Effects of Diaphorina citri Population Density on Daily Timing of Vibrational Communication Calls: Potential Benefits in Finding Forage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioassay Arena
2.2. D. citri Handling Procedures: Treatments at P5 and P25 Densities
2.3. Recording Procedures: Treatments at P5 and P25 Densities
2.4. D. citri Handling Procedures: Flush Shoot Aggregation Sex Ratio Tests
2.5. D. citri Handling Procedures: Flush Age Tests
2.6. Statistical Analyses
3. Results
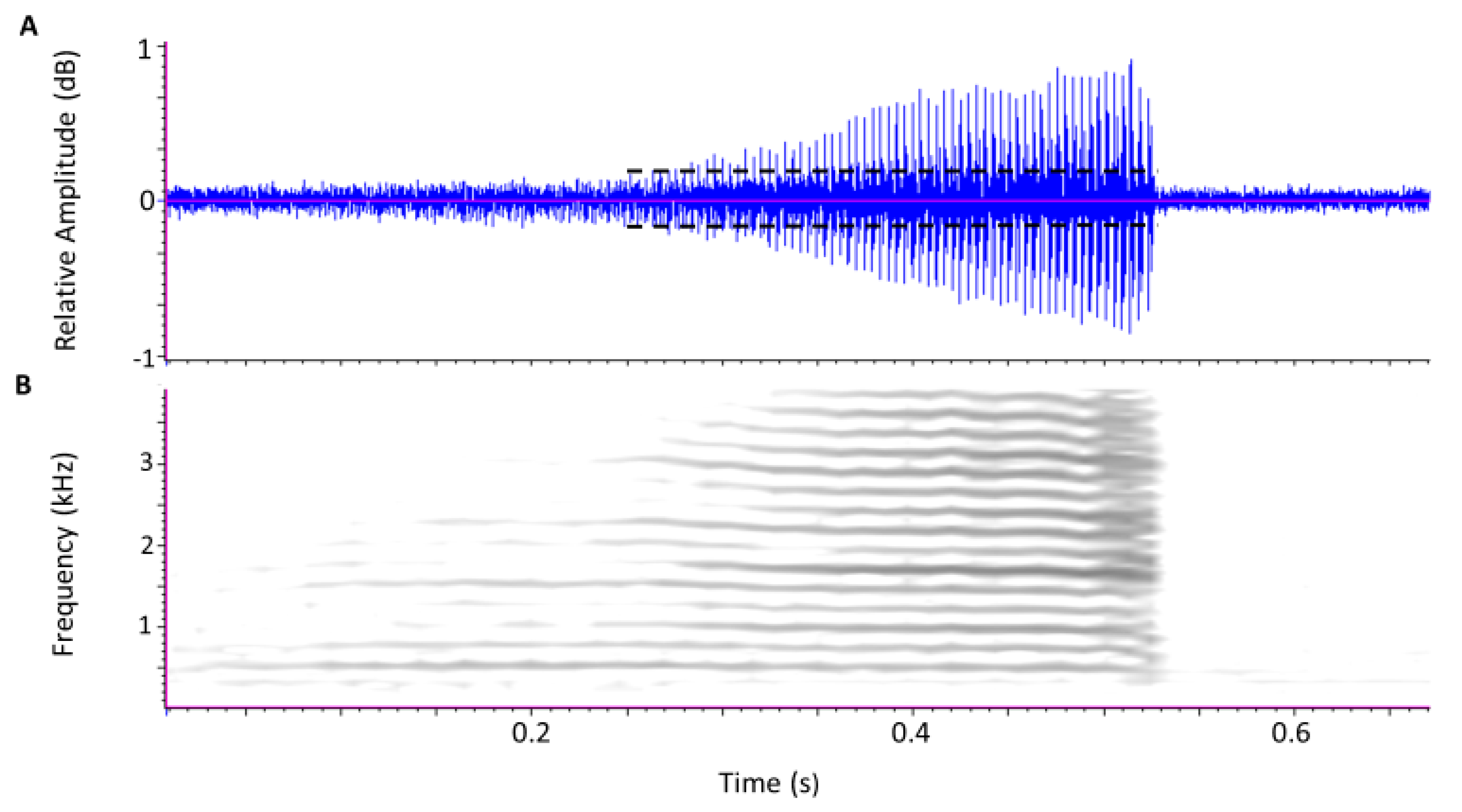
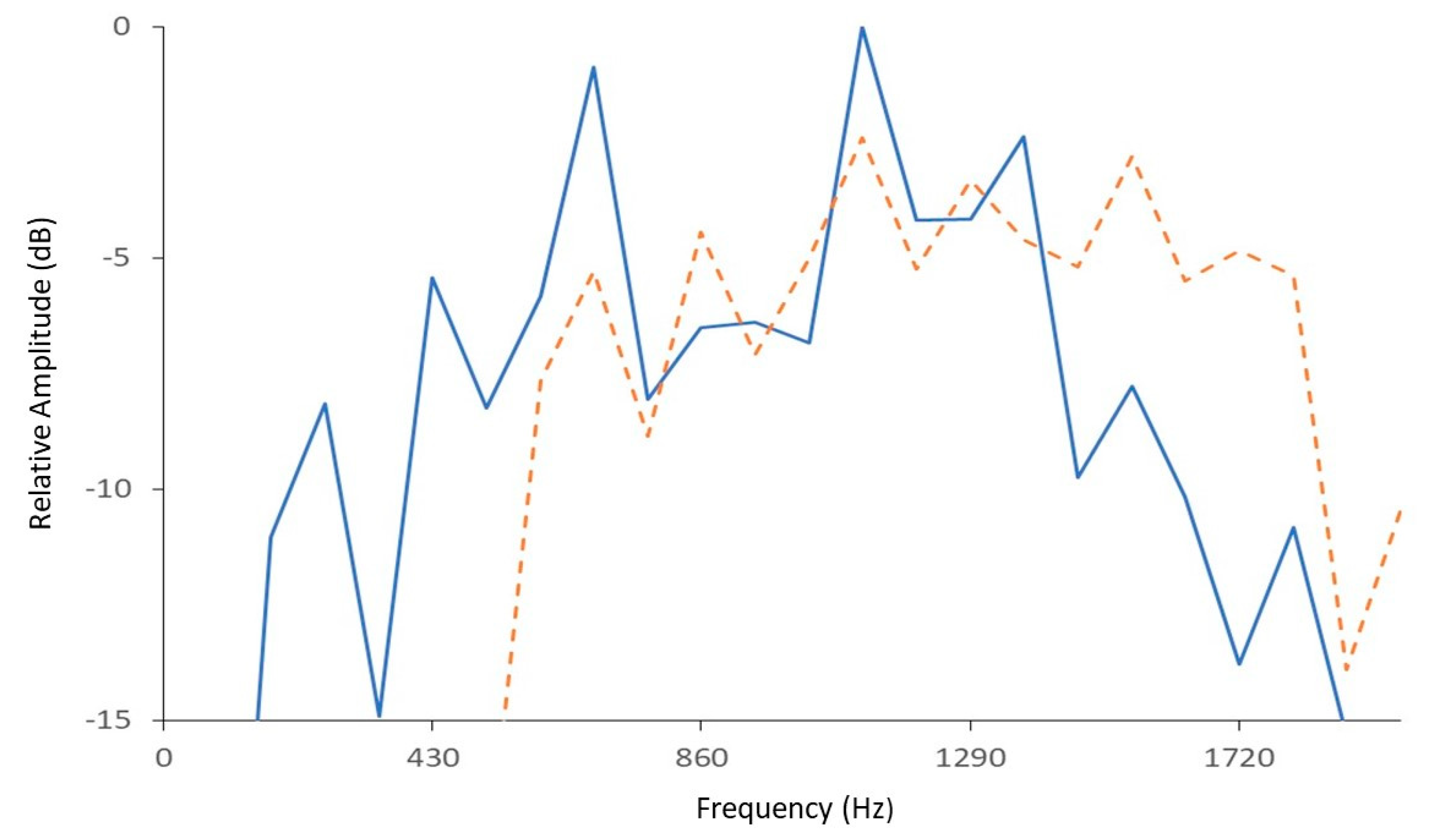
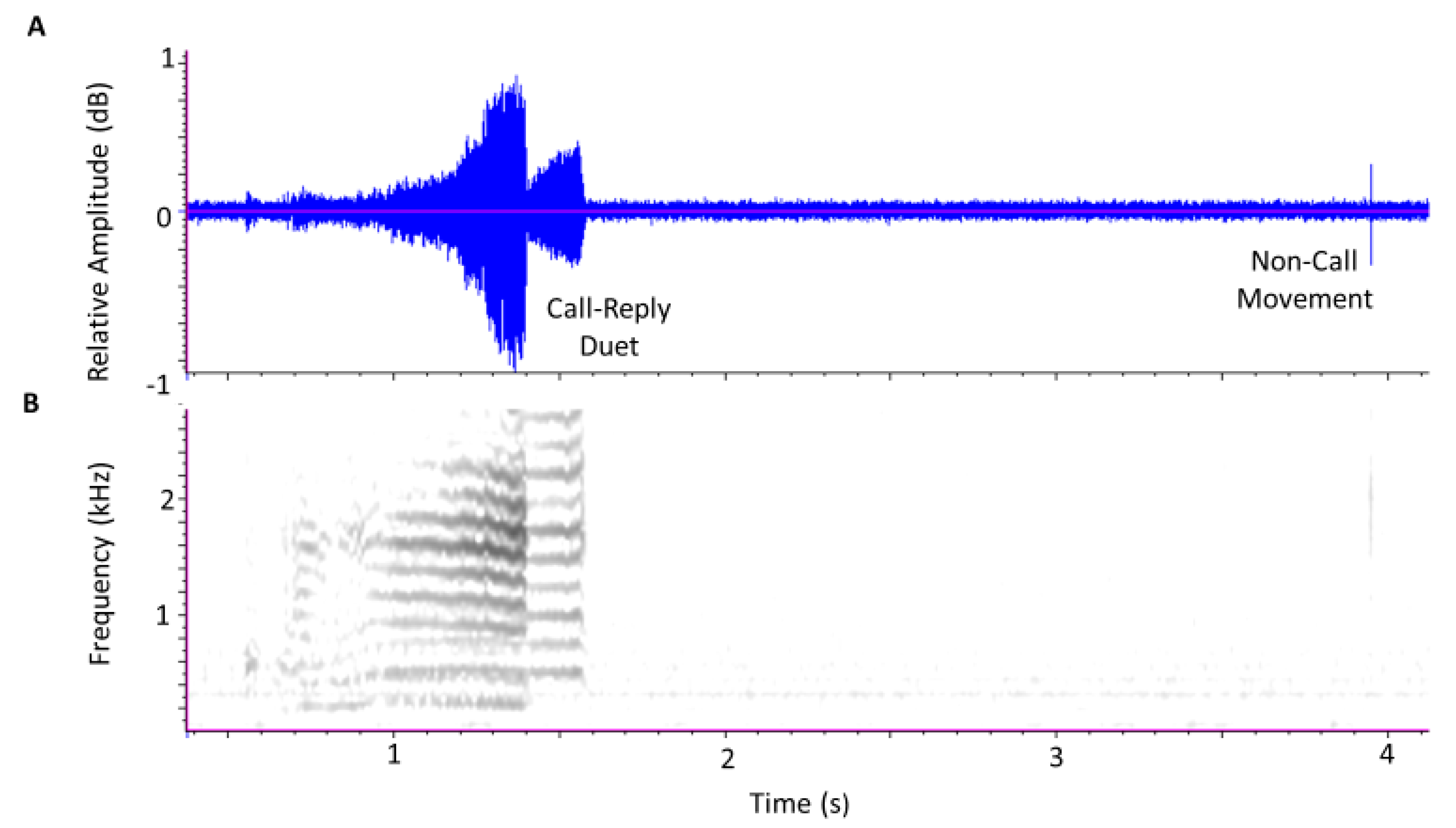
3.1. Calls and Movement Vibrations in Treatments at P5 and P25 Densities
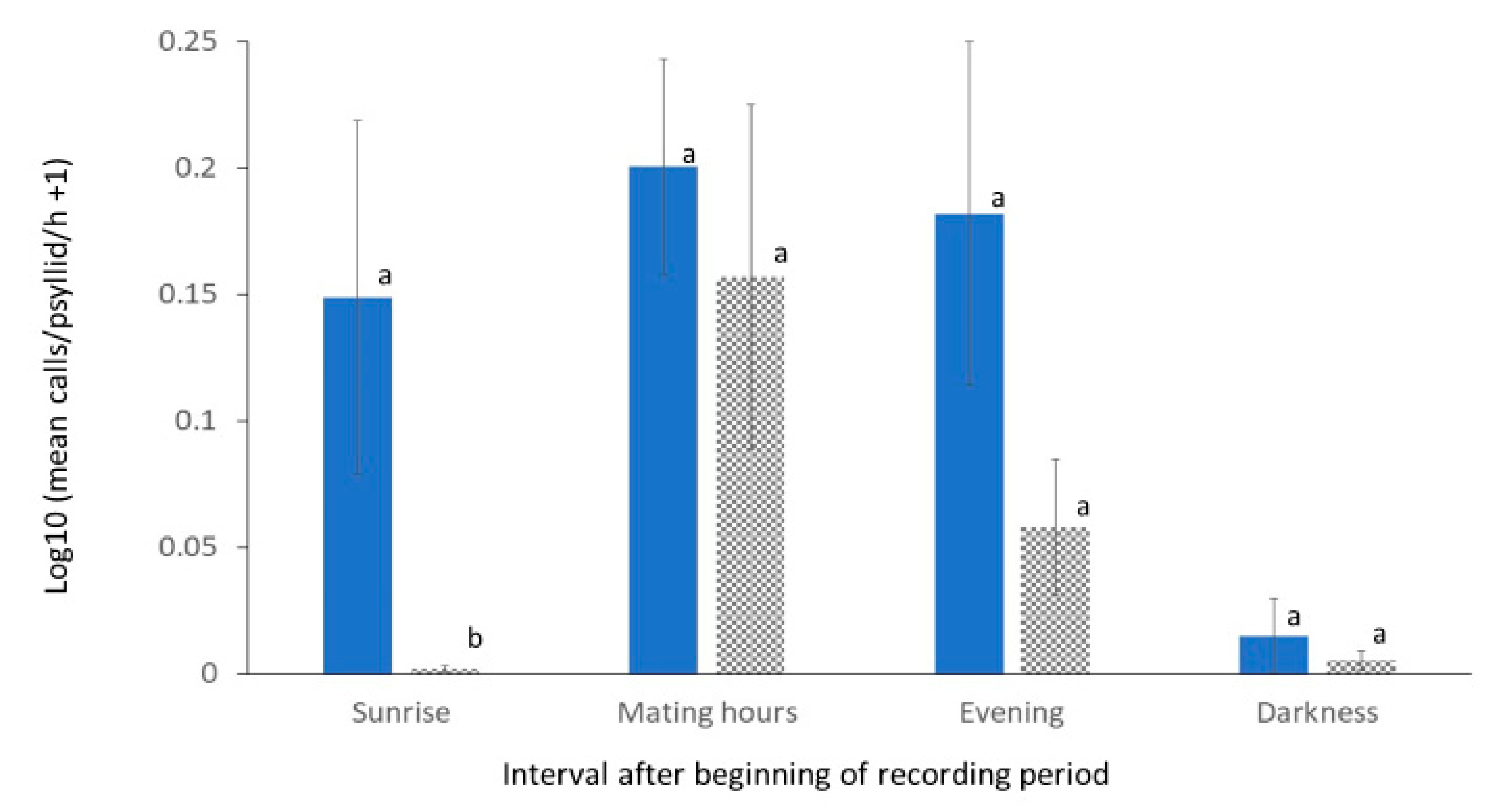
3.2. Calling Rates in Treatments at P5 and P25 Densities
3.3. Distributions of Females and Males in Flush Aggregations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hall, D.G.; Richardson, M.L.; Ammar, E.-D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2012, 146, 207–223. [Google Scholar] [CrossRef]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenninger, E.J.; Stelinski, L.L.; Hall, D.G. Roles of olfactory cues, visual cues, and mating status in orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to four different host plants. Environ. Entomol. 2009, 38, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, X.; Hoffman, M.; Coy, M.R.; Stelinski, L.L.; Pelz-Stelinski, K.S. Infection of an insect vector with a bacterial plant pathogen increases its propensity for dispersal. PLoS ONE 2015, 10, e0129373. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.G.; Albrigo, L.G. Estimating the relative abundance of flush shoots in citrus with implications on monitoring insects associated with flush. HortScience 2007, 42, 364–368. [Google Scholar] [CrossRef] [Green Version]
