Landscape Effects on the Abundance of Apolygus lucorum in Cotton Fields


Abstract
:1. Introduction
2. Materials and Methods
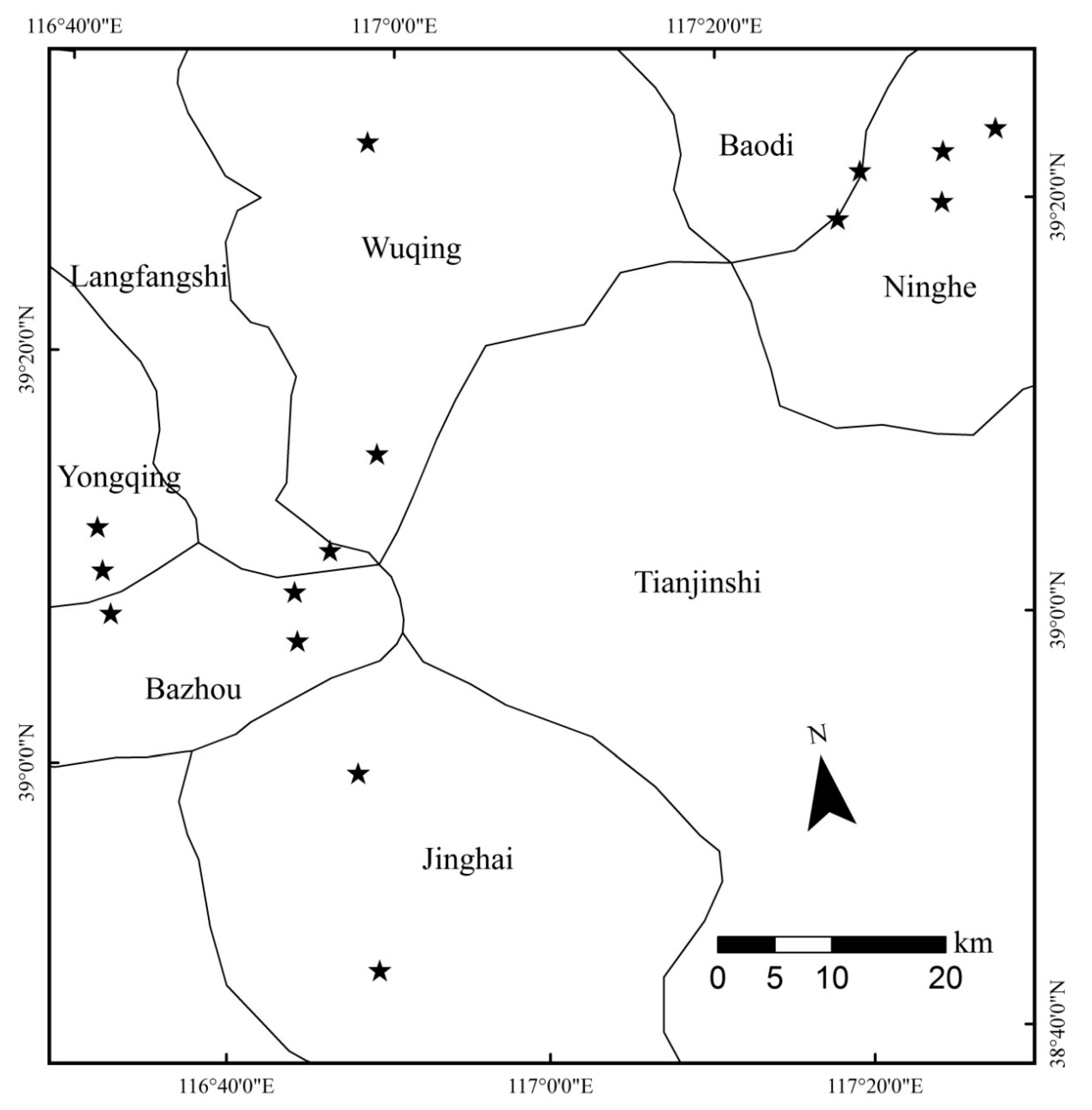
2.1. Study Locations
2.2. Sampling Method
2.3. Land Use Data and Analysis
2.4. Statistical Analysis
3. Results
3.1. Abundance of A. lucorum
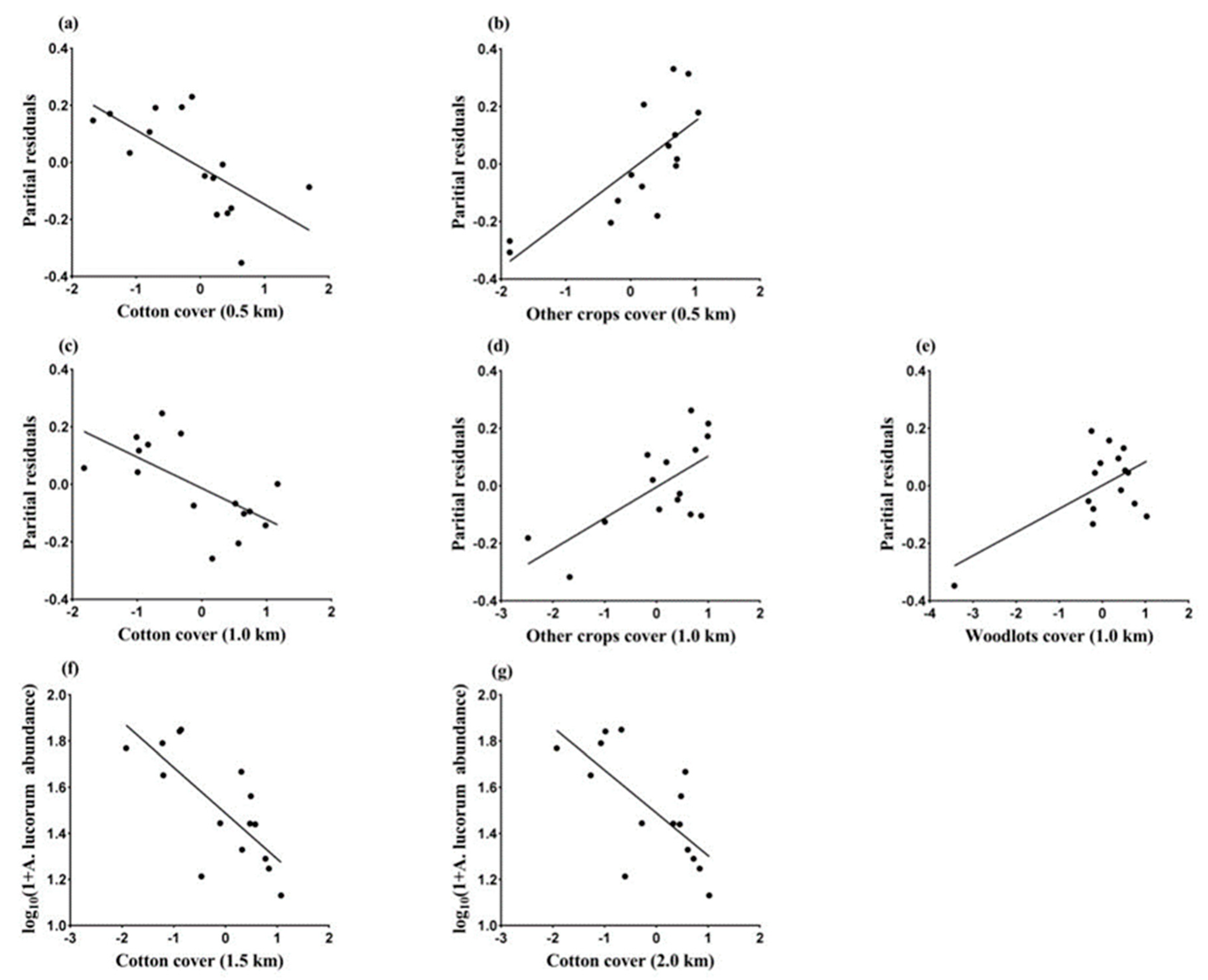
3.2. Landscape Effects on Abundance of A. lucorum in July
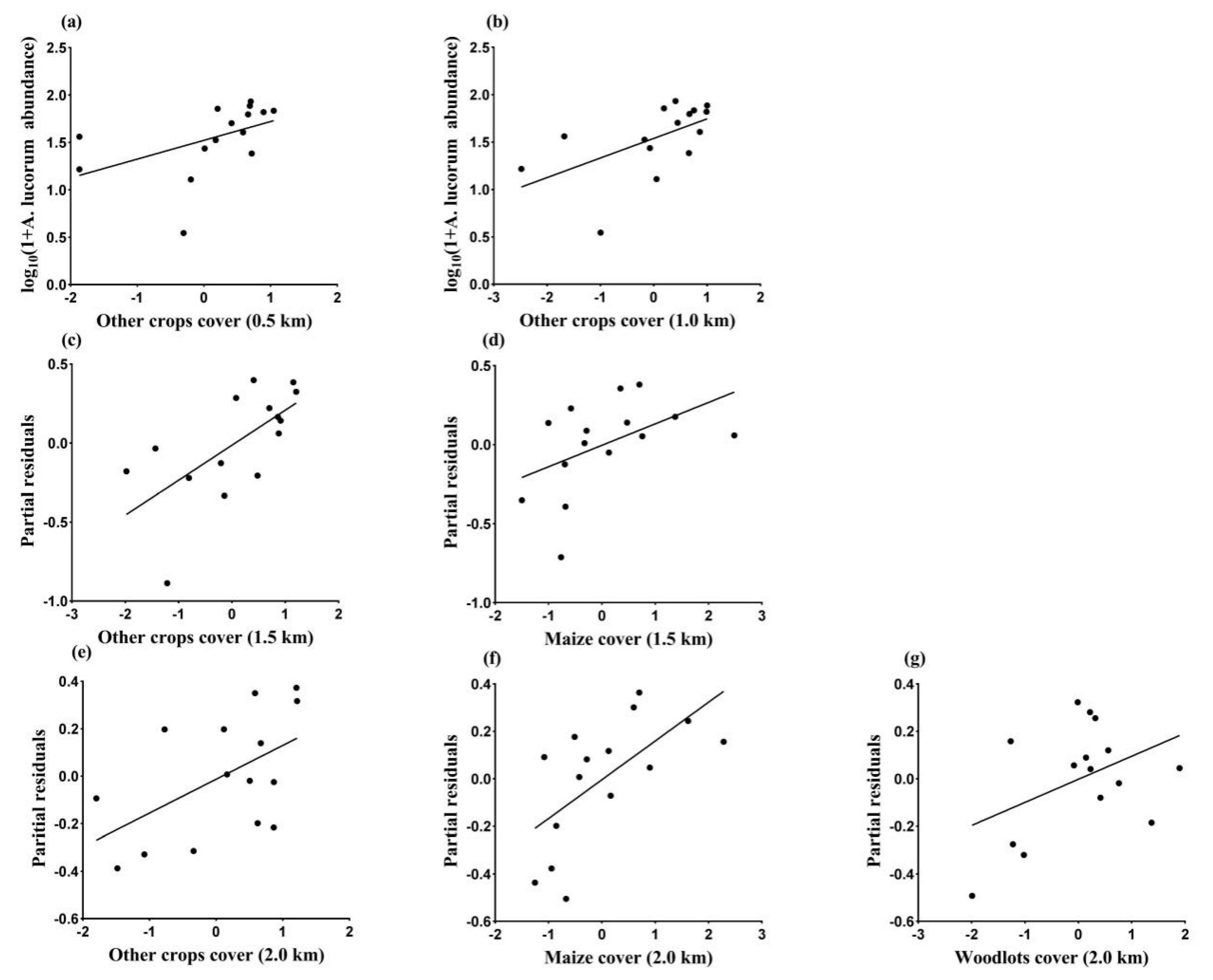
3.3. Landscape Effects on Abundance of A. lucorum in August
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bianchi, F.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. Lond. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Merenlender, A.M. Landscapes that work for biodiversity and people. Science 2018, 362, eaau6020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Ann. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin–containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, M.; Landis, D.; Gratton, C.; DiFonzo, C.; O’neal, M.; Chacon, J.; Wayo, M.; Schmidt, N.; Mueller, E.; Heimpel, G. Landscape diversity enhances biological control of an introduced crop pest in the north-central USA. Ecol. Appl. 2009, 19, 143–154. [Google Scholar] [CrossRef]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Xia, B.; Li, P.; Feng, H.Q.; Wyckhuys, K.A.; Guo, Y.Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Guo, Y.Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef]
- Sheng, J.; Gao, F.; Andile, M.; Wang, L.Y.; Sandhu, H.S.; Ou Yang, F.; Zhao, Z.H. Crop Diversity and Land Simplification Effects on Pest Damage in Northern China. Ann. Entomol. Soc. Am. 2017, 110, 91–96. [Google Scholar] [CrossRef]
- Kennedy, G.G.; Storer, N.P. Life systems of polyphagous arthropod pests in temporally unstable cropping systems. Ann. Rev. Entomol. 2000, 45, 467–493. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Goodell, P.B.; Ellers-Kirk, C.; Larocque, G.; Dutilleul, P.; Naranjo, S.E.; Ellsworth, P.C. Effects of local and landscape factors on population dynamics of a cotton pest. PLoS ONE 2012, 7, e39862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellhorn, N.A.; Gagic, V.; Bommarco, R. Time will tell: Resource continuity bolsters ecosystem services. Trends Ecol. Evol. 2015, 30, 524–530. [Google Scholar] [CrossRef]
- Tsafack, N.; Alignier, A.; Head, G.P.; Kim, J.H.; Goulard, M.; Menozzi, P.; Ouin, A. Landscape effects on the abundance and larval diet of the polyphagous pest Helicoverpa armigera in cotton fields in North Benin. Pest Manag. Sci. 2015, 72, 1613–1626. [Google Scholar] [CrossRef]
- Sivakoff, F.S.; Rosenheim, J.A.; Dutilleul, P.; Carrière, Y. Influence of the surrounding landscape on crop colonization by a polyphagous insect pest. Entomol. Exp. Appl. 2013, 149, 11–21. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.; Broyles, M.; Jones, G. Moth responses to sympathetic hedgerow management in temperate farmland. Agric. Ecosyst. Environ. 2019, 270–271, 55–64. [Google Scholar] [CrossRef]
- Carriere, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Natl. Acad. Sci. USA 2003, 100, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Wu, K.M. Mirid Bugs in China: Pest Status and Management Strategies. Outlooks Pest Manag. 2011, 22, 248–252. [Google Scholar] [CrossRef]
- Pan, H.S.; Lu, Y.H.; Wyckhuys, K.A.G.; Wu, K.M. Preference of a polyphagous mirid bug, Apolygus lucorum (Meyer-Dür) for flowering host plants. PLoS ONE 2013, 8, e68980. [Google Scholar] [CrossRef]
- Lu, Y.H.; Wu, K.M.; Guo, Y.Y. Flight Potential of Lygus lucorum (Meyer-Du¨r) (Heteroptera: Miridae). Environ. Entomol. 2007, 36, 1007–1013. [Google Scholar] [PubMed]
- Fu, X.W.; Liu, H.Q.; Li, C.; Lu, Y.H.; Li, Y.H.; Wu, K.M. Seasonal Migration of Apolygus lucorum (Hemiptera: Miridae) Over the Bohai Sea in Northern China. J. Econ. Entomol. 2014, 107, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.S.; Tena, A.; Xiu, C.; Liu, B.; Lu, Y.H.; Desneux, N. Floral feeding increases diet breadth in a polyphagous mirid. J. Pest Sci. 2019, 92, 1089–1100. [Google Scholar] [CrossRef]
