Comparative Transcriptome Analysis of Two Root-Feeding Grape Phylloxera (D. vitifoliae) Lineages Feeding on a Rootstock and V. vinifera

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect and Plant Material
2.2. Insect Samples for RNA Extraction
2.3. RNA Extraction and RNA Sequencing Analysis
2.4. Effector Candidate Characterization and BlastP Analysis
3. Results
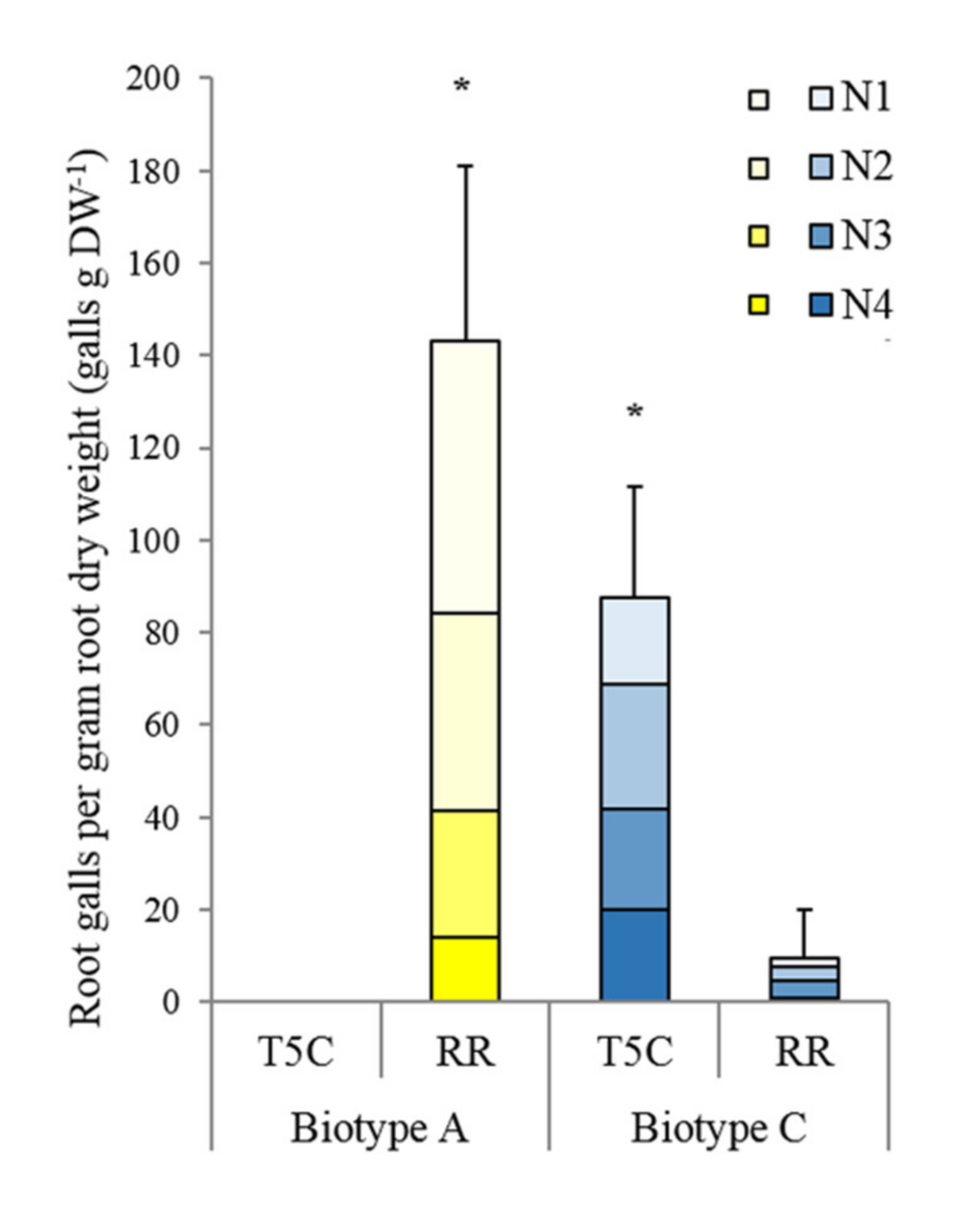
3.1. Biotype Confirmation
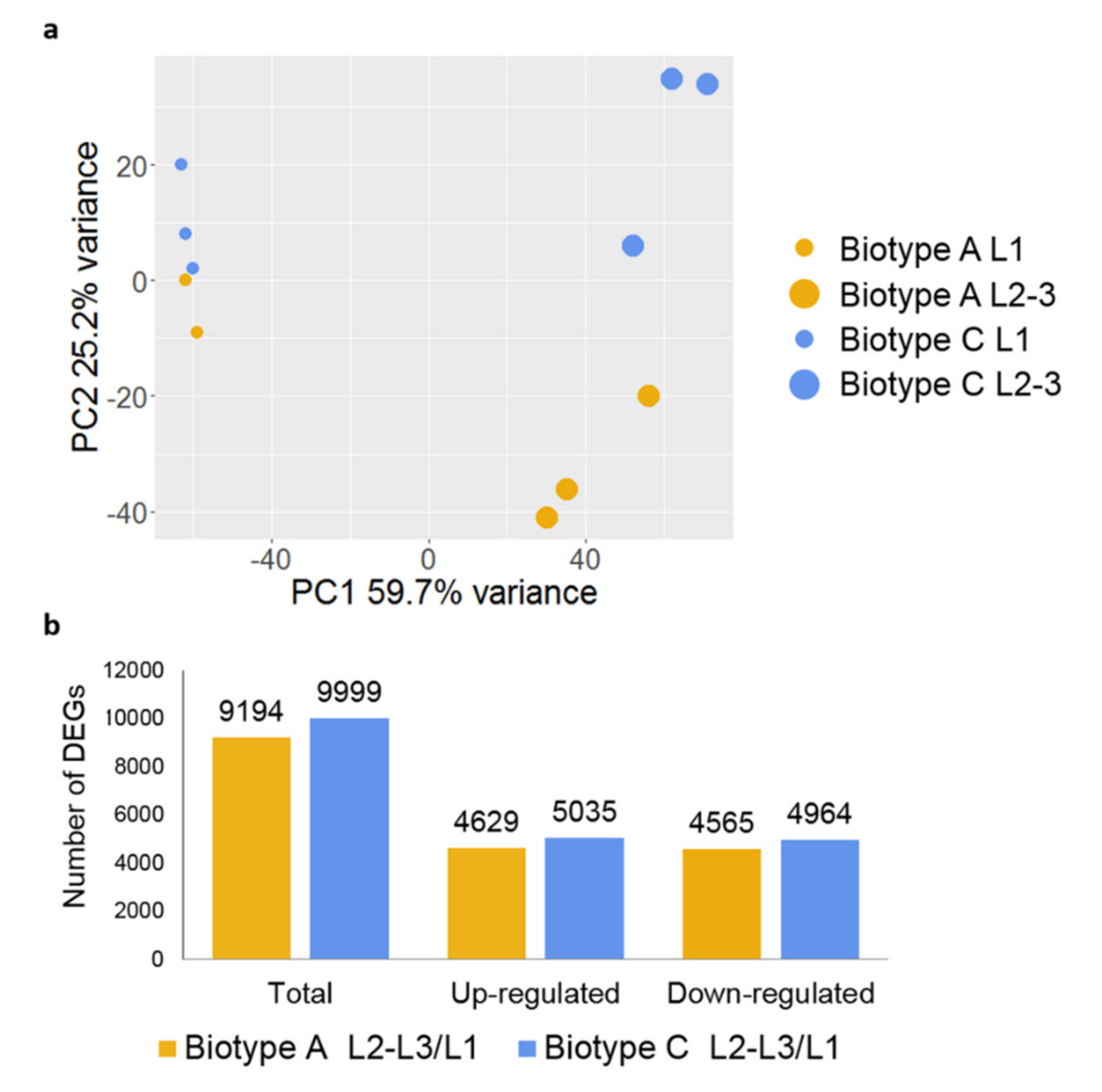
3.2. Differentially Expressed Genes between Probing and Feeding Larvae
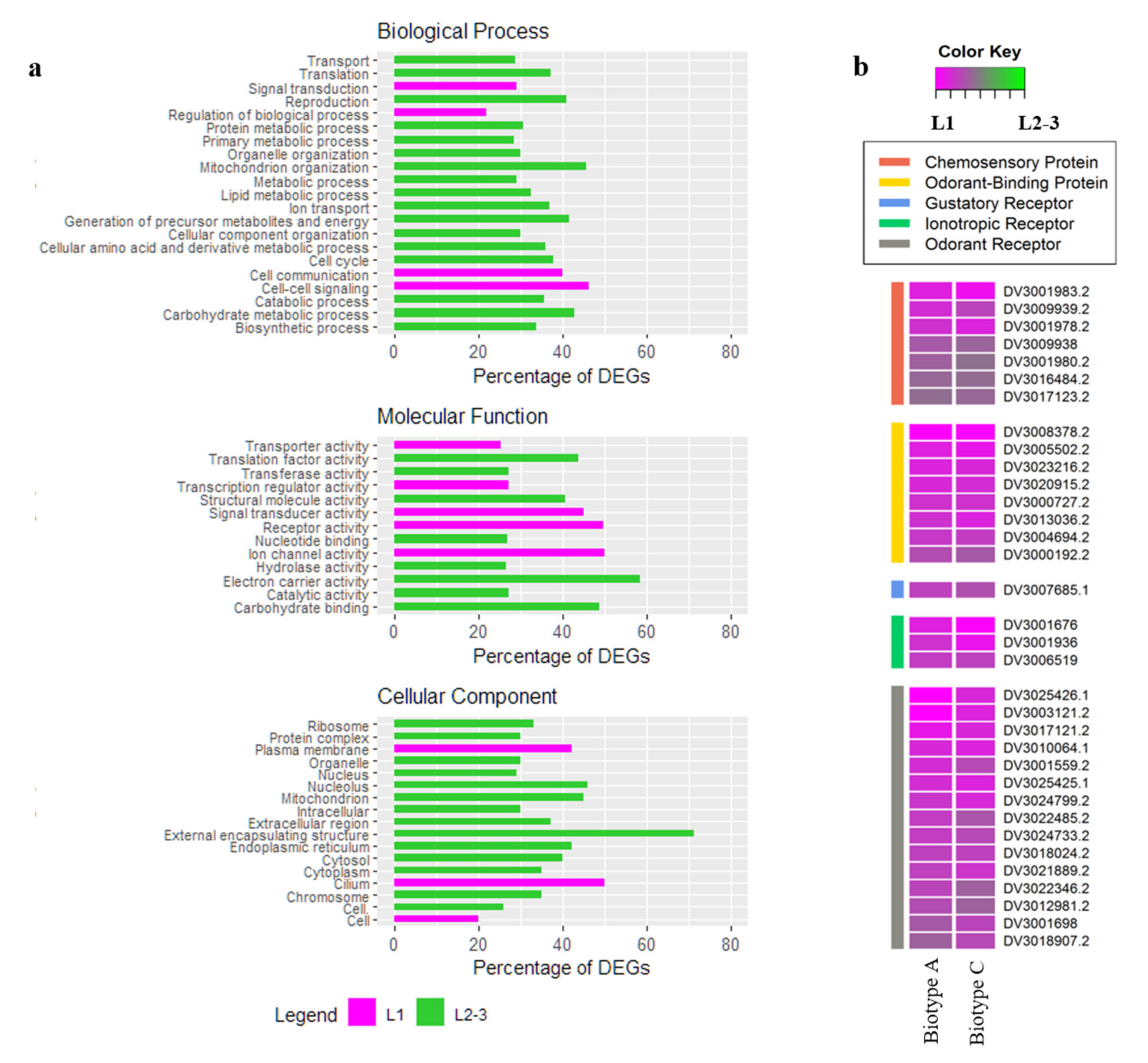
3.3. DEGs in L1 Larvae Probing on Riesling vs. Teleki 5C
3.4. DEGs in L2-3 Larvae Feeding on Riesling vs. Teleki 5C
4. Discussion
4.1. Specific Genes Involved in Larval Chemoreception
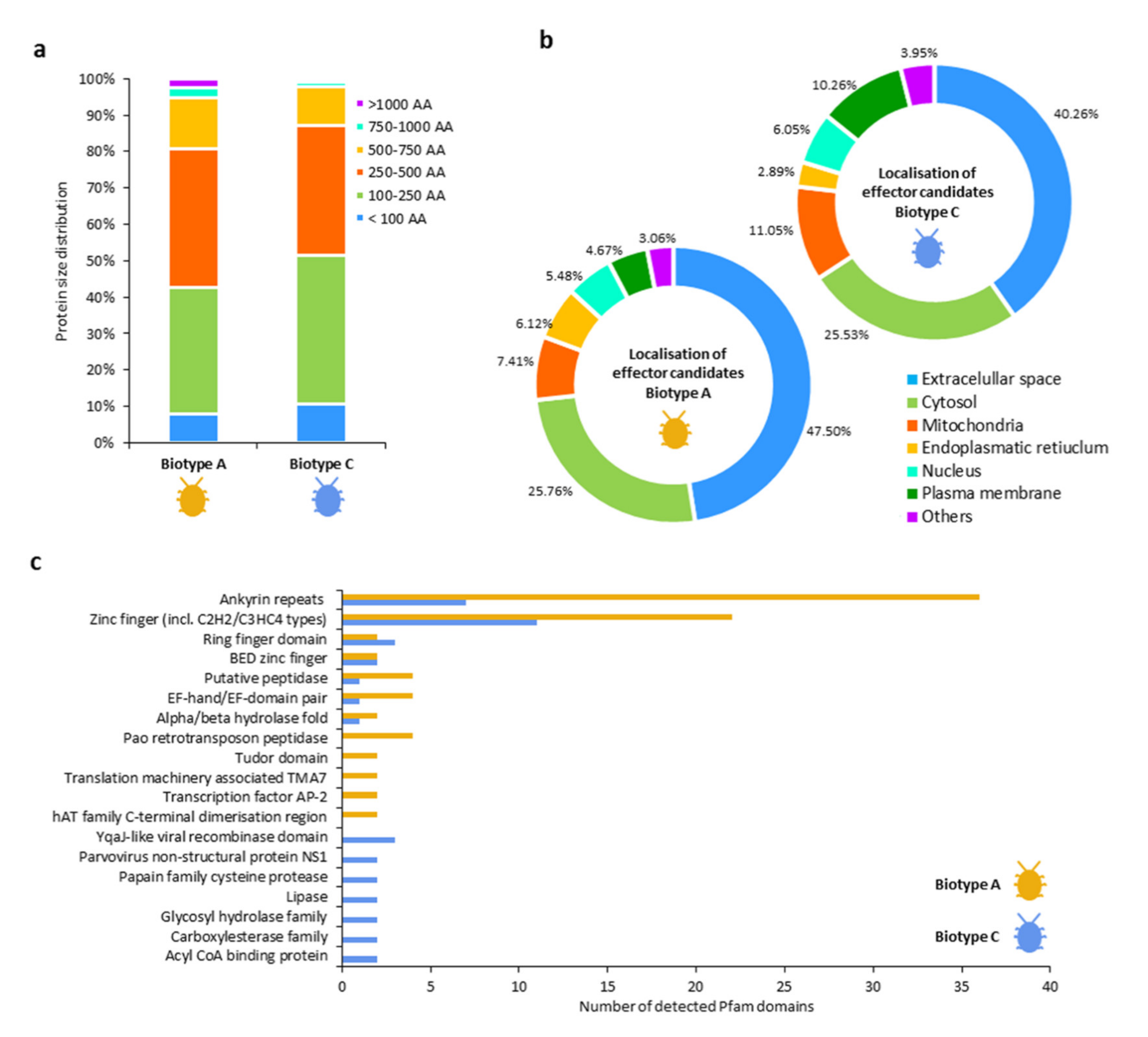
4.2. DEGs Associated with Putative Effector Candidates Involved with Host Plant Physiology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effector Candidate | Putative Function | Grape Phylloxera | Host Tissue | Reference | ||
|---|---|---|---|---|---|---|
| Genetic Structure | Life Stage | Leaf Gall | Root Gall | |||
| Calreticulins, calmodulins | Interference with Ca+ signaling, regulation of cell division/proliferation traits | Pop, sfl | L2/3/4 & A | Frontenac, Harmony, V.arizonica, V.riparia | Cab. Sauvignon, T5C | [42,43] * |
| Carboxypeptidases, serine proteases | Degradation of host defensive proteins, facilitation of amino acid uptake | Sfl | L2/3 | Harmony | Cab. Sauvignon, T5C | [40,42] * |
| Esterases, mannosidases | Cell wall degradation and loosening, host defense detoxification | Sfl | L2/3 | - | T5C | [42] * |
| Glucose dehydrogenases | Interference of host defense signaling (SA/JA/ET), detoxification, sugar intake | Sfl | L2/3 | - | T5C | [42] * |
