Evaluation of a ‘Preventative’ Strategy to Manage Spider Mites on Almond

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Effectiveness of Preventative Acaricide Applications
2.2. Natural Enemy Sampling
2.3. Spider Mite Colony to Determine Base-Line Susceptibility to Abamectin
2.4. Collection of Field Populations for Bioassays
2.5. Laboratory Bioassays
2.6. Statistical Analyses
3. Results
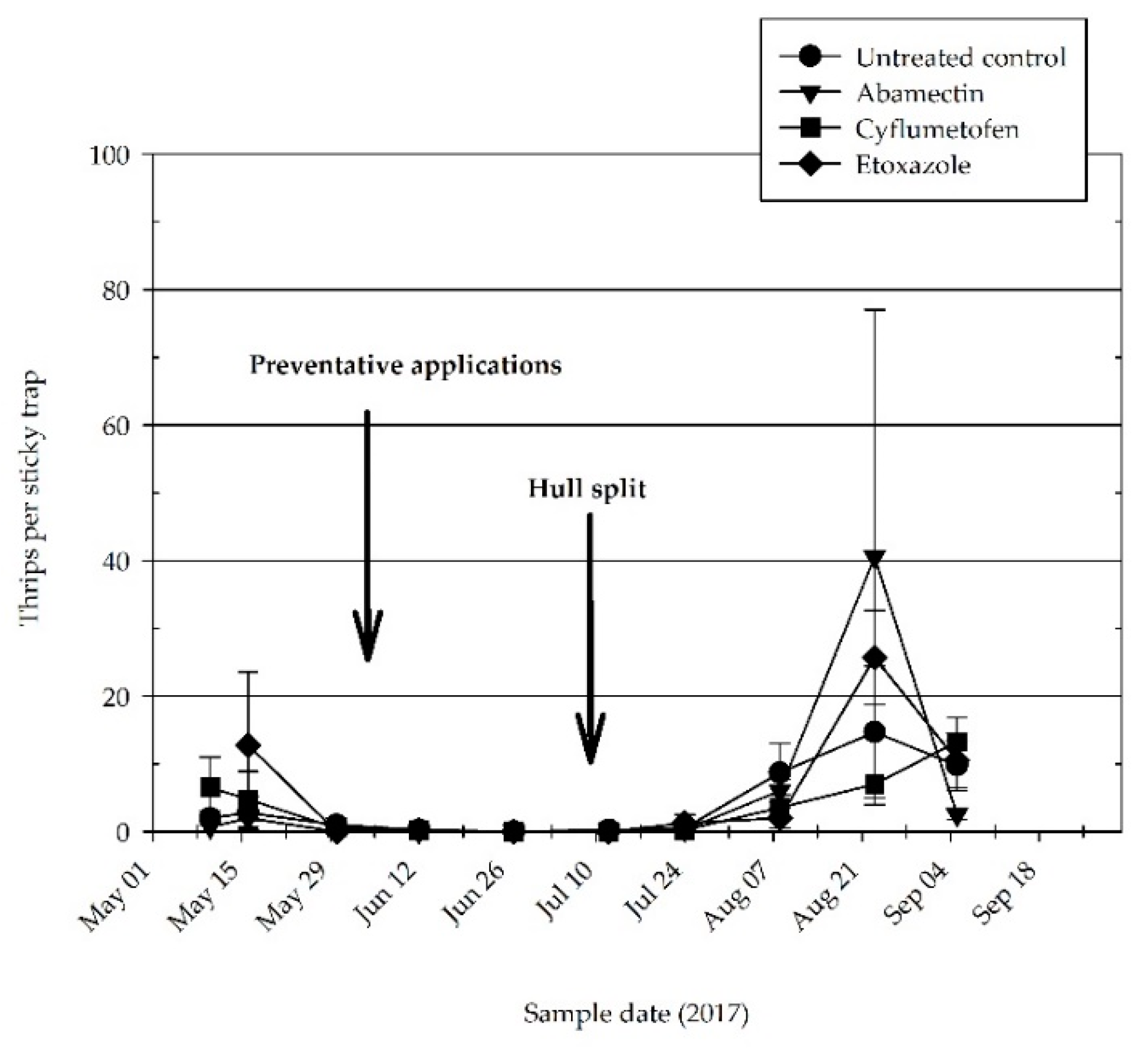
3.1. Effectiveness of Preventative Acaricide Applications
3.2. Spider Mite Predators
3.3. Susceptible Spider Mites to Abamectin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USDA. California Almond Acreage Report; USDA: Sacramento, CA, USA, 2017; p. 8.