- Sétamou, M.; Flores, D.; French, J.V.; Hall, D.G. Dispersion patterns and sampling plans for Diaphorina citri (Hemiptera: Psyllidae) in citrus. J. Econ. Entomol. 2008, 101, 1478–1487. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Da Graça, J.V.; Bassanezi, R.B. Citrus huanglongbing: The pathogen and its impact. Plant Health Prog. 2007. [Google Scholar] [CrossRef] [Green Version]
- Pregmon, E.A.; Lujo, S.; Norton, K.; Hartman, E.; Rohde, B.; Zagvazdina, N.; Mankin, R.W. A “walker” tool to place Diaphorina citri (Hemiptera: Liviidae) adults at predetermined sites for bioassays of behavior in citrus (Sapindales: Rutaceae) trees. Fla. Entomol. 2016, 99, 308–310. [Google Scholar] [CrossRef]
- Sétamou, M.; Sanchez, A.; Patt, J.M.; Nelson, S.D.; Jifon, J.; Louzada, E.S. Diurnal patterns of flight activity and effects of light on host finding behavior of the Asian citrus psyllid. J. Insect. Behav. 2012, 25, 264–276. [Google Scholar] [CrossRef]
- Paris, T.M.; Allan, S.A.; Udell, B.J.; Stansly, P.A. Wavelength and polarization affect phototaxis of the Asian citrus psyllid. Insects 2017, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Ammar, E.-D.; Hall, D.G.; Shatters, R.G., Jr. Stylet morphometrics and citrus leaf vein structure in relation to feeding behavior of the Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing bacterium. PLoS ONE 2013, 8, e59914. [Google Scholar] [CrossRef] [Green Version]
- Sétamou, M.; Simpson, C.R.; Alabi, O.J.; Nelson, S.D.; Telagamsetty, S.; Jifon, J.L. Quality matters: Influences of citrus flush physicochemical characteristics on population dynamics of the Asian citrus psyllid (Hemiptera: Liviidae). PLoS ONE 2016, 11, e0168997. [Google Scholar] [CrossRef] [PubMed]
- Patt, J.M.; Sétamou, M. Responses of the Asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environ. Entomol. 2010, 39, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.; Jiang, Q.; Zhang, B.; Xu, C.; Chen, K. Citrus leaf volatiles as affected by developmental stage and genetic type. Int. J. Mol. Sci. 2013, 14, 17744–17766. [Google Scholar] [CrossRef] [PubMed]