| Heat shock proteins | Co-regulation of abiotic stress pathways | Pop, sfl | L2/3/4 & A | Vitis spp. | T5C | [42,78] |
| Peroxidase, peroxidoredoxins | H2O2/ROS detoxification | Sfl | L2/3 | - | T5C | [42] * |
| Protein disulfide isomerases, mucins | Stylet sheath formation | Sfl | L2/3 | - | T5C | [42] |
| Proteins with RING-type zinc finger, EF-hand, ankyrin repeat domains | Modulation of cellular growth/development processes, ETI signaling | Pop, sfl | L2/3/4 & A | Frontenac, Harmony, V.arizonica, V.riparia | Cab. Sauvignon, T5C | [41,43,44] * |
| RING-containing E3 ligases | Interaction with cellulose biosynthesis (VviCSLD5), protein translation (VviRPS4B) | Pop | L2/3/4 & A | V. riparia | − | [41] |
4.3. DEGs Associated with Gall Formation Traits
4.4. DEGs Associated with Host Defense Response and Insect Digestive Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Basky, Z. Biotypic and pest status differences between Hungarian and South African populations of Russian wheat aphid, Diuraphis noxia (Kurdjumov) (Homoptera: Aphididae). Pest Manag. Sci. 2003, 59, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Jones, C.M.; Devine, G.; Zhang, F.; Denholm, I.; Gorman, K. Insecticide resistance in Bemisia tabaci biotype Q (Hemiptera: Aleyrodidae) from China. Crop Prot. 2010, 29, 429–434. [Google Scholar] [CrossRef]
- Pawlowski, M.; Hill, C.B.; Voegtlin, D.J.; Hartman, G.L. Soybean aphid intrabiotype variability based on colonization of specific soybean genotypes. Insect Sci. 2015, 22, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Forneck, A.; Huber, L. (A)sexual reproduction—A review of life cycles of grape phylloxera, Daktulosphaira vitifoliae. Entomol. Exp. Appl. 2009, 131, 1–10. [Google Scholar] [CrossRef]
- Delmotte, F.; Papura, D.; Rispe, C.; Legeai, F.; Jaquiéry, J.; Breteaudeau, A.; Tagu, D.; Powell, K.S.; Forneck, A. The grape phylloxera genome sequencing project. Acta Hortic. 2014, 15–19. [Google Scholar] [CrossRef]
