Male-Produced (−)-δ-Heptalactone, Pheromone of Fruit Fly Rhagoletis batava (Diptera: Tephritidae), a Sea Buckthorn Berries Pest

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
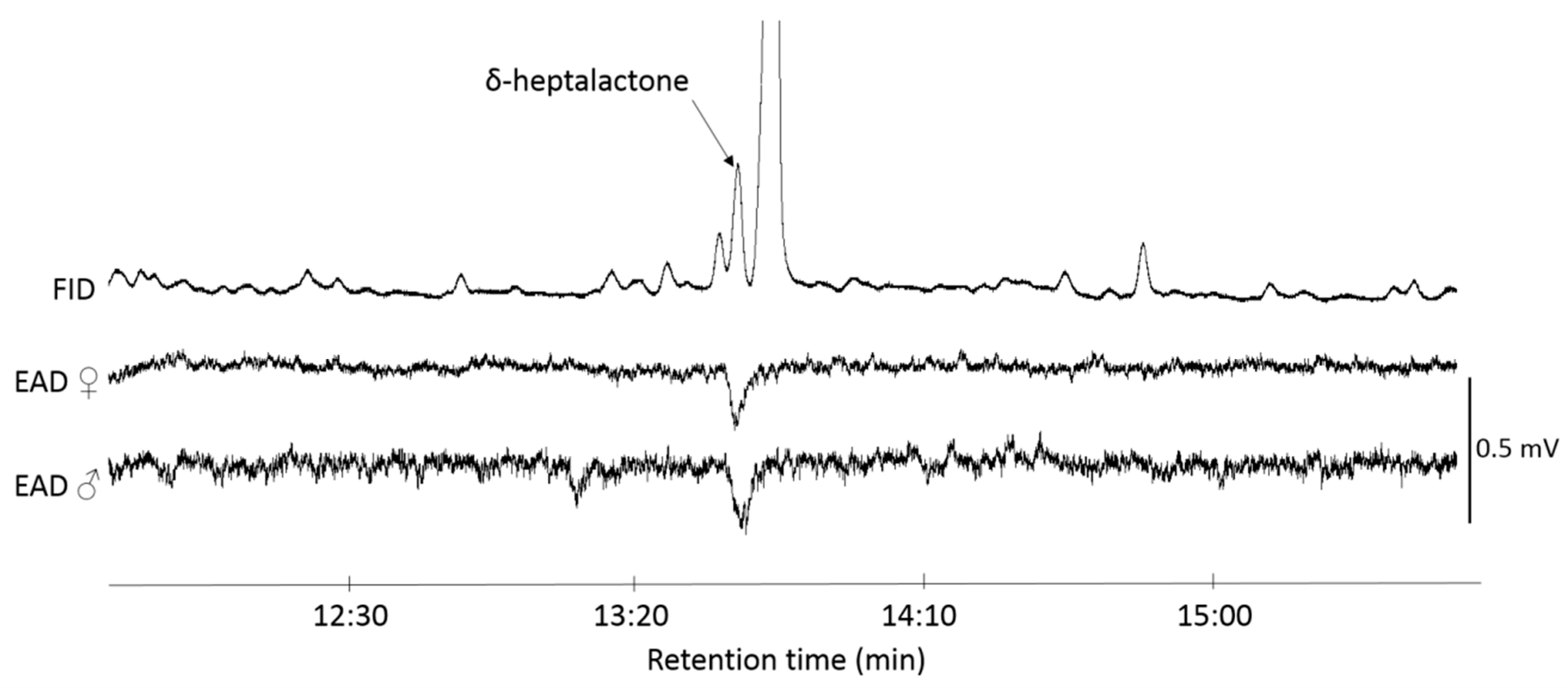
2.2. Sampling of R. batava-Produced Volatiles
2.3. GC-EAD Detection
2.4. GC-MS Analysis
2.5. Synthesis
2.6. EAG Dose-Response
2.7. Field Assay
2.8. Statistical Analysis
3. Results
3.1. EAD Active Compound
3.2. EAD Active Enantiomer of δ-Heptalactone
3.3. Active Enantiomer of δ-Heptalactone in R. batava
3.4. Dose Response
3.5. Behavioural Test under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shamanskaya, L.D. Bioecology of the sea-buckthorn fly (Rhagoletis batava obscuriosa Kol.) and pest control treatment in Altai. In Proceedings of Producing Sea Buckthorn of High Quality; Natural Resources Institute Finland: Naantali, Finland, 2015; pp. 7–20. [Google Scholar]
- Shalkevich, M.S.; Koltun, N.Y.; Pleskatsevich, R.I. Sea buckthorn pests and diseases in Belarus. In Proceedings of Producing Sea Buckthorn of High Quality; Natural Resources Institute Finland: Naantali, Finland, 2015; pp. 83–86. [Google Scholar]
- Li, T.S.C.; Schroeder, W.R. Sea buckthorn (Hippophae rhamnoides L.): A multipurpose plant. Hort. Technol. 1996, 6, 370–380. [Google Scholar]
- Hakeem, K.; Ozturk, M.; Altay, V.; Letchamo, W.; Musayev, M.; Mamedov, N. An alternative potential natural genetic resource: Sea buckthorn [Elaeagnus rhamnoides (syn.: Hippophae rhamnoides)]. In Global Perspectives on Underutilized Crops; Ozturk, M., Hakeem, K.H., Ashraf, M., Ahmad, M.S.A., Eds.; Springer: Cham, Switzerland, 2018; pp. 25–82. [Google Scholar] [CrossRef]
- Small, E.; Catling, P.M.; Li, T.S.C. Blossoming treasures of biodiversity: 5. Sea Buckthorn (Hippophae rhamnoides)—An ancient crop with modern virtues. Biodiversity 2002, 3, 25–27. [Google Scholar] [CrossRef]
- Ruan, C.J.; da Silva, J.A.T.; Jin, H.; Li, H.; Li, D.Q. Research and biotechnology in sea buckthorn (Hippophae spp.). Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 47–60. [Google Scholar]
- Bal, L.M.; Meda, V.; Naik, S.N.; Satya, S. Sea buckthorn berries: A potential source of valuable nutrients for nutraceuticals and cosmoceuticals. Food Res. Int. 2011, 44, 1718–1727. [Google Scholar] [CrossRef]
- Kaur, T.; Singh, G.; Kapoor, D.N. A review on pharmacognostic, phytochemical and pharmacological data of various species of Hippophae (Sea buckthorn). Int. J. Green Pharm. 2017, 11, s62–s75. [Google Scholar]
- Eagle, D. Are quality standards necessary for sea buckthorn—A business perspective. In Proceedings of Producing Sea Buckthorn of High Quality; Natural Resources Institute Finland: Naantali, Finland, 2015; pp. 44–48. [Google Scholar]
- Brūvelis, A. Experiences about sea buckthorn cultivation and harvesting in Latvia. In Proceedings of Producing Sea Buckthorn of High Quality; Natural Resources Institute Finland: Naantali, Finland, 2015; pp. 36–41. [Google Scholar]