- Martini, X.; Kuhns, E.M.; Hoyte, A.; Stelinski, L.L. Plant volatiles and density-dependent conspecific female odors are used by Asian citrus psyllid to evaluate host suitability on a spatial scale. Arth. Plant Interact. 2014, 8, 453–460. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Hall, D.G.; Mankin, R.W. Vibrational communication between the sexes in Diaphorina citri (Hemiptera: Psyllidae). Ann. Entomol. Soc. Am. 2009, 102, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Zagvazdina, N.Y.; Paris, T.M.; Udell, B.J.; Stanislauskas, M.; McNeill, S.; Allan, S.A.; Mankin, R.W. Effects of atmospheric pressure trends on calling, mate-seeking, and phototaxis of Diaphorina citri (Hemiptera: Liviidae). Ann. Entomol. Soc. Am. 2015, 108, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Lujo, S.; Hartman, E.; Norton, K.; Pregmon, E.; Rohde, B.; Mankin, R.W. Disrupting mating behavior of Diaphorina citri (Liviidae). J. Econ. Entomol. 2016, 109, 2373–2379. [Google Scholar] [CrossRef] [Green Version]
- Mann, R.S.; Rouseff, R.L.; Smoot, J.; Rao, N.; Meyer, W.L.; Lapointe, S.L.; Robbins, P.S.; Cha, D.; Linn, C.E.; Webster, F.X.; et al. Chemical and behavioral analysis of the cuticular hydrocarbons from Asian citrus psyllid, Diaphorina Citri. Insect Sci. 2013, 20, 367–378. [Google Scholar] [CrossRef]
- Miklas, N.; Lasnier, T.; Renou, M. Male bugs modulate pheromone emission in response to vibratory signals from conspecifics. J. Chem. Ecol. 2003, 29, 561–574. [Google Scholar] [CrossRef]
- Hall, D.G.; Hentz, M.G.; Patt, J.M. Behavioral assay on Asian citrus psyllid attraction to orange jasmine. J. Insect Behav. 2015, 28, 555–568. [Google Scholar] [CrossRef]