- Favret, C.; Blackman, R.; Miller, G.; Victor, B. Catalog of the phylloxerids of the world (Hemiptera: Phylloxeridae). ZooKeys 2016, 629, 83–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, R.; Bragard, C.; Candresse, T.; Gilioli, G.; Grégoire, J.; Holb, I.; Jeger, M.J.; Karadjova, O.E.; Magnusson, C.; Makowski, D.; et al. Scientific opinion on the risk to plant health posed by Daktulosphaira vitifoliae (Fitch) in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2014, 12, 3678. [Google Scholar] [CrossRef]
- Forneck, A.; Kleinmann, S.; Blaich, R.; Anvari, S.F. Histochemistry and anatomy of phylloxera (Daktulosphaira vitifoliae) nodosities on young roots of grapevine (Vitis spp). Vitis 2002, 41, 93–98. [Google Scholar]
- Kellow, A.V.; Sedgley, M.; Van Heeswijck, R. Interaction between Vitis vinifera and grape phylloxera: Changes in root tissue during nodosity formation. Ann. Bot. 2004, 93, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Nabity, P.D.; Haus, M.J.; Berenbaum, M.R.; DeLucia, E.H. Leaf-galling phylloxera on grapes reprograms host metabolism and morphology. Proc. Natl. Acad. Sci. USA 2013, 110, 16663–16668. [Google Scholar] [CrossRef] [Green Version]
- Ryan, F.J.; Omer, A.D.; Aung, L.H.; Granett, J. Effects of infestation by grape phylloxera on sugars, free amino acids, and starch of grapevine roots. Vitis 2000, 39, 175–176. [Google Scholar]
- Porten, M.; Huber, L. An assessment method for the quantification of Daktulosphaira vitifoliae (Fitch) (Hem., Phylloxeridae) populations in the field. J. Appl. Entomol. 2003, 127, 157–162. [Google Scholar] [CrossRef]