- Toth, M.; Lerche, S.; Holz, U.; Kerber, A.; Henning, R.; Voigt, E.; Kelemen, D. Addition of synthetic feeding attractant increases catches of Rhagoletis batava Hering and Carpomyia schineri Loew. in fluorescent yellow sticky traps. Acta Phytopathologica et Entomologica Hungarica 2016, 51, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Cheng, T.M.; Li, S.S.; Su, Z.; Wei, J.R. Attractants for Rhagoletis batava obseuriosa, a fruit fly pest of sea buckthorn. Int. J. Pest Manag. 2019, 65, 324–331. [Google Scholar] [CrossRef]
- Papaj, D.R.; Garcia, J.M.; AlonsoPimentel, H. Marking of host fruit by male Rhagoletis boycei cresson flies (Diptera: Tephritidae) and its effect on egg-laying. J. Insect Behav. 1996, 9, 585–598. [Google Scholar] [CrossRef]
- Sarles, L.; Boullis, A.; Fassotte, B.; Lognay, G.; Verhaeghe, A.; Francis, F.; Verheggen, F.J. Identification of walnut husk (Juglans regia L.) volatiles and the behavioural response of the invasive Walnut Husk Fly, Rhagoletis completa Cresson. Pest Manag. Sci. 2017, 73, 2100–2104. [Google Scholar] [CrossRef]
- Sarles, L.; Fassotte, B.; Boullis, A.; Lognay, G.; Verhaeghe, A.; Marko, I.; Verheggen, F.J. Improving the monitoring of the walnut husk fly (Diptera: Tephritidae) using male-produced lactones. J. Econ. Entomol. 2018, 111, 2032–2037. [Google Scholar] [CrossRef]
- Raptopoulos, D.; Haniotakis, G.; Koutsaftikis, A.; Kelly, D.; Mavraganis, V. Biological-activity of chemicals identified from extracts and volatiles of male Rhagoletis cerasi. J. Chem. Ecol. 1995, 21, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.A.; Weatherly, C.A.; Armstrong, D.W. Chiral Gas Chromatography. In Chiral Analysis, 2nd ed.; Polavarapu, P.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 468–505. [Google Scholar] [CrossRef]
- Chalk, S.J. Retention index, I in column chromatography. In IUPAC. Compendium of Chemical Terminology, the “Gold Book”; McNaught, A.D., Wilkinson, A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1997. [Google Scholar] [CrossRef]
- Reid, B.T.; Mailyan, A.K.; Zakarian, A. Total synthesis of (+)-guadinomic acid via hydroxyl-directed guanidylation. J. Org. Chem. 2018, 83, 9492–9496. [Google Scholar] [CrossRef] [PubMed]
- Soai, K.; Yokoyama, S.; Hayasaka, T.; Ebihara, K. Catalytic asymmetric-synthesis of alkyl substituted lactones by enantioselective and chemoselective alkylation of formylesters with dialkylzincs using n,n-dibutylnorephedrine. Chem. Lett. 1988, 843–846. [Google Scholar] [CrossRef]
- Aleknavičius, D.; Būda, V. Trapping peculiarities, flight and mating dynamics of sea buckhorn fruit fly (Rhagoletis batava) in Lithuania. Zemdirbyste 2019, 106, 81–86. [Google Scholar] [CrossRef]
- Bewick, V.; Cheek, L.; Ball, J. Statistics review 9: One-way analysis of variance. Crit. Care 2004, 8, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Daane, K.M.; Canale, A.; Niu, C.Y.; Messing, R.H.; Vargas, R.I. Sexual communication and related behaviours in Tephritidae: Current knowledge and potential applications for Integrated Pest Management. J. Pest Sci. 2014, 87, 385–405. [Google Scholar] [CrossRef]