- Zalom, F.G.; Wilson, L.T.; Hoy, M.A.; Barnett, W.; Smilanick, J.M. Sampling Tetranychus spider mites on almond. Calif. Agric. 1984, 38, 17–19. [Google Scholar]
- Welter, S.C.; Barnes, M.M.; Ting, I.P.; Hayashi, J.T. Impact of various levels of late-season spider mite (Acari: Tetranychidae) feeding damage on almond growth and yield. J. Environ. Entomol. 1984, 12, 52–55. [Google Scholar] [CrossRef]
- Youngman, R.R.; Jones, V.P.; Welter, S.C.; Barnes, M.M. Comparison of feeding damage caused by four tetranychid mite species on gas-exchange rates of almond leaves. Environ. Entomol. 1986, 15, 190–193. [Google Scholar] [CrossRef]
- Barnes, M.M.; Andrews, K.L. Effects of spider mites on almond tree growth and productivity. J. Econ. Entomol. 1978, 3, 555–558. [Google Scholar] [CrossRef]
- California Department of Pesticide Regulation. Summary of Pesticide Use Report: Indexed by Commodity; California Department of Pesticide Regulation: Sacramento, CA, USA, 2017.
- Beers, E.H.; Andersen, A.; Brown, R.D. Absorption and translaminar activity of abamectin in apple and pear foliage as determined by spider mite (Acari: Tetranychidae) mortality. J. Econ. Entomol. 1997, 90, 566–573. [Google Scholar] [CrossRef]
- Knowles, C.O. Mechanisms of resistance to acaricides. In Molecular Mechanisms of Resistance to Agrochemicals; Springer: New York, NY, USA, 1997; pp. 55–77. [Google Scholar]
- Haviland, D. Managing mites on almond with abamecin. The Almond Doctor. Doll, D., Ed.; Available online: https://thealmonddoctor.com/ (accessed on 13 August 2020).
- Vacante, V. The Handbook of Mites of Economic Plants; CAB International: Wallingford, UK, 2016; p. 872. [Google Scholar]
- Mound, L.A. Species recongnition in the genus Scolothrips (Thysanoptera, Thripidae), predators of leaf-feeding mites. Zootaxa 2011, 2797, 45–53. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute. SAS/STAT® 9.3 User’s Guide; SAS Institute Inc.: Carey, NC, USA, 2011. [Google Scholar]
- Critchlow, D.E.; Fligner, M.A. On distribution-free multiple comparisons in the one-way analysis of variance. Commun. Stat. Theory 1991, 20, 127–139. [Google Scholar]
- Robertson, J.L.; Russell, R.M.; Preisler, H.K.; Savin, N.E. Bioassays with Arthropods, 2nd ed.; CRC Press: Boca Roton, FL, USA, 2007; p. 199. [Google Scholar]
- Yan, W.Q.; Wang, J.J.; Wang, S.L. Population dynamics of the vegetable whitefly, Bemisia tabaci, and its resistance to neonicotinoid insecticides in Beijing area. Plant Prot. 2012, 38, 154–157. [Google Scholar]
- Almond Board of California. Almond Almanac 2019. Almond Board of California; Almond Board of California: Modesto, CA, USA, 2019. [Google Scholar]
- Ross, N.W.; Catlin, P.B. Rootstocks and Root Physiology; Publication, 4092; Warren, D., Kester, D., Eds.; University of California Press: Berkeley, CA, USA, 1978; pp. 46–51. [Google Scholar]