- Griesser, M.; Lawo, N.C.; Crespo-Martinez, S.; Schoedl-Hummel, K.; Wieczorek, K.; Gorecka, M.; Liebner, F.; Zweckmair, T.; Stralis Pavese, N.; Kreil, D.; et al. Phylloxera (Daktulosphaira vitifoliae Fitch) alters the carbohydrate metabolism in root galls to allowing the compatible interaction with grapevine (Vitis ssp.) roots. Plant Sci. 2015, 234, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, L.; Wolf, T.; Eimert, K.; Schröder, M.-B. Differential gene expression during hypersensitive response in Phylloxera-resistant rootstock ‘Börner’ using custom oligonucleotide arrays. J. Plant Interact. 2009, 4, 261–269. [Google Scholar] [CrossRef]
- Lawo, N.C.; Weingart, G.J.F.; Schuhmacher, R.; Forneck, A. The volatile metabolome of grapevine roots: First insights into the metabolic response upon phylloxera attack. Plant Physiol. Biochem. 2011, 49, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Eitle, M.W.; Loacker, J.; Meng-Reiterer, J.; Schuhmacher, R.; Griesser, M.; Forneck, A. Polyphenolic profiling of roots (Vitis spp.) under grape phylloxera (D. vitifoliae Fitch) attack. Plant Physiol. Biochem. 2019, 135, 174–181. [Google Scholar] [CrossRef]
- Eitle, M.W.; Griesser, M.; Vankova, R.; Dobrev, P.; Aberer, S.; Forneck, A. Grape phylloxera (D. vitifoliae) manipulates SA/JA concentrations and signalling pathways in root galls of Vitis spp. Plant Physiol. Biochem. 2019, 144, 85–91. [Google Scholar] [CrossRef]
- Wang, F.-P.; Zhao, P.-P.; Zhang, L.; Zhai, H.; Du, Y.-P. Functional characterization of WRKY46 in grape and its putative role in the interaction between grape and phylloxera (Daktulosphaira vitifoliae). Hortic. Res. 2019, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Huber, L. Schaderrerger im Wurzelraum von Reben (Vitis spp.)—Vorkommen, Wirkung, Interaktionen—und Möglichkeiten zu deren Kontrolle durch Maßnahmen des Integrated Pest Managements (IPM). Ph.D. Thesis, Johannes Gutenberg Universität Mainz, Fachbereich Biologie, Mainz, Germany, 2007. [Google Scholar]