- Mori, K. Significance of chirality in pheromone science. Bioorganic Med. Chem. 2007, 15, 7505–7523. [Google Scholar] [CrossRef]
- Pask, G.M.; Romaine, I.M.; Zwiebel, L.J. The molecular receptive range of a lactone receptor in Anopheles gambiae. Chem. Senses 2013, 38, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, Z.S.; Butt, T.M. Preferences of the peripheral olfactory system of western flower thrips, Frankliniella occidentalis towards stereoisomers of common plant volatiles. Chemoecology 2015, 25, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Yu, J.; Yang, Y.Q.; Wan, X.C. Response to enantiomers of (Z3Z9)-6,7-epoxy-octadecadiene, sex pheromone component of Ectropis obliqua Prout (Lepidoptera: Geometridae): Electroantennagram test, field trapping and in silico study. Fla. Entomol. 2019, 102, 549–554. [Google Scholar] [CrossRef]
- Leal, W.S. Enantiomeric anosmia in scarab beetles. J. Chem. Ecol. 1999, 25, 1055–1066. [Google Scholar] [CrossRef]
- Svensson, G.P.; Oleksa, A.; Gawronski, R.; Lassance, J.M.; Larsson, M.C. Enantiomeric conservation of the male-produced sex pheromone facilitates monitoring of threatened European hermit beetles (Osmoderma spp.). Entomologia Experimentalis Et Applicata 2009, 133, 276–282. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Animal Behaviour: Communication by Smell and Taste; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar] [CrossRef]
- Benelli, G.; Giunti, G.; Canale, A.; Messing, R.H. Lek dynamics and cues evoking mating behavior in tephritid flies infesting soft fruits: Implications for behavior-based control tools. Appl. Entomol. Zool. 2014, 49, 363–373. [Google Scholar] [CrossRef]
- Messina, F.J.; Subler, J.K. Conspecific and heterospecific interactions of male Rhagoletis flies (Diptera, Tephritidae) on a shared host. J. Kans. Entomol. Soc. 1995, 68, 206–213. [Google Scholar]
- Aluja, M.; Birke, A. Habitat use by adults of Anastrepha obliqua (Diptera, Tephritidae) in a mixed mango and tropical plum orchard. Ann. Entomol. Soc. Am. 1993, 86, 799–812. [Google Scholar] [CrossRef]
- Schulz, S.; Hotling, S. The use of the lactone motif in chemical communication. Nat. Prod. Rep. 2015, 32, 1042–1066. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.; Read, D.M.Y.; Gurr, G.M. Application of Chemical Cues in Arthropod Pest Management for Organic Crops. In Chemical Ecology of Insect Parasitoids; Wajnberg, E., Colazza, S., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2013; pp. 266–281. [Google Scholar] [CrossRef]
- Daniel, C. Experiences of integrated management of European Cherry Fruit Fly (Rhagoletis cerasi) and how to utilize this knowledge for Sea Buckthorn Fly (Rhagoletis batava). In Proceedings of Producing Sea Buckthorn of High Quality; Natural Resources Institute Finland: Naantali, Finland, 2015; pp. 21–30. [Google Scholar]
