.A. Physicochemical characteristics and pollen spectrum of some Saudi honeys. Food. Chem. 1999, 67, 21–25. [Google Scholar] [CrossRef]
- Owayss, A.A.; Elbanna, K.; Iqbal, J.; Abulreesh, H.H.; Organji, S.R.; Raweh, H.S.; Alqarni, A.S. In vitro antimicrobial activities of Saudi honeys originating from Ziziphus spina-christi L. and Acacia gerrardii Benth. trees. Food Sci. Nutr. 2020, 8, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Adgaba, N.; Al-Ghamdi, A.; Tadesse, Y.; Getachew, A.; Awad, A.M.; Ansari, M.J.; Owayss, A.A.; Mohammed, S.E.A.; Alqarni, A.S. Nectar secretion dynamics and honey production potentials of some major honey plants in Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S.; Hannan, M.A.; Owayss, A.A.; Engel, M.S. The indigenous honey bees of Saudi Arabia (Hymenoptera, Apidae, Apis mellifera jemenitica Ruttner): Their natural history and role in beekeeping. Zookeys 2011, 134, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S.; Hassan, A.; Owayss, A. Evaluation of Acacia gerrardii benth. (Fabaceae: Mimosoideae) as a honey plant under extremely hot-dry conditions: Flowering phenology, nectar yield and honey potentiality. J. Anim. Plant Sci. 2015, 25, 1667–1674. [Google Scholar]
- Alqarni, A.S.; Awad, A.M.; Raweh, H.S.A.; Owayss, A.A. Pollination ecology of Acacia gerrardii Benth. (Fabaceae: Mimosoideae) under extremely hot-dry conditions. Saudi J. Biol. Sci. 2017, 24, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.M.; Owayss, A.A.; Alqarni, A.S. Performance of two honey bee subspecies during harsh weather and Acacia gerrardii nectar-rich flow. Sci. Agric. 2017, 74, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Abou-Shaara, H.F.; Al-Ghamdi, A.A.; Mohamed, A.A. Honey bee colonies performance enhance by newly modified beehives. J. Apic. Sci. 2013, 57, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S. Honeybee foraging, nectar secretion, and honey potential of wild jujube trees, Ziziphus nummularia. Neotrop. Entomol. 2015, 44, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.M.A.; Giovanetti, M.; Raweh, H.S.A.; Owayss, A.A.; Ansari, M.J.; Nuru, A.; Radloff, S.E.; Alqarni, A.S. Nectar secretion dynamics of Ziziphus nummularia: A melliferous species of dry land ecosystems. Saudi J. Biol. Sci. 2017, 24, 1470–1474. [Google Scholar] [CrossRef]
- Iqbal, J.; Ali, H.; Owayss, A.A.; Raweh, H.S.A.; Engel, M.S.; Alqarni, A.S.; Smith, B.H. Olfactory associative behavioral differences in three honey bee Apis mellifera L. races under the arid zone ecosystem of central Saudi Arabia. Saudi J. Biol. Sci. 2019, 26, 563–568. [Google Scholar] [CrossRef]
- Alqarni, A.S.; Ali, H.; Iqbal, J.; Owayss, A.A.; Smith, B.H. Expression of heat shock proteins in adult honey bee (Apis mellifera L.) workers under hot-arid subtropical ecosystems. Saudi J. Biol. Sci. 2019, 26, 1372–1376. [Google Scholar] [CrossRef]
- Iqbal, J.; Alqarni, A.S.; Raweh, H.S.A. Effect of sub-lethal doses of imidacloprid on learning and memory formation of indigenous Arabian bee (Apis mellifera jemenitica Ruttner) adult foragers. Neotrop. Entomol. 2019, 48, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.M.; Owayss, A.A.; Iqbal, J.; Raweh, H.S.A.; Alqarni, A.S. GIS approach for determining the optimum spatiotemporal plan for beekeeping and honey production in hot-arid subtropical ecosystems. J. Econ. Entomol. 2019. [Google Scholar] [CrossRef]
- Alfarhan, A.H. A floristic account on Raudhat Khuraim central province Saudi Arabia. Saudi J. Biol. Sci. 2001, 8, 80–103. [Google Scholar]
- Beekman, M.; Sumpter, D.J.T.; Seraphides, N.; Ratnieks, F.L.W. Comparing foraging behaviour of small and large honey-bee colonies by decoding waggle dances made by foragers. Funct. Ecol 2004, 18, 829–835. [Google Scholar] [CrossRef]
- Pernal, S.F.; Currie, R.W. The influence of pollen quality on foraging behavior in honeybees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2001, 51, 53–68. [Google Scholar] [CrossRef]