- Powell, K.S.; Cooper, P.D.; Forneck, A. Chapter four—The biology, physiology and host–plant interactions of grape phylloxera Daktulosphaira vitifoliae. In Advances in Insect Physiology; Behaviour and Physiology of Root Herbivores; Academic Press: Cambridge, MA, USA, 2013; Volume 45, pp. 159–218. [Google Scholar]
- Savi, T.; García González, A.; Herrera, J.C.; Forneck, A. Gas exchange, biomass and non-structural carbohydrates dynamics in vines under combined drought and biotic stress. BMC Plant Biol. 2019, 19, 408. [Google Scholar] [CrossRef]
- Kingston, K.B.; Powell, K.S.; Cooper, P.D. Characterising the root-feeding habits of grape phylloxera using electrical penetration graph. Acta Hortic. 2007, 33–46. [Google Scholar] [CrossRef]
- Raman, A.; Beiderbeck, R.; Herth, W. Early subcellular responses of susceptible and resistant Vitis taxa to feeding by grape phylloxera Daktulosphaira vitifoliae. Bot. Helv. 2009, 119, 31–39. [Google Scholar] [CrossRef]
- Hofmann, E.L. Die Histologie der Nodositäten verschiedener Rebensorten bei Reblausbefall. Vitis 1957, 1, 125–141. [Google Scholar] [CrossRef]
- Riaz, S.; Pap, D.; Uretsky, J.; Laucou, V.; Boursiquot, J.-M.; Kocsis, L.; Andrew Walker, M. Genetic diversity and parentage analysis of grape rootstocks. Theor. Appl. Genet. 2019, 132, 1847–1860. [Google Scholar] [CrossRef] [PubMed]
- Granett, J.; Walker, M.A.; Kocsis, L.; Omer, A.D. Biology and management of grape phylloxera. Annu. Rev. Entomol. 2001, 46, 387–412. [Google Scholar] [CrossRef] [PubMed]
- Arancibia, C.; Riaz, S.; Agüero, C.; Ramirez-Corona, B.; Alonso, R.; Buscema, F.; Martínez, L.; Walker, M.A. Grape phylloxera (Daktulosphaira vitifoliae Fitch) in Argentina: Ecological associations to diversity, population structure and reproductive mode. Aust. J. Grape Wine Res. 2018, 24, 284–291. [Google Scholar] [CrossRef]