- Daniel, C.; Grunder, J. Integrated management of European cherry fruit fly Rhagoletis cerasi (L.): Situation in Switzerland and Europe. Insects 2012, 3, 956–988. [Google Scholar]
- Mozūraitis, R.; Aleknavičius, D.; Vepštaitė-Monstavičė, I.; Stanevičienė, R.; Noushin Emami, S.; Apšegaitė, V.; Radžiutė, S.; Blažytė-Čereškienė, L.; Servienė, E.; Būda, V. Hippophae rhamnoides berry related Pichia kudriavzevii yeast volatiles modify behaviour of Rhagoletis batava flies. J. Adv. Res. 2019. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Navarro, D.M.A.; De Aquino, N.C.; Ferreira, L.L.; Tavares, R.F.; da Silva, R.D.C.; Lima-Mendonca, A.; Vanickova, L.; Mendonca, A.L.; Do Nascimento, R.R. Identification of male-borne attractants in Anastrepha fraterculus (Diptera: Tephritidae). Chemoecology 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Lima, I.S.; House, P.E.; do Nascimento, R.R. Volatile substances from male Anastrepha fraterculus wied. (Diptera: Tephritidae): Identification and behavioural activity. J. Braz. Chem. Soc. 2001, 12, 196–201. [Google Scholar] [CrossRef]
- Caceres, C.; Segura, D.F.; Vera, M.T.; Wornoayporn, V.; Cladera, J.L.; Teal, P.; Sapountzis, P.; Bourtzis, K.; Zacharopoulou, A.; Robinson, A.S. Incipient speciation revealed in Anastrepha fraterculus (Diptera; Tephritidae) by studies on mating compatibility, sex pheromones, hybridization and cytology. Biol. J. Linn. Soc. 2009, 97, 152–165. [Google Scholar] [CrossRef]
- Rocca, J.R.; Nation, J.L.; Strekowski, L.; Battiste, M.A. Comparison of volatiles emitted by male caribbean and mexican fruit-flies. J. Chem. Ecol. 1992, 18, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Chuman, T.; Sivinski, J.; Heath, R.R.; Calkins, C.O.; Tumlinson, J.H.; Battiste, M.A.; Wydra, R.L.; Strekowski, L.; Nation, J.L. Suspensolide, a new macrolide component of male caribbean fruit-fly (Anastrepha suspensa Loew ) volatiles. Tetrahedron Lett. 1988, 29, 6561–6564. [Google Scholar] [CrossRef]
- Lu, F.; Teal, P.E.A. Sex pheromone components in oral secretions and crop of male Caribbean fruit files, Anastrepha suspensa (Loew). Arch. Insect Biochem. Physiol. 2001, 48, 144–154. [Google Scholar] [CrossRef]
- Baker, J.D.; Heath, R.R. NMR spectral assignment of lactone pheromone components emitted by caribbean and mexican fruit-flies. J. Chem. Ecol. 1993, 19, 1511–1519. [Google Scholar] [CrossRef]
- Battiste, M.A.; Strekowski, L.; Vanderbilt, D.P.; Visnick, M.; King, R.W.; Nation, J.L. Anastrephin and epianastrephin, novel lactone components isolated from the sex-pheromone blend of male caribbean and mexican fruit-flies. Tetrahedron Lett. 1983, 24, 2611–2614. [Google Scholar] [CrossRef]
- Stokes, J.B.; Uebel, E.C.; Warthen, J.D.; Jacobson, M.; Flippenanderson, J.L.; Gilardi, R.; Spishakoff, L.M.; Wilzer, K.R. Isolation and identification of novel lactones from male mexican fruit-flies. J. Agric. Food Chem. 1983, 31, 1162–1167. [Google Scholar] [CrossRef]
- Robacker, D.C.; Ingle, S.J.; Hart, W.G. Mating frequency and response to male-produced pheromone by virgin and mated females of the mexican fruit-fly (Diptera, Tephritidae). Southw. Entomol. 1985, 10, 215–221. [Google Scholar]