- Rohde, B.; Paris, T.M.; Heatherington, E.M.; Hall, D.G.; Mankin, R.W. Responses of Diaphorina citri (Hemiptera: Psyllidae) to conspecific vibrational signals and synthetic mimics. Ann. Entomol. Soc. Am. 2013, 106, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Cocroft, R.B. Vibrational communication and the ecology of group-living herbivorous insects. Am. Zool. 2001, 41, 1215–1221. [Google Scholar] [CrossRef] [Green Version]
- Cocroft, R.B. Vibrational communication facilitates cooperative foraging in a phloem-feeding insect. Proc. Roy. Soc. B Biol. Sci. 2005, 272, 1023–1029. [Google Scholar] [CrossRef]
- Mazzoni, V.; Prešern, J.; Lucchi, A.; Virant-Doberlet, M. Reproductive strategy of the Nearctic leafhopper Scaphoideus titanus Ball (Hemiptera: Cicadellidae). Bull. Entomol. Res. 2009, 99, 401–413. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Muhamad, R.; Omar, D.; Majid, D.L.; Gnanasegaram, M. Influence of male age on producing rival calls in brown planthopper Nilaparvata lugens Stål (Homoptera: Delphacidae). J. Entomol. Zool. Stud. 2016, 4, 163–166. [Google Scholar]
- Čokl, A.; Kosi, A.Z.; Laumann, R.A.; Virant-Doberlet, M. Female competition for availability of males in insects: The Nezara viridula (Linnaeus, 1758) model. Insect Sci. 2019. [Google Scholar] [CrossRef]
- Tishechkin, D.Y. Vibratory communication in Psylloidea (Hemiptera). In Insect Sounds and Communication: Physiology, Behavior, Ecology, and Evolution; Drosopoulos, S., Claridge, M.F., Eds.; CRC Press: New York, NY, USA, 2005; pp. 357–363. [Google Scholar]
- Wenninger, E.J.; Hall, D.G. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Fla. Entomol. 2007, 90, 715–722. [Google Scholar] [CrossRef]
- Mankin, R.W.; Rohde, B.; McNeill, S. Vibrational duetting mimics to trap and disrupt mating of the devastating Asian citrus psyllid insect pest. Proc. Meet. Acoust. 2016, 25, 010006. [Google Scholar]