- Malerbo-Souza, D.T. The corn pollen as a food source for honeybees. Acta. Sci-Agron. 2011, 33, 701–704. [Google Scholar] [CrossRef]
- Pearce, F.C.R.; Couvillon, M.J.; Ratnieks, F.L.W. Hive relocation does not adversely affect honey bee (Hymenoptera: Apidae) foraging. Psyche 2013, 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.S.; Balhareth, H.M.; Owayss, A.A. Performance evaluation of indigenous and exotic honey bee (Apis mellifera L.) races in Assir region, southwestern Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Abou-Shaara, H.F.; Al-Ghamdi, A.A.; Mohamed, A.A. Tolerance of two honey bee races to various temperature and relative humidity gradients. Environ. Exp. Biol. 2012, 10, 133–138. [Google Scholar]
- Hunt, G.J.; Page, R.E.; Fondrk, M.K.; Dullum, C.J. Major quantitative trait loci affecting honey bee foraging behavior. Genetics 1995, 141, 1537–1545. [Google Scholar]
- Pankiw, T.; Tarpy, D.R.; Page, R.E. Genotype and rearing environment affect honeybee perception and foraging behaviour. Anim. Behav. 2002, 64, 663–672. [Google Scholar] [CrossRef]
- Page, R.E.; Rueppell, O.; Amdam, G.V. Genetics of reproduction and regulation of honeybee (Apis mellifera L.) social behavior. Annu. Rev. Genet. 2012, 46, 97–119. [Google Scholar] [CrossRef] [Green Version]
- Page, R.E.; Fondrk, M.K. The effects of colony level selection on the social-organization of honey-bee (Apis mellifera L) colonies: Colony level components of pollen hoarding. Behav. Ecol. Sociobiol. 1995, 36, 135–144. [Google Scholar] [CrossRef]
- Pankiw, T. Directional change in a suite of foraging behaviors in tropical and temperate evolved honey bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2003, 54, 458–464. [Google Scholar] [CrossRef]
- Page, R.E.; Fondrk, M.K.; Rueppell, O. Complex pleiotropy characterizes the pollen hoarding syndrome in honey bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2012, 66, 1459–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueppell, O.; Hunggims, E.; Tingek, S. Association between larger ovaries and pollen foraging in queenless Apis cerana workers supports the reproductive ground-plan hypothesis of social evolution. J. Insect Behav. 2008, 21, 317–321. [Google Scholar] [CrossRef]
- Silva, D.P.; Moisan-De Serres, J.; Souza, D.R.; Hilgert-Moreira, S.B.; Fernandes, M.Z.; Kevan, P.G.; Freitas, B.M. Efficiency in pollen foraging by honey bees: Time, motion and pollen depletion on flowers of Sisyrinchium palmifolium Linnaeus (Asparagales: Iridaceae). J. Pollinat. Ecol. 2013, 11, 27–32. [Google Scholar] [CrossRef]
- Adgaba, N.; Awad, A.M.; Al-Ghamdi, A.A.; Alqarni, A.S.; Radloff, S.E. Nectar of Ziziphus spina-christi (L.) willd (Rhamnaceae): Dynamics of secretion and potential for honey production. J. Apic. Sci. 2012, 56, 49–59. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies | Activity | Temp. | RH | Wind Speed |
|---|---|---|---|---|
| Indigenous | Foraging | −0.357 ** | 0.297 ** | −0.335 ** |
| Pollen gathering | −0.301 ** | 0.289 ** | −0.207 ** | |
| Exotic | Foraging | −0.426 ** | 0.293 ** | −0.319 ** |
| Pollen gathering | −0.449 ** | 0.347 ** | −0.240 ** | |
| Mean | Foraging | −0.424 ** | 0.325 ** | −0.362 ** |
| Pollen gathering | −0.390 ** | 0.339 ** | −0.240 ** |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqarni, A.S. Differential Foraging of Indigenous and Exotic Honeybee (Apis mellifera L.) Races on Nectar-Rich Flow in a Subtropical Ecosystem. Insects 2020, 11, 254. https://doi.org/10.3390/insects11040254
Alqarni AS. Differential Foraging of Indigenous and Exotic Honeybee (Apis mellifera L.) Races on Nectar-Rich Flow in a Subtropical Ecosystem. Insects. 2020; 11(4):254. https://doi.org/10.3390/insects11040254
Chicago/Turabian StyleAlqarni, Abdulaziz S. 2020. "Differential Foraging of Indigenous and Exotic Honeybee (Apis mellifera L.) Races on Nectar-Rich Flow in a Subtropical Ecosystem" Insects 11, no. 4: 254. https://doi.org/10.3390/insects11040254