- Forneck, A.; Mammerler, R.; Tello, J.; Breuer, M.; Müller, J.; Fahrentrapp, J. First European leaf-feeding grape phylloxera (Daktulosphaira vitifoliae Fitch) survey in Swiss and German commercial vineyards. Eur. J. Plant Pathol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, L.; Granett, J.; Walker, M.A.; Lin, H.; Omer, A.D. Grape phylloxera populations adapted to Vitis berlandieri x V. riparia rootstocks. Am. J. Enol. Vitic. 1999, 50, 101–106. [Google Scholar]
- Granett, J.; Timper, P.; Lider, L.A. Grape phylloxera (Daktulosphaira vitifoliae) (Homoptera: Phylloxeridae) biotypes in California. J. Econ. Entomol. 1985, 78, 1463–1467. [Google Scholar] [CrossRef]
- Forneck, A.; Powell, K.S.; Walker, M.A. Scientific opinion: Improving the definition of grape phylloxera biotypes and standardizing biotype screening protocols. Am. J. Enol. Vitic. 2016, 67, 371–376. [Google Scholar] [CrossRef] [Green Version]
- King, P.D.; Rilling, G. Variations in the galling reaction of grapevines: Evidence of different phylloxera biotypes and clonal reaction to phylloxera. Vitis 1985, 24, 32–42. [Google Scholar]
- Eitle, M.W.; Forneck, A. Comparison of bioassays to biotype grape phylloxera (Daktulosphaira vitifoliae Fitch) on Vitis ssp. Vitis 2017, 56, 141–146. [Google Scholar]
- Kocsis, L.; Granett, J.; Walker, M.A. Performance of Hungarian phylloxera strains on Vitis riparia rootstocks. J. Appl. Entomol. 2002, 126, 567–571. [Google Scholar] [CrossRef]
- Eitle, M.W.; Griesser, M.; Dobrev, F.; Vankova, R.; Forneck, A. First insights on phytohormones during the compatible grapevine-phylloxera interaction. Acta Hortic. 2016, 1188, 255–264. [Google Scholar] [CrossRef]