- Li, Y.F.; Dang, Z.H.; Gao, Z.L.; Wang, J.Q.; Yuan, L.B.; Shen, Y.Y.; Yuan, W.L.; Pan, W.L. Distribution of Apolygus lucorum on different host plants in Hebei Province. Plant Prot. 2009, 35, 118–121. [Google Scholar]
- Lu, Y.H.; Wu, K.M.; Wyckhuys, K.A.G.; Guo, Y.Y. Overwintering hosts of Apolygus lucorum (Hemiptera: Miridae) in northern China. Crop Prot. 2010, 29, 1026–1033. [Google Scholar] [CrossRef]
- Luo, S.P.; Naranjo, S.E.; Wu, K.M. Biological control of cotton pests in China. Biol. Control 2014, 68, 6–14. [Google Scholar] [CrossRef]
- Li, J.H.; Yang, F.; Wang, Q.; Pan, H.S.; Yuan, H.B.; Lu, Y.H. Predation by generalist arthropod predators on Apolygus lucorum (Hemiptera: Miridae): Molecular gut-content analysis and field-cage assessment. Pest Manag. Sci. 2017, 73, 628–635. [Google Scholar] [CrossRef]
- Zhang, T. The Study of the Extraction, Identification and Application of Sex Pheromone Produced by Apolygus Lucorum. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2011. [Google Scholar]
- Zhang, T.; Mei, X.D.; Zhang, X.F.; Lu, Y.H.; Ning, J.; Wu, K.M. Identification and field evaluation of the sex pheromone of Apolygus lucorum (Hemiptera: Miridae) in China. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef]
- Lu, Y.H.; Wu, K.M. Biology and Control of Cotton Mirids; Golden Shield Press: Beijing, China, 2008. [Google Scholar]
- Yang, L.; Zhang, Q.; Liu, B.; Zeng, Y.D.; Pan, Y.F.; Li, M.L.; Lu, Y.H. Mixed effects of landscape complexity and insecticide use on ladybeetle abundance in wheat fields. Pest Manag. Sci. 2019, 75, 1638–1645. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop Version 10.2; Environmental Systems Research Institute: Redlands, CA, USA, 2013. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Soc. Meth. Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Grueber, C.; Nakagawa, S.; Laws, R.; Jamieson, I. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2017. Available online: https://www.R-project.org/ (accessed on 15 January 2020).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version1.40.0. 2017. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 January 2020).