- Paris, T.M.; Rohde, B.B.; Allan, S.A.; Mankin, R.W.; Stansly, P.A. Synchronized rearing of mated and unmated Diaphorina citri (Hemiptera: Liviidae) of known age. Fla. Entomol. 2013, 96, 1631–1634. [Google Scholar] [CrossRef]
- Charif, R.A.; Waack, A.M.; Strickman, L.M. Raven Pro 1.3 User’s Manual; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2008. [Google Scholar]
- Mankin, R.W.; Stanaland, D.; Haseeb, M.; Rohde, B.; Menocal, O.; Carrillo, D. Assessment of plant structural characteristics, health, and ecology using bioacoustic tools. Proc. Mtg. Acoust. 2018, 33, 010003. [Google Scholar]
- Mankin, R.W.; Brandhorst-Hubbard, J.; Flanders, K.L.; Zhang, M.; Crocker, R.L.; Lapointe, S.L.; McCoy, C.W.; Fisher, J.R.; Weaver, D.K. Eavesdropping on insects hidden in soil and interior structures of plants. J. Econ. Entomol. 2000, 93, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Mankin, R.W.; Anderson, J.B.; Mizrach, A.; Epsky, N.D.; Shuman, D.; Heath, R.R.; Mazor, M.; Hetzroni, A.; Grinshpun, J.; Taylor, P.W.; et al. Broadcasts of wingfanning vibrations recorded from calling male Ceratitis capitata (Diptera: Tephritidae) increase captures of females in traps. J. Econ. Entomol. 2004, 97, 1299–1309. [Google Scholar] [CrossRef]
- Cifuentes-Arenas, J.C.; de Goes, A.; de Miranda, M.P.; Beattie, G.A.C.; Lopes, S.A. Citrus flush shoot ontogeny modulates biotic potential of Diaphorina citri. PLoS ONE 2018, 13, e0190563. [Google Scholar] [CrossRef]
- SAS Institute Inc. Base SAS 9.4 Procedures Guide, Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Mankin, R.W.; Hodges, R.D.; Schal, C.; Pereira, R.M.; Koehler, P.G. Acoustic indicators for targeted detection of stored product and urban insect pests by inexpensive infrared, acoustic, and vibrational detection of movement. J. Econ. Entomol. 2010, 103, 1135–1143. [Google Scholar] [CrossRef]
