Concanavalin A Toxicity Towards Potato Psyllid and Apoptosis Induction in Midgut Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect
2.2. Feeding Bioassays
2.3. Nuclear Architecture and TUNEL Assay of Gut Epithelial Cells
2.4. Gene Expression of Caspases
2.5. Statistical Analyses
3. Results
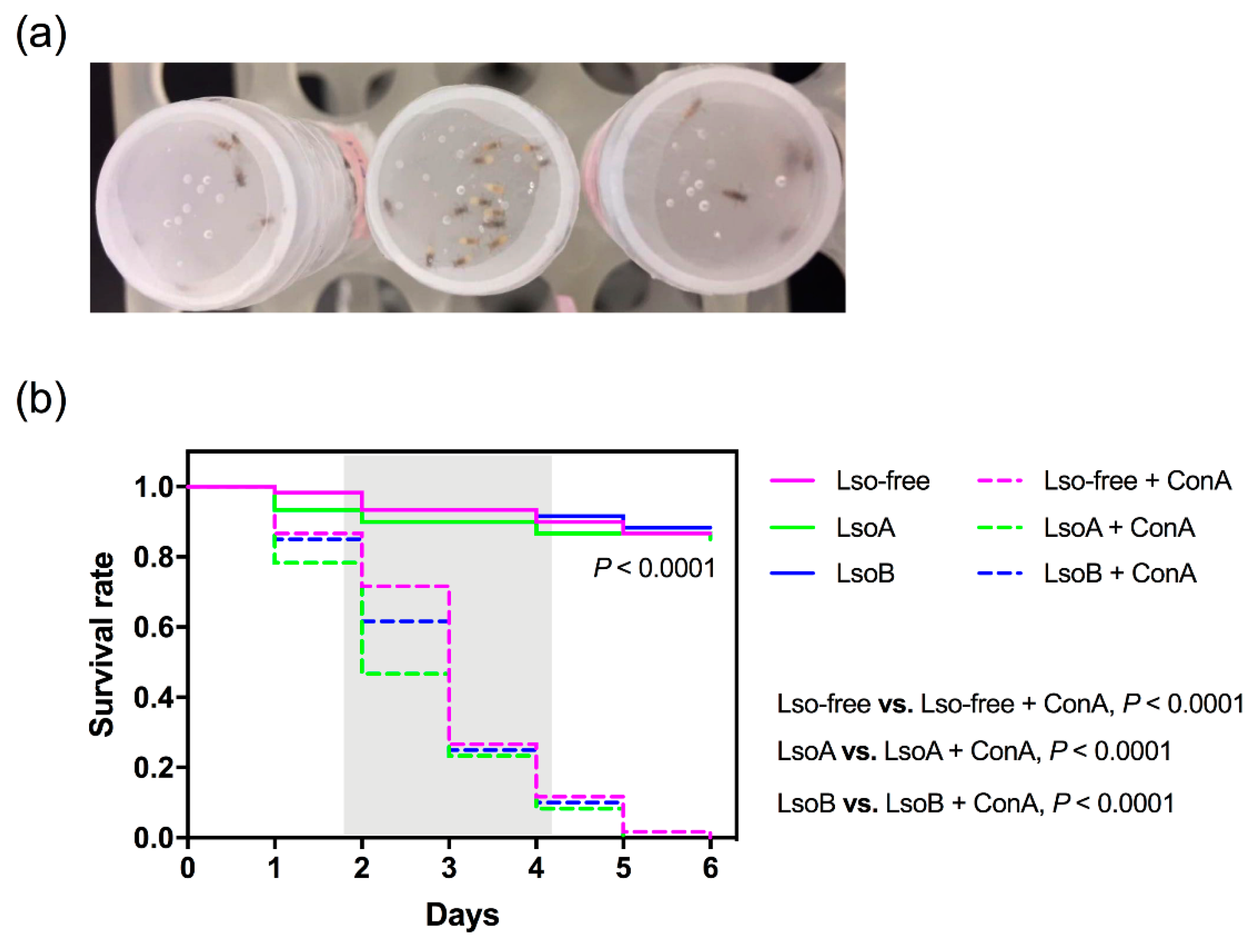
3.1. Mortality of Potato Psyllids Following ConA Treatment
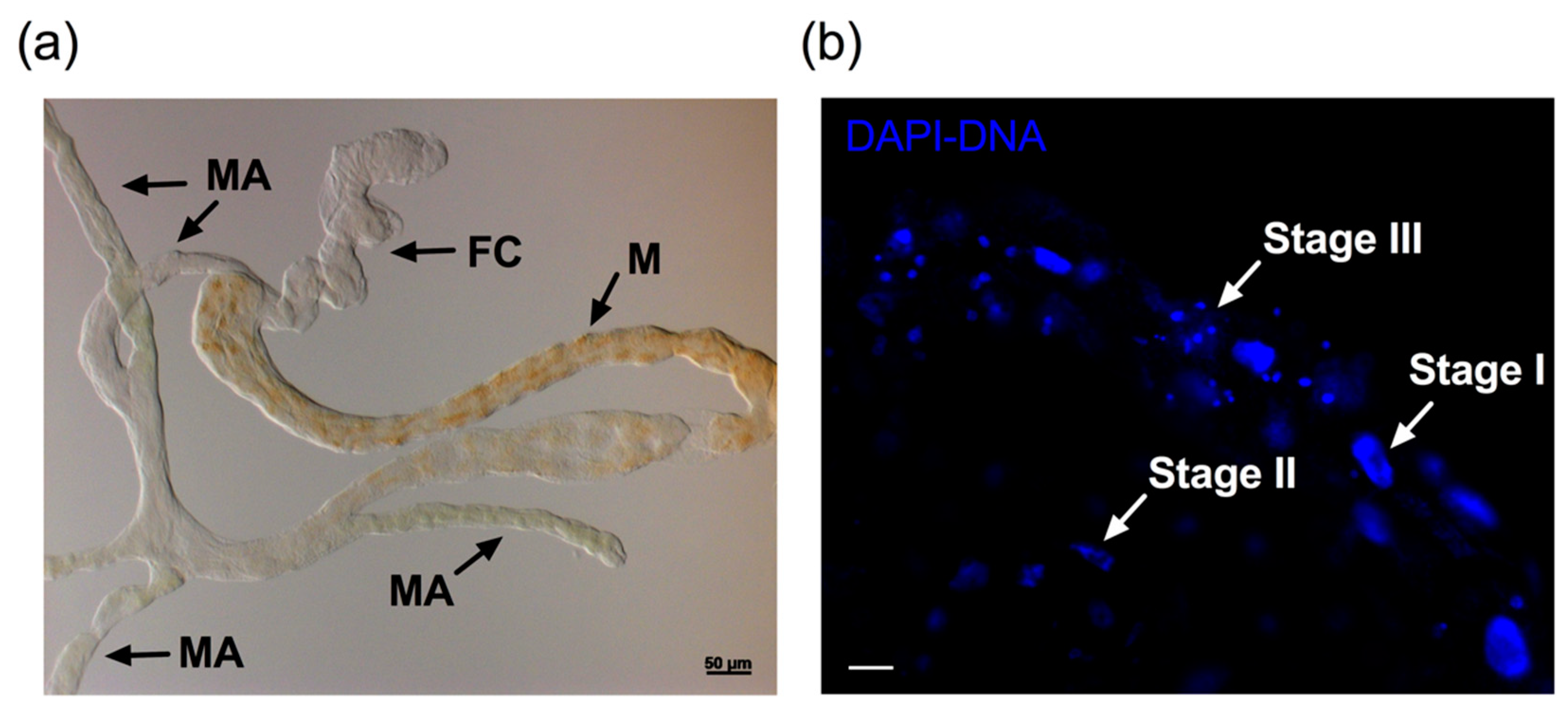
3.2. Nuclear Architecture and TUNEL Assay
3.3. Expression of Caspase Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peumans, W.J.; van Damme, E. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347. [Google Scholar] [CrossRef] [PubMed]
- Kesherwani, V.; Sodhi, A. Differential activation of macrophages in vitro by lectin Concanavalin A, Phytohemagglutinin and wheat germ agglutinin: Production and regulation of nitric oxide. Nitric Oxide 2007, 16, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Vandenborre, G.; Smagghe, G.; Van Damme, E.J. Plant lectins as defense proteins against phytophagous insects. Phytochemistry 2011, 72, 1538–1550. [Google Scholar] [CrossRef] [PubMed]
- Lannoo, N.; Van Damme, E.J. Lectin domains at the frontiers of plant defense. Front. Plant Sci. 2014, 5, 397. [Google Scholar] [CrossRef] [Green Version]
- Jouanin, L.; Bonadé-Bottino, M.; Girard, C.; Morrot, G.; Giband, M. Transgenic plants for insect resistance. Plant. Sci. 1998, 131, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fitches, E.; Wiles, D.; Douglas, A.E.; Hinchliffe, G.; Audsley, N.; Gatehouse, J.A. The insecticidal activity of recombinant garlic lectins towards aphids. Insect Biochem. Mol. Biol. 2008, 38, 905–915. [Google Scholar] [CrossRef]
- Kale, S.; Deshmukh, A. Unexplored and potential applications of pectins in crop protection: A review. J. Cell Tissue Res. Vol. 2017, 17, 6141–6145. [Google Scholar]
- Jang, S.; Yayeh, T.; Leem, Y.-H.; Park, E.-M.; Ito, Y.; Oh, S. Concanavalin a induces cortical neuron apoptosis by causing ROS accumulation and tyrosine kinase activation. Neurochem. Res. 2017, 42, 3504–3514. [Google Scholar] [CrossRef]