- Bivand, R.; Piras, G. Comparing Implementations of Estimation Methods for Spatial Econometrics. J. Stat. Soft. 2015, 63, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.Z.; Baker, G. Spatial and temporal dynamics of Helicoverpa armigera (Lepidoptera, Noctuidae) in contrasting agricultural landscapes in northwestern China. Int. J. Pest Manag. 2013, 59, 25–34. [Google Scholar] [CrossRef]
- Downes, S.; Kriticos, D.; Parry, H.; Paull, C.; Schellhorn, N.; Zalucki, M.P. A perspective on management of Helicoverpa armigera: Transgenic Bt cotton, IPM, and landscapes. Pest Manag. Sci. 2017, 73, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lu, Y.H.; van der Werf, W.; Huang, J.; Wu, F.; Zhou, K.; Deng, X.; Jiang, Y.; Wu, K.; Rosegrant, M.W. Multidecadal, county-level analysis of the effects of land use, Bt cotton, and weather on cotton pests in China. Proc. Natl. Acad. Sci. USA 2018, 115, E7700–E7709. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Qi, F.J.; Zhang, Y.J. Integrated Management of Diseases and Insect Pests in Cotton; Golden Shield Press: Beijing, China, 2010. [Google Scholar]
- Abel, C.A.; Snodgrass, G.L.; Jackson, R.; Allen, C. Oviposition and development of the tarnished plant bug (Heteroptera: Miridae) on field maize. Environ. Entomol. 2010, 39, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Z.B.; Jaworski, C.C.; Lu, Y.H.; Ye, L.F.; Wu, K.M.; Desneux, N. Maize fields are a potential sink for an outbreaking mirid bug pest in Chinese Bt-cotton agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Z.B. Temporal and Spatial Distribution, Life Table Analysis and Population Simulation in Apolygus lucorum (Meyer-Dur) (Heteroptera: Miridae). Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2011. [Google Scholar]