- Paris, T.M.; Croxton, S.D.; Stansly, P.A.; Allan, S.A. Temporal response and attraction of Diaphorina citri to visual stimuli. Ent. Exp. Appl. 2015, 155, 137–147. [Google Scholar] [CrossRef]
- Bedoya, C.L.; Brockerhoff, E.G.; Hayes, M.; Pawson, S.M.; Nanar-Rodriguez, A.; Nelson, X.J. Acoustic communication of the red-haired bark beetle Hylurgus ligniperda. Physiol. Entomol. 2019. [Google Scholar] [CrossRef]
- Hunt, R.E.; Nault, L.R. Roles of interplant movement, acoustic communication and phototaxis in mate-location of the leafhopper Graminella nigrifrons. Beh. Ecol. Sociobiol. 1991, 28, 315–320. [Google Scholar] [CrossRef]
- Catling, H.D. Distribution of the psyllid vectors of citrus greening disease, with notes on the biology and bionomics of Diaphorina citri. FAO Plant Protect. Bull. 1970, 18, 8–15. [Google Scholar]
- Pande, Y.D. Biology of citrus psylla, Diaphorina citri Kuw. (Hemiptera: Psyllidae). Isr. J. Entomol. 1971, 6, 307–311. [Google Scholar]
- Alves, G.R.; Diniz, A.J.F.; Parra, J.R.P. Biology of the huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. J. Econ. Entomol. 2014, 107, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Patt, J.M.; Stockton, D.; Meikle, W.G.; Sétamou, M.; Mafra-Neto, A.; Adamczyk, J.J. Innate and conditioned responses to chemosensory and visual cues in asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae), vector of huanglongbing pathogens. Insects 2014, 5, 921–941. [Google Scholar] [CrossRef] [PubMed]
- Stockton, D.G.; Martini, X.; Stelinski, L.L. Male psyllids differentially learn in the context of copulation. Insects 2017, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockton, D.G.; Pescitelli, L.E.; Martini, X.; Stelinski, L.L. Female mate preference in an invasive phytopathogen vector: How learning may influence mate choice and fecundity in Diaphorina citri. Entomol. Exp. Appl. 2017, 164, 16–26. [Google Scholar] [CrossRef]