- Shukla, S.; Arora, R. Biological activity of soybean trypsin inhibitor and plant lectins against cotton bollworm/legume pod borer, Helicoverpa armigera. Plant Biotechnol. 2005, 22, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Sauvion, N.; Nardon, C.; Febvay, G.; Gatehouse, A.M.; Rahbé, Y. Binding of the insecticidal lectin Concanavalin A in pea aphid, Acyrthosiphon pisum (Harris) and induced effects on the structure of midgut epithelial cells. J. Insect Physiol. 2004, 50, 1137–1150. [Google Scholar] [CrossRef]
- Sprawka, I.; Goławska, S.; Parzych, T.; Sytykiewicz, H.; Czerniewicz, P. Apoptosis induction by concanavalin A in gut cells of grain aphid. Arthropod Plant Interact. 2015, 9, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Sprawka, I.; Goławska, S.; Parzych, T.; Goławski, A.; Czerniewicz, P.; Sytykiewicz, H. Mechanism of entomotoxicity of the concanavalin A in Rhopalosiphum padi (Hemiptera: Aphididae). J. Insect Sci. 2014, 14, 232. [Google Scholar] [CrossRef] [Green Version]
- Zhu-Salzman, K.; Salzman, R.A. Functional mechanics of the plant defensive Griffonia simplicifolia lectin II: Resistance to proteolysis is independent of glycoconjugate binding in the insect gut. J. Econ. Entomol. 2001, 94, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Lagarda-Diaz, I.; Guzman-Partida, A.M.; Urbano-Hernandez, G.; Ortega-Nieblas, M.M.; Robles-Burgueño, M.R.; Winzerling, J.; Vazquez-Moreno, L. Insecticidal action of PF2 lectin from Olneya tesota (Palo Fierro) against Zabrotes subfasciatus larvae and midgut glycoconjugate binding. J. Agric. Food Chem. 2008, 57, 689–694. [Google Scholar] [CrossRef]
- Roy, A.; Das, S. Molecular mechanism underlying the entomotoxic effect of Colocasia esculenta tuber agglutinin against Dysdercus cingulatus. Insects 2015, 6, 827–846. [Google Scholar]
- Walski, T.; Van Damme, E.J.; Smagghe, G. Penetration through the peritrophic matrix is a key to lectin toxicity against Tribolium castaneum. J. Insect Physiol. 2014, 70, 94–101. [Google Scholar] [CrossRef]
- Foissac, X.; Loc, N.T.; Christou, P.; Gatehouse, A.M.; Gatehouse, J.A. Resistance to green leafhopper (Nephotettix virescens) and brown planthopper (Nilaparvata lugens) in transgenic rice expressing snowdrop lectin (Galanthus nivalis agglutinin; GNA). J. Insect Physiol. 2000, 46, 573–583. [Google Scholar] [CrossRef]
- Michiels, K.; Van Damme, E.J.; Smagghe, G. Plant-insect interactions: What can we learn from plant lectins? Arch. Insect Biochem. 2010, 73, 193–212. [Google Scholar] [CrossRef]