- Geng, H.H.; Pan, H.S.; Lu, Y.H.; Yang, Y.Z. Nymphal and adult performance of Apolygus lucorum (Hemiptera: Miridae) on a preferred host plant, mungbean Vigna radiata. Appl. Entomol. Zool. 2012, 47, 191–197. [Google Scholar] [CrossRef]
- Dong, J.W.; Pan, H.S.; Lu, Y.H.; Yang, Y.Z. Nymphal performance correlated with adult preference for flowering host plants in a polyphagous mirid bug, Apolygus lucorum (Heteroptera: Miridae). Arthropod-Plant Inter. 2013, 7, 83–91. [Google Scholar] [CrossRef]
- Pan, H.S.; Liu, B.; Lu, Y.H. Host-plant switching promotes the population growth of Apolygus lucorum: Implications for laboratory rearing. B Entomol. Res. 2019, 109, 309–315. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Scales (km) | Model | K | logLik | AICc | ΔAICc | Weight | Adjusted R2 |
|---|---|---|---|---|---|---|---|
| 0.5 | Cotton + Other crops | 4 | 10.00 | −7.99 | 0.00 | 0.43 | 0.67 |
| Cotton + Other crops + Woodlots | 5 | 12.17 | −7.66 | 0.33 | 0.36 | 0.73 | |
| Cotton + Dwellings + Other crops | 5 | 10.72 | −4.76 | 3.23 | 0.08 | 0.67 | |
| Cotton + Dwellings + Other crops + Woodlots | 6 | 13.38 | −4.26 | 3.73 | 0.07 | 0.75 | |
| Cotton + Maize + Other crops | 5 | 10.37 | −4.08 | 3.92 | 0.06 | 0.66 | |
| 1.0 | Cotton + Other crops + Woodlots | 5 | 12.57 | −8.47 | 0.00 | 0.46 | 0.74 |
| Cotton + Maize + Other crops + Woodlots | 6 | 14.97 | −7.45 | 1.02 | 0.28 | 0.79 | |
| Cotton + Other crops | 4 | 9.02 | −6.04 | 2.42 | 0.14 | 0.62 | |
| Maize + Other crops + Woodlots | 5 | 11.2 | −5.74 | 2.72 | 0.12 | 0.74 | |
| 1.5 | Cotton | 3 | 6.74 | −5.30 | 0.00 | 0.33 | 0.53 |
| Cotton + Woodlots | 4 | 8.46 | −4.93 | 0.37 | 0.27 | 0.59 | |
| Cotton + Other crops | 4 | 7.82 | −3.65 | 1.65 | 0.14 | 0.56 | |
| Cotton + Dwellings | 4 | 7.52 | −3.04 | 2.26 | 0.11 | 0.54 | |
| Cotton + Other crops + Woodlots | 5 | 9.24 | −1.80 | 3.50 | 0.06 | 0.60 | |
| Cotton + Maize | 4 | 6.83 | −1.67 | 3.63 | 0.05 | 0.49 | |