- Du, Y.-P.; Wang, Z.-S.; Yang, Y.; Zhao, Q.; Zhai, H.; Wang, Z.-Y. Nodosity formation and nutrition consumption in grape cultivars with different phylloxera resistance and infested by grape phylloxera. Acta Entomol. Sin. 2008, 51, 1050–1054. [Google Scholar]
- Lawo, N.C.; Griesser, M.; Forneck, A. Expression of putative expansin genes in phylloxera (Daktulosphaira vitifoliae Fitch) induced root galls of Vitis spp. Eur. J. Plant Pathol. 2013, 136, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tello, J.; Forneck, A. Use of DNA markers for grape phylloxera population and evolutionary genetics: From RAPDs to SSRs and beyond. Insects 2019, 10, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forneck, A.; Anhalt, U.C.M.; Mammerler, R.; Griesser, M. No evidence of superclones in leaf-feeding forms of Austrian grape phylloxera (Daktulosphaira vitifoliae). Eur. J. Plant Pathol. 2015, 142, 441–448. [Google Scholar] [CrossRef]
- Rispe, C.; Legeai, F.; Papura, D.; Bretaudeau, A.; Hudaverdian, S.; Le Trionnaire, G.; Tagu, D.; Jaquiéry, J.; Delmotte, F. De novo transcriptome assembly of the grapevine phylloxera allows identification of genes differentially expressed between leaf- and root-feeding forms. BMC Genom. 2016, 17, 219. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Rispe, C.; Nabity, P.D. Secretory RING finger proteins function as effectors in a grapevine galling insect. BMC Genom. 2019, 20, 923. [Google Scholar] [CrossRef]
- Eitle, M.W.; Carolan, J.C.; Griesser, M.; Forneck, A. The salivary gland proteome of root-galling grape phylloxera (Daktulosphaira vitifoliae Fitch) feeding on Vitis spp. PLoS ONE 2019, 14, e0225881. [Google Scholar] [CrossRef]
- Taylor, T. Identification and Characterization of Candidate Calcium-Binding Effectors in Grape Phylloxera. UC Riverside: University Honors. 2019. Available online: https://escholarship.org/uc/item/8mk8m260 (accessed on 1 September 2020).