- Li, X.; Ishimoto, H.; Kamikouchi, A. Auditory experience controls the maturation of song discrimination and sexual response in Drosophila. eLife 2018, 7, e34348. [Google Scholar] [CrossRef] [PubMed]
- Yadav, C.; Guedes, R.N.C.; Matheson, S.M.; Timbers, T.A.; Yack, J.E. Invitation by vibration: Recruitment to feeding shelters in social caterpillars. Behav. Ecol. Sociobiol. 2017, 71, 51. [Google Scholar] [CrossRef]
- Čokl, A.A.; Millar, J.G. Manipulation of insect signaling for monitoring and control of insect pests. In Biorational Control of Arthropod Pests; Ishaaya, I., Horowitz, A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 279–316. [Google Scholar]
- Hofstetter, R.W.; Dunn, D.D.; McGuire, R.; Potter, K.A. Using acoustic technology to reduce bark beetle reproduction. Pest Manag. Sci. 2014, 70, 24–27. [Google Scholar] [CrossRef]
- Polajnar, J.; Eriksson, A.; Lucchi, A.; Anfora, G.; Virant-Doberlet, M.; Mazzoni, V. Manipulating behaviour with substrate-borne vibrations-potential for insect pest control. Pest Manag. Sci. 2016, 71, 15–23. [Google Scholar] [CrossRef]
- Mazzoni, V.; Polajnar, J.; Baldini, M.; Rossi Stacconi, M.V.; Anfora, G.; Guidetti, R.; Maistrello, L. Use of substrate-borne vibrational signals to attract the brown marmorated stink bug, Halyomorpha halys. J. Pest. Sci. 2017, 90, 1219–1229. [Google Scholar] [CrossRef]
- Nieri, R.; Mazzoni, V.; Gordon, S.D.; Krugner, R. Mating behavior and vibrational mimicry in the glassy-winged sharpshooter, Homalodisca vitripennis. J. Pest Sci. 2017, 90, 887–899. [Google Scholar] [CrossRef]
- Mazzoni, V.; Gordon, S.D.; Nieri, R.; Krugner, R. Design of a candidate vibrational signal for mating disruption against the glassy-winged sharpshooter, Homalodisca vitripennis. Pest Man. Sci. 2017, 73, 2328–2333. [Google Scholar] [CrossRef] [PubMed]
- Nieri, R.; Mazzoni, V. Vibrational mating disruption of Empoasca vitis by natural or artificial disturbance noises. Pest Man. Sci. 2019, 75, 1065–1073. [Google Scholar] [CrossRef]