- Liu, B.; Bian, H.-J.; Bao, J.-K. Plant lectins: Potential antineoplastic drugs from bench to clinic. Cancer Lett. 2010, 287, 1–12. [Google Scholar] [CrossRef]
- Shi, Z.; Li, W.-W.; Tang, Y.; Cheng, L.-J. A novel molecular model of plant lectin-induced programmed cell death in cancer. Biol. Pharm. Bull. 2017, 40, 1625–1629. [Google Scholar] [CrossRef] [Green Version]
- Kochubei, T.; Maksymchuk, O.; Piven, O.; Lukash, L. Isolectins of phytohemagglutinin are able to induce apoptosis in HEp-2 carcinoma cells in vitro. Exp. Oncol. 2015, 37, 116–119. [Google Scholar] [CrossRef]
- Chan, Y.S.; Yu, H.; Xia, L.; Ng, T.B. Lectin from green speckled lentil seeds (Lens culinaris) triggered apoptosis in nasopharyngeal carcinoma cell lines. Chin. Med. 2015, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Katsuno, Y.; Miyoshi, N.; Hayakawa, S.; Mita, T.; Muto, H.; Isemura, S.; Aoyagi, Y.; Isemura, M. Apoptosis induction by lectin isolated from the mushroom Boletopsis leucomelas in U937 cells. Biosci. Biotechnol. Biochem. 2002, 66, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Tamborindeguy, C.; Huot, O.B.; Ibanez, F.; Levy, J. The influence of bacteria on multitrophic interactions among plants, psyllids, and pathogen. Insect Sci. 2017, 24, 961–974. [Google Scholar] [CrossRef]
- Guenthner, J.; Goolsby, J.; Greenway, G. Use and cost of insecticides to control potato psyllids and zebra chip on potatoes. Southwest. Entomol. 2012, 37, 263–271. [Google Scholar] [CrossRef]
- Levy, J.; Scheuring, D.C.; Koym, J.W.; Henne, D.C.; Tamborindeguy, C.; Pierson, E.; Miller, J.C., Jr. Investigations on putative zebra chip tolerant potato selections. Am. J. Potato Res. 2015, 92, 417–425. [Google Scholar] [CrossRef]
- Gao, L.-Y.; Kwaik, Y.A. Hijacking of apoptotic pathwaysby bacterial pathogens. Microbes Infect. 2000, 2, 1705–1719. [Google Scholar] [CrossRef]
- Yao, J.; Saenkham, P.; Levy, J.; Ibanez, F.; Noroy, C.; Mendoza, A.; Huot, O.; Meyer, D.F.; Tamborindeguy, C. Interactions ‘Candidatus Liberibacter solanacearum’—Bactericera cockerelli: Haplotype effect on vector fitness and gene expression analyses. Front. Cell. Infect. Microbiol. 2016, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.T.; Ibanez, F.; Tamborindeguy, C. Quenching autofluorescence in the alimentary canal tissues of Bactericera cockerelli (Hemiptera: Triozidae) for immunofluorescence labeling. Insect Sci. 2019. [Google Scholar] [CrossRef]