- Ohinata, K.; Jacobson, M.; Kobayashi, R.M.; Chambers, D.L.; Fujimoto, M.S.; Higa, H.H. Oriental fruit fly(Diptera, Tephritidae) and melon fly(Diptera, Tephritidae)—Biological and chemical studies of smoke produced by males. J. Environ. Sci. Health Part A Environ. Sci. Eng. Toxic Hazard. Subst. Control 1982, 17, 197–216. [Google Scholar] [CrossRef]
- Baker, R.; Herbert, R.H.; Grant, G.G. Isolation and identification of the sex-pheromone of the Mediterranean fruit-fly, Ceratitis capitata (Wied). J. Chem. Soc. Chem. Commun. 1985, 824–825. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catalyst | Product | Yield | ee 1 | CP 2 |
|---|---|---|---|---|
| (1R,2S)-(+)-N,N-dibutylnorephidrine | (R)-(+)-3 | 35% over 2 steps | 84% | 97% |
| (1S,2R)-(−)-N,N-dibutylnorephidrine | (S)-(−)-3 | 32% over 2 steps | 86% | 96% |
| Compound Name | CAS No. 1 | Species | Emitter | Reference |
|---|---|---|---|---|
| Lavender lactone | 1073-11-6 | Anastrepha fraterculus | M | [39] |
| Suspensolide | 111351-08-7 | Anastrepha fraterculus | M | [39,40,41] |
| Anastrepha ludens | M | [42] | ||
| Anastrepha suspensa | M | [42,43,44] | ||
| Anastrepha sp. | M | [45] | ||
| S,S-Anastrephin | 77670-94-1 | Anastrepha fraterculus | M | [39,40,41] |
| Anastrepha ludens | M | [42,46,47,48] | ||
| Anastrepha suspensa | M | [42,43,44,46,48] | ||
| Anastrepha sp. | M | [45] | ||
| S,S-Epianastrephin | 77670-93-0 | Anastrepha fraterculus | M | [39,40,41] |
| Anastrepha ludens | M | [42,46,47] | ||
| Anastrepha suspensa | M | [42,43,44,46,48] | ||
| Anastrepha sp. | M | [45] | ||
| (E)-5-(3,6-heptadienyl)-dihydro-2(3H)-furanone | 81693-14-3 | Bactrocera cucurbitae | M | [49] |
| Dihydro-3-methylfuran-2(3H)-one | 1679-47-6 | Ceratitis capitata | M | [50] |
| δ-Hexalactone | 823-22-3 | Rhagoletis completa | M | [15] |
| δ-Heptalactone | 3301-90-4 | Rhagoletis completa | M | [15] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Būda, V.; Blažytė-Čereškienė, L.; Radžiutė, S.; Apšegaitė, V.; Stamm, P.; Schulz, S.; Aleknavičius, D.; Mozūraitis, R. Male-Produced (−)-δ-Heptalactone, Pheromone of Fruit Fly Rhagoletis batava (Diptera: Tephritidae), a Sea Buckthorn Berries Pest. Insects 2020, 11, 138. https://doi.org/10.3390/insects11020138
Būda V, Blažytė-Čereškienė L, Radžiutė S, Apšegaitė V, Stamm P, Schulz S, Aleknavičius D, Mozūraitis R. Male-Produced (−)-δ-Heptalactone, Pheromone of Fruit Fly Rhagoletis batava (Diptera: Tephritidae), a Sea Buckthorn Berries Pest. Insects. 2020; 11(2):138. https://doi.org/10.3390/insects11020138
Chicago/Turabian StyleBūda, Vincas, Laima Blažytė-Čereškienė, Sandra Radžiutė, Violeta Apšegaitė, Patrick Stamm, Stefan Schulz, Dominykas Aleknavičius, and Raimondas Mozūraitis. 2020. "Male-Produced (−)-δ-Heptalactone, Pheromone of Fruit Fly Rhagoletis batava (Diptera: Tephritidae), a Sea Buckthorn Berries Pest" Insects 11, no. 2: 138. https://doi.org/10.3390/insects11020138