| Other crops | 3 | 4.75 | −1.33 | 3.97 | 0.04 | 0.38 | |
| 2.0 | Cotton | 3 | 5.61 | −3.04 | 0.00 | 0.44 | 0.45 |
| Cotton + Woodlots | 4 | 6.5 | −1.00 | 2.04 | 0.16 | 0.47 | |
| Cotton + Other crops | 4 | 6.24 | −0.49 | 2.55 | 0.12 | 0.45 | |
| Cotton + Dwellings | 4 | 5.88 | 0.25 | 3.29 | 0.08 | 0.43 | |
| Woodlots | 3 | 3.83 | 0.52 | 3.56 | 0.07 | 0.30 | |
| Cotton + Maize | 4 | 5.61 | 0.78 | 3.82 | 0.06 | 0.40 | |
| Other crops | 3 | 3.7 | 0.79 | 3.82 | 0.06 | 0.29 |
| Scales (km) | Variable | Estimate | z Value | Pr (>|z|) | Relative Importance |
|---|---|---|---|---|---|
| 0.5 | Intercept | 1.511 | 39.271 | < 0.001 *** | |
| Cotton | −0.222 | 2.676 | 0.007 ** | 1.00 | |
| Dwellings | −0.085 | 1.027 | 0.304 | 0.15 | |
| Maize | −0.067 | 0.669 | 0.503 | 0.06 | |
| Other crops | 0.318 | 3.853 | < 0.001 *** | 1.00 | |
| Woodlots | 0.134 | 1.726 | 0.084 | 0.43 | |
| 1.0 | Intercept | 1.511 | 41.921 | < 0.001 *** | |
| Cotton | −0.209 | 2.468 | 0.014 * | 0.88 | |
| Maize | 0.192 | 1.774 | 0.076 | 0.40 | |
| Other crops | 0.252 | 3.105 | 0.002 ** | 1.00 | |
| Woodlots | 0.227 | 2.111 | 0.035* | 0.86 | |
| 1.5 | Intercept | 1.511 | 32.500 | < 0.001 *** | |
| Cotton | −0.325 | 2.926 | 0.003 ** | 0.96 | |
| Dwellings | 0.100 | 1.029 | 0.303 | 0.11 | |
| Maize | −0.037 | 0.349 | 0.727 | 0.05 | |
| Other crops | 0.172 | 1.247 | 0.212 | 0.24 | |
| Woodlots | 0.151 | 1.536 | 0.125 | 0.33 | |
| 2.0 | Intercept | 1.511 | 28.997 | < 0.001 *** | |
| Cotton | −0.311 | 2.587 | 0.010 ** | 0.86 | |
| Dwellings | 0.067 | 0.593 | 0.553 | 0.08 | |
| Maize | −0.002 | 0.014 | 0.989 | 0.06 | |
| Other crops | 0.178 | 1.193 | 0.233 | 0.19 | |
| Woodlots | 0.187 | 1.318 | 0.187 | 0.23 |
| Scales (km) | Model | K | logLik | AICc | ΔAICc | Weight | Adjusted R2 |
|---|---|---|---|---|---|---|---|
| 0.5 | Other crops | 3 | −4.04 | 16.30 | 0.00 | 0.30 | 0.17 |
| Null | 2 | −5.99 | 16.99 | 0.68 | 0.21 | 0.00 | |
| Maize | 3 | −5.3 | 18.78 | 2.48 | 0.09 | 0.02 | |
| Dwellings | 3 | −5.32 | 18.82 | 2.52 | 0.09 | 0.02 | |