- Martin, D.E.; Kester, D. Almond Growth and Development; Publication, 4092; Warren, D., Kester, D., Eds.; University of California Press: Berkeley, CA, USA, 1978; pp. 46–51. [Google Scholar]
- Yee, W.L.; Phillips, P.A.; Faber, B.A. Effects of aerial spray volume, coverage, and abamectin on Scirtothrips perseae (Thysanoptera: Thripidae). J. Econ. Entomol. 2001, 94, 1090–2001. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Morse, J.G. Abamectin grower cooperator citrus thrips trials, 1994. In Arthropod Management Tests; Saxena, C.R., Ed.; Entomological Society of America: Lanham, MD, USA, 1997; p. 73. [Google Scholar]
- Dybas, R.A. Abamectin use in crop protection. In Ivermectin and Abemectin; Campbell, C.W., Ed.; Springer: New York, NY, USA, 1989; pp. 287–310. [Google Scholar]
- Coville, P.L.; Allen, W.W. Life table and feeding habits of Scolothrips sexmaculatus (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am. 1977, 70, 11–16. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A. Mechanisms of acaricide resistance in the two spotted spider mite Tetranychus urticae. In Biorational Control of Arthropod Pests; Ishaaya, I., Horowitz, A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 347–393. [Google Scholar]
- Ferreira, C.; Fernanda, B.S.; Rodriquez, A.R.S.; Siqueira, H.A.A.; Gondim, M.G.C. Resistance in field populations of Tetranychus urticae to acaricides and characterization of the inheritance of abamectin. Crop Protect. 2015, 67, 77–83. [Google Scholar] [CrossRef]
- Campos, F.; Dybas, R.A.; Krupa, D.A. Susceptibility of twospotted spider mite (Acari: Tetranychidae) populations in California to abamectin. J. Econ. Entomol. 1995, 88, 225–231. [Google Scholar] [CrossRef]
- Zalom, F.G. Strawberry Insect and Mite Control; California Strawberry Commission: Watsonville, CA, USA, 2014; p. 81. Available online: https://www.calstrawberry.com/en-us/Farm-Research/Annual-Research-Reports (accessed on 1 April 2020).
- Stumpf, N.; Nauen, R. Cross-resistance, inheritance, and biochemistry of mitochondrial electron transport inhibitor-acaricide resistance in Tetranychus urticae (Acari: Tetranychidae). J. Econ. Entomol. 2001, 94, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Silva, E.; Raga, A.; Souza, F.M.F. Abamectin resistance in Tetranychus urticae Koch (Acari: Tetranychidae): Selection, cross resistance, and stability of resistance. Neotrop. Entomol. 2005, 34, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Kerns, D.; Gore, J.; Lorenz, G.; Stewert, S. Susceptibility of twospotted spider mites (Tetranychus urticae) to abamecintin in Midsouth cotton. Crop Protect. 2017, 98, 179–183. [Google Scholar] [CrossRef]
- Dennehy, T.J.; Nyrop, J.P.; Martinson, T.E. Characterization and exploitation of instability of spider mite resistance to acaricides in managing resistance to agrochemicals. In Managing Resistance to Agrochemicals; Symposium Series, 421; Green, B.M., Homer, M., Moberg, W., Eds.; American Chemical Society: Washington, DC, USA, 1990. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Spider Mites ± SEM Per Leaf | ||||