- Wang, X.R.; Wang, C.; Wang, X.W.; Qian, L.X.; Chi, Y.; Liu, S.S.; Liu, Y.Q.; Wang, X.W. The functions of caspase in whitefly Bemisia tabaci apoptosis in response to ultraviolet irradiation. Insect Mol. Biol. 2018, 27, 739–751. [Google Scholar] [CrossRef]
- Nachappa, P.; Levy, J.; Tamborindeguy, C. Transcriptome analyses of Bactericera cockerelli adults in response to “Candidatus Liberibacter solanacearum” infection. Mol. Genet. Genom. 2012, 287, 803–817. [Google Scholar] [CrossRef]
- Tang, X.T.; Tamborindeguy, C. No evidence of apoptotic response of the potato psyllid, Bactericera cockerelli, to “Candidatus Liberibacter solanacearum” at the gut interface. Infect. Immun. 2019. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C T method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, F.; Tamborindeguy, C. Selection of reference genes for expression analysis in the potato psyllid, Bactericera cockerelli. Insect Mol. Biol. 2016, 25, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Kihlmark, M.; Imreh, G.; Hallberg, E. Sequential degradation of proteins from the nuclear envelope during apoptosis. J. Cell Sci. 2001, 114, 3643–3653. [Google Scholar]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Oliveira, C.F.; Oliveira, C.T. Insecticidal activity of plant lectins and potential application in crop protection. Molecules 2015, 20, 2014–2033. [Google Scholar] [CrossRef] [Green Version]
- Napoleão, T.H.; Albuquerque, L.P.; Santos, N.D.; Nova, I.C.; Lima, T.A.; Paiva, P.M.; Pontual, E.V. Insect midgut structures and molecules as targets of plant-derived protease inhibitors and lectins. Pest Manag. Sci. 2019, 75, 1212–1222. [Google Scholar] [CrossRef]
- Down, R.E.; Fitches, E.C.; Wiles, D.P.; Corti, P.; Bell, H.A.; Gatehouse, J.A.; Edwards, J.P. Insecticidal spider venom toxin fused to snowdrop lectin is toxic to the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae) and the rice brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Pest Manag. Sci. 2006, 62, 77–85. [Google Scholar] [CrossRef]