| Dwellings + Other crops | 4 | −3.74 | 19.48 | 3.18 | 0.06 | 0.14 | |
| Maize + Other crops | 4 | −3.81 | 19.62 | 3.31 | 0.06 | 0.13 | |
| Cotton | 3 | −5.77 | 19.72 | 3.42 | 0.05 | −0.05 | |
| Cotton + Other crops | 4 | −3.93 | 19.86 | 3.55 | 0.05 | 0.11 | |
| Woodlots | 3 | −5.89 | 19.96 | 3.66 | 0.05 | −0.06 | |
| Other crops + Woodlots | 4 | −4.05 | 20.10 | 3.79 | 0.04 | 0.10 | |
| 1.0 | Other crops | 3 | −3.12 | 14.42 | 0.00 | 0.29 | 0.27 |
| Maize + Other crops | 4 | −1.65 | 15.31 | 0.89 | 0.19 | 0.35 | |
| Maize | 3 | −.19 | 16.56 | 2.15 | 0.10 | 0.15 | |
| Null | 2 | −5.99 | 16.99 | 2.57 | 0.08 | 0.00 | |
| Cotton + Other crops | 4 | −2.61 | 17.22 | 2.80 | 0.07 | 0.26 | |
| Cotton | 3 | −4.66 | 17.50 | 3.08 | 0.06 | 0.10 | |
| Maize + Woodlots | 4 | −2.79 | 17.57 | 3.16 | 0.06 | 0.24 | |
| Maize + Other crops + Woodlots | 5 | −0.46 | 17.58 | 3.16 | 0.06 | 0.39 | |
| Dwellings + Other crops | 4 | −3.11 | 18.23 | 3.81 | 0.04 | 0.21 | |
| Other crops + Woodlots | 4 | −3.12 | 18.23 | 3.81 | 0.04 | 0.21 | |
| 1.5 | Maize + Other crops | 4 | −0.48 | 12.97 | 0.00 | 0.19 | 0.44 |
| Other crops | 3 | −2.4 | 12.99 | 0.02 | 0.19 | 0.33 | |
| Maize + Other crops + Woodlots | 5 | 1.6 | 13.46 | 0.49 | 0.15 | 0.54 | |
| Maize + Woodlots | 4 | −0.9 | 13.80 | 0.83 | 0.13 | 0.41 | |
| Cotton | 3 | −3.12 | 14.42 | 1.46 | 0.09 | 0.27 | |
| Dwellings + Maize + Other crops | 5 | 0.81 | 15.04 | 2.07 | 0.07 | 0.49 | |
| Cotton + Other crops | 4 | −1.72 | 15.44 | 2.47 | 0.06 | 0.34 | |
| Other crops + Woodlots | 4 | −2.31 | 16.61 | 3.64 | 0.03 | 0.29 | |
| Dwellings + Maize + Woodlots | 5 | 0.02 | 16.63 | 3.67 | 0.03 | 0.43 | |
| Cotton + Maize | 4 | −2.33 | 16.65 | 3.69 | 0.03 | 0.28 | |
| Dwellings + Other crops | 4 | −2.4 | 16.80 | 3.84 | 0.03 | 0.28 | |
| 2.0 | Maize + Other crops + Woodlots | 5 | 3.4 | 9.87 | 0.00 | 0.34 | 0.64 |
| Maize + Other crops | 4 | 1.05 | 9.90 | 0.03 | 0.34 | 0.54 | |
| Other crops | 3 | −1.45 | 11.08 | 1.20 | 0.19 | 0.41 | |