- Rispe, C.; Legeai, F.; Nabity, P.D.; Fernández, R.; Arora, A.K.; Baa-Puyoulet, P.; Banfill, C.R.; Bao, L.; Barberà, M.; Bouallègue, M.; et al. The genome sequence of the grape phylloxera provides insights into the evolution, adaptation, and invasion routes of an iconic pest. BMC Biol. 2020, 18, 90. [Google Scholar] [CrossRef]
- Downie, D.A. Phylogeography in a galling insect, grape phylloxera, Daktulosphaira vitifoliae (Phylloxeridae) in the fragmented habitat of the Southwest USA. J. Biogeogr. 2004, 31, 1759–1768. [Google Scholar] [CrossRef]
- Tello, J.; Mammerler, R.; Čajić, M.; Forneck, A. Major outbreaks in the nineteenth century shaped grape phylloxera contemporary genetic structure in Europe. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Y.; Fan, D.; Feng, J. Identification and expression profiling of odorant-binding proteins and chemosensory proteins of Daktulosphaira vitifoliae (Hemiptera: Phylloxeridae). J. Econ. Entomol. 2017, 110, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.C.; Edger, P.P.; Body, M.J.A.; Appel, H.M. A galling insect activates plant reproductive programs during gall development. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- De Benedictis, J.A.; Granett, J. Variability of responses of grape phylloxera (Homoptera: Phylloxeridae) to bioassays that discriminate between California biotypes. J. Econ. Entomol. 1992, 85, 1527–1534. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierleoni, A.; Martelli, P.L.; Casadio, R. PredGPI: A GPI-anchor predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. 2004, 17, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Carolan, J.C.; Caragea, D.; Reardon, K.T.; Mutti, N.S.; Dittmer, N.; Pappan, K.; Cui, F.; Castaneto, M.; Poulain, J.; Dossat, C.; et al. Predicted effector molecules in the salivary secretome of the pea aphid (Acyrthosiphon pisum): A dual transcriptomic/proteomic approach. J. Proteome Res. 2011, 10, 1505–1518. [Google Scholar] [CrossRef] [PubMed]
- Boulain, H.; Legeai, F.; Guy, E.; Morlière, S.; Douglas, N.E.; Oh, J.; Murugan, M.; Smith, M.; Jaquiéry, J.; Peccoud, J.; et al. Fast evolution and lineage-specific gene family expansions of aphid salivary effectors driven by interactions with host-plants. Genome Biol. Evol. 2018, 10, 1554–1572. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, P.; Cock, P.J.A.; Bos, J. Comparative transcriptomics and proteomics of three different aphid species identifies core and diverse effector sets. BMC Genom. 2016, 17, 172. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, J.; Sun, J.; Francis, F.; Chen, J. Transcriptome analysis of the salivary glands of the grain aphid, Sitobion avenae. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. MPMI 2009, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell. Mol. Life Sci. CMLS 2018, 75, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Montagné, N.; de Fouchier, A.; Newcomb, R.D.; Jacquin-Joly, E. Advances in the identification and characterization of olfactory receptors in insects. Prog. Mol. Biol. Transl. Sci. 2015, 130, 55–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, W.; Zhang, Z.; Zhang, Y.; Guo, Y. Functional characteristics of a novel chemosensory protein in the cotton bollworm Helicoverpa armigera (Hübner). J. Integr. Agric. 2013, 12, 853–861. [Google Scholar] [CrossRef