- Chougule, N.P.; Bonning, B.C. Toxins for transgenic resistance to hemipteran pests. Toxins 2012, 4, 405–429. [Google Scholar] [CrossRef]
- George, B.S.; Silambarasan, S.; Senthil, K.; Jacob, J.P.; Ghosh Dasgupta, M. Characterization of an insecticidal protein from Withania somnifera against lepidopteran and hemipteran pest. Mol. Biotechnol. 2018, 60, 290–301. [Google Scholar] [CrossRef]
- Sauvion, N.; Charles, H.; Febvay, G.; Rahbé, Y. Effects of jackbean lectin (ConA) on the feeding behaviour and kinetics of intoxication of the pea aphid, Acyrthosiphon pisum. Entomol. Exp. Appl. 2004, 110, 31–44. [Google Scholar] [CrossRef]
- Albuquerque Tomilhero Frias, A.; Ibanez, F.; Mendoza, A.; de Carvalho Nunes, W.M.; Tamborindeguy, C. Effects of “Candidatus Liberibacter solanacearum”(haplotype B) on Bactericera cockerelli fitness and vitellogenesis. Insect Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M.; Fattah-Hosseini, S.; Levy, A.; Cilia, M. Morphological abnormalities and cell death in the Asian citrus psyllid (Diaphorina citri) midgut associated with Candidatus Liberibacter asiaticus. Sci. Rep. UK 2016, 6, 33418. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef]
- Trębicki, P.; Harding, R.M.; Powell, K.S. Anti-metabolic effects of Galanthus nivalis agglutinin and wheat germ agglutinin on nymphal stages of the common brown leafhopper using a novel artificial diet system. Entomol. Exp. Appl. 2009, 131, 99–105. [Google Scholar] [CrossRef]
- Liener, I. The Lectins: Properties, Functions, and Applications in Biology and Medicine; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Pusztai, A. Plant Lectins-Chemistry and Pharmacology of Natural Products; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Majumder, P.; Banerjee, S.; Das, S. Identification of receptors responsible for binding of the mannose specific lectin to the gut epithelial membrane of the target insects. Glycoconj. J. 2003, 20, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Pace, K.E.; Hahn, H.P.; Baum, L.G. Preparation of recombinant human galectin-1 and use in T-cell death assays. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2003; Volume 363, pp. 499–518. [Google Scholar]