| Maize + Woodlots | 4 | −0.29 | 12.59 | 2.71 | 0.09 | 0.45 | |
| Dwellings + Maize + Other crops | 5 | 1.4 | 13.86 | 3.99 | 0.05 | 0.53 |
| Scales (km) | Variable | Estimate | z Value | Pr (>|z|) | Relative Importance |
|---|---|---|---|---|---|
| 0.5 | Intercept | 1.548 | 15.126 | < 0.001 *** | |
| Cotton | −0.108 | 0.494 | 0.622 | 0.10 | |
| Dwellings | −0.185 | 0.843 | 0.399 | 0.15 | |
| Maize | 0.184 | 0.824 | 0.410 | 0.14 | |
| Other crops | 0.346 | 1.674 | 0.094 | 0.51 | |
| Woodlots | −0.059 | 0.264 | 0.792 | 0.09 | |
| 1.0 | Intercept | 1.548 | 16.624 | < 0.001 *** | |
| Cotton | −0.231 | 1.063 | 0.288 | 0.13 | |
| Dwellings | −0.016 | 0.077 | 0.939 | 0.04 | |
| Maize | 0.360 | 1.521 | 0.128 | 0.41 | |
| Other crops | 0.391 | 2.025 | 0.043 * | 0.70 | |
| Woodlots | 0.234 | 0.847 | 0.397 | 0.16 | |
| 1.5 | Intercept | 1.548 | 18.607 | < 0.001 *** | |
| Cotton | −0.349 | 1.575 | 0.115 | 0.18 | |
| Dwellings | 0.198 | 0.860 | 0.390 | 0.13 | |
| Maize | 0.405 | 1.864 | 0.062 | 0.60 | |
| Other crops | 0.407 | 2.149 | 0.032 * | 0.72 | |
| Woodlots | 0.369 | 1.604 | 0.109 | 0.34 | |
| 2.0 | Intercept | 1.548 | 21.308 | < 0.001 *** | |
| Dwellings | 0.123 | 0.650 | 0.516 | 0.05 | |
| Maize | 0.345 | 2.186 | 0.029 * | 0.81 | |
| Other crops | 0.452 | 2.643 | 0.008 ** | 0.91 | |
| Woodlots | 0.332 | 1.850 | 0.069 | 0.43 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Yang, L.; Pan, Y.; Zhang, Q.; Yuan, H.; Lu, Y. Landscape Effects on the Abundance of Apolygus lucorum in Cotton Fields. Insects 2020, 11, 185. https://doi.org/10.3390/insects11030185
Li M, Yang L, Pan Y, Zhang Q, Yuan H, Lu Y. Landscape Effects on the Abundance of Apolygus lucorum in Cotton Fields. Insects. 2020; 11(3):185. https://doi.org/10.3390/insects11030185
Chicago/Turabian StyleLi, Minlong, Long Yang, Yunfei Pan, Qian Zhang, Haibin Yuan, and Yanhui Lu. 2020. "Landscape Effects on the Abundance of Apolygus lucorum in Cotton Fields" Insects 11, no. 3: 185. https://doi.org/10.3390/insects11030185