- Gordon, S.D.; Sandoval, N.; Mazzoni, V.; Krugner, R. Mating interference of glassy-winged sharpshooters, Homalodisca vitripennis. Entomol. Exp. Appl. 2017, 164, 27–34. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Killiny, N. Better together: Association with ‘Candidatus Liberibacter Asiaticus’ increases the reproductive fitness of its insect vector, Diaphorina citri (Hemiptera: Liviidae). Ann. Entomol. Soc. Am. 2016, 109, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | Mean Square | F | p |
|---|---|---|---|---|
| Psyllid density | 1 | 0.224 | 5.06 | 0.031 * |
| Calling period interval | 3 | 0.172 | 4.76 | 0.039 * |
| Interval–density interaction | 3 | 0.036 | 1.00 | 0.390 |
| Sex | Areal Density (No. psyllids/cm2) On Flush Type | Score | U | p | |
|---|---|---|---|---|---|
| Young (n = 12) | Mature (n = 23) | ||||
| Female | 2.72 ± 0.42 | 0.41 ± 0.07 | 280 | 4.29 | <0.0001 |
| Male | 1.60 ± 0.29 | 0.47 ± 0.08 | 252 | 3.19 | 0.0013 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mankin, R.W.; Patel, R.; Grugnale, M.; Jetter, E. Effects of Diaphorina citri Population Density on Daily Timing of Vibrational Communication Calls: Potential Benefits in Finding Forage. Insects 2020, 11, 182. https://doi.org/10.3390/insects11030182
Mankin RW, Patel R, Grugnale M, Jetter E. Effects of Diaphorina citri Population Density on Daily Timing of Vibrational Communication Calls: Potential Benefits in Finding Forage. Insects. 2020; 11(3):182. https://doi.org/10.3390/insects11030182
Chicago/Turabian StyleMankin, Richard W., Rikin Patel, Mason Grugnale, and Ethan Jetter. 2020. "Effects of Diaphorina citri Population Density on Daily Timing of Vibrational Communication Calls: Potential Benefits in Finding Forage" Insects 11, no. 3: 182. https://doi.org/10.3390/insects11030182