- Hamshou, M.; Smagghe, G.; Shahidi-Noghabi, S.; De Geyter, E.; Lannoo, N.; Van Damme, E.J. Insecticidal properties of Sclerotinia sclerotiorum agglutinin and its interaction with insect tissues and cells. Insect Biochem. Mol. Biol. 2010, 40, 883–890. [Google Scholar] [CrossRef]
- Green, D.R. Means to An End: Apoptosis and Other Cell Death Mechanisms; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2011. [Google Scholar]
- Lorenzo, H.K.; Susin, S.A.; Penninger, J.; Kroemer, G. Apoptosis inducing factor (AIF): A phylogenetically old, caspase-independent effector of cell death. Cell Death Differ. 1999, 6, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770. [Google Scholar] [CrossRef]
- Clarke, T.E.; Clem, R.J. Insect defenses against virus infection: The role of apoptosis. Int. Rev. Immunol. 2003, 22, 401–424. [Google Scholar] [CrossRef] [PubMed]
- Clem, R. The role of apoptosis in defense against baculovirus infection in insects. In Role of Apoptosis in Infection; Springer: Berlin/Heidelberg, Germany, 2005; pp. 113–129. [Google Scholar]
- Huang, H.-J.; Bao, Y.-Y.; Lao, S.-H.; Huang, X.-H.; Ye, Y.-Z.; Wu, J.-X.; Xu, H.-J.; Zhou, X.-P.; Zhang, C.-X. Rice ragged stunt virus-induced apoptosis affects virus transmission from its insect vector, the brown planthopper to the rice plant. Sci. Rep. 2015, 5, 11413. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell 2002, 9, 459–470. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef] [PubMed]
- Habibi, J.; Backus, E.A.; Huesing, J.E. Effects of phytohemagglutinin (PHA) on the structure of midgut epithelial cells and localization of its binding sites in western tarnished plant bug, Lygus hesperus Knight. J. Insect Physiol. 2000, 46, 611–619. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, X.-T.; Ibanez, F.; Tamborindeguy, C. Concanavalin A Toxicity Towards Potato Psyllid and Apoptosis Induction in Midgut Cells. Insects 2020, 11, 243. https://doi.org/10.3390/insects11040243
Tang X-T, Ibanez F, Tamborindeguy C. Concanavalin A Toxicity Towards Potato Psyllid and Apoptosis Induction in Midgut Cells. Insects. 2020; 11(4):243. https://doi.org/10.3390/insects11040243
Chicago/Turabian StyleTang, Xiao-Tian, Freddy Ibanez, and Cecilia Tamborindeguy. 2020. "Concanavalin A Toxicity Towards Potato Psyllid and Apoptosis Induction in Midgut Cells" Insects 11, no. 4: 243. https://doi.org/10.3390/insects11040243