|---|---|---|---|---|
| Date | Untreated Control | Abamectin | Cyflumetofen | Etoxazole |
| 7/25 | 0.79 ± 0.27 a | 0.06 ± 0.04 ab | 0.02 ± 0.01 b | 0.39 ± 0.13 ab |
| 8/1 | 5.64 ± 1.40 a | 0.70 ± 0.17 b | 0.73 ± 0.35 b | 0.48 ± 0.16 b |
| 8/8 | 9.68 ± 2.44 a | 1.13 ± 0.29 b | 0.85 ± 0.28 b | 0.07 ± 0.03 b |
| 8/15 1 | ||||
| 8/22 | 6.16 ± 0.86 a | 3.80 ± 0.67 ab | 2.15 ± 0.35 b | 4.95 ± 1.28 ab |
| Mean Spider Mites ± SEM Per Leaf | ||||
|---|---|---|---|---|
| Date | Untreated Control | Abamectin | Cyflumetofen | Etoxazole |
| 7/24 | 0.67 ± 0.17 a | 0.75 ± 0.37 a | 0.38 ± 0.13 a | 0.03 ± 0.02 a |
| 7/31 | 1.18 ± 0.31 a | 0.58 ± 0.18 a | 1.12 ± 0.41 a | 1.40 ± 0.36 a |
| 8/7 | 7.27 ± 1.72 a | 2.38 ± 0.65 a | 5.87 ± 2.10 a | 5.00 ± 0.98 a |
| 8/14 | 5.31 ± 0.93 a | 3.75 ± 0.64 a | 6.94 ± 1.11 a | 3.86 ± 0.87 a |
| 8/21 | 4.27 ± 0.69 a | 1.05 ± 0.48 b | 1.55 ± 0.52 b | 0.32 ± 0.10 b |
| Population | N 1 | Slope log10(ppm) (SE) | Χ2 (p Value) | LC50 (95% CL), ppm | Resistance Ratio 2 |
|---|---|---|---|---|---|
| SUS1 | 455 | 1.33 (0.17) | 59.80 (<0.0001) | 0.39 (0.27–0.52) | - |
| FRSCO1 | 945 | 1.50 (0.16) | 84.10 (<0.0001) | 1.16 (0.98–1.14) | 3 (0.95–9.43) |
| FRSCO2 | 189 | 2.10 (0.1498) | |||
| FRSCO3 | 467 | 0.72 (0.14) | 27.90 (<0.0001) | 6.24 (3.63–12.77) | 16 (3.88–65.91) |
| FRSCO4 | 617 | 1.07 (0.18) | 33.80 (<0.0001) | 7.23 (4.7–14.26) | 18.5 (3.69–92.84) |
| FRSCO5 | 791 | 0.81 (0.12) | 42.20 (<0.0001) | 2.28 (1.51–3.55) | 5.8 (1.49–22.52) |
| KERCO1 | 381 | 0.59 (0.4415) | |||
| KERCO2 | 477 | 1.68 (0.1948) | |||
| KERCO3 | 250 | 0.24 (0.2444) | |||
| TULCO1 | 376 | 0.53 (0.21) | 6.71 (<0.0096) | 5.11 (1.83–2375) | 13.1 (2.37–72.54) |
| Code | Location |
|---|---|
| SUS1 | University of California, Kearney Ag Station |
| FRSCO1 | Navelencia, Fresno Co |
| FRSCO2 | Fresno, Fresno Co |
| FRSCO3 | Raisin City, Fresno Co |
| FRSCO4 | University of California, Westside Field Station |
| FRSCO5 | West Manning Ave |
| KERCO1 | McFarland, Kern Co |
| KERCO2 | Schofield, Kern Co (experimental site 1) |
| KEROC3 | Whistler, Kern Co (experimental site 2) |
| TULCO1 | Corcoran, Tulare Co |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tollerup, K.; Higbee, B. Evaluation of a ‘Preventative’ Strategy to Manage Spider Mites on Almond. Insects 2020, 11, 772. https://doi.org/10.3390/insects11110772
Tollerup K, Higbee B. Evaluation of a ‘Preventative’ Strategy to Manage Spider Mites on Almond. Insects. 2020; 11(11):772. https://doi.org/10.3390/insects11110772
Chicago/Turabian StyleTollerup, Kristen, and Bradley Higbee. 2020. "Evaluation of a ‘Preventative’ Strategy to Manage Spider Mites on Almond" Insects 11, no. 11: 772. https://doi.org/10.3390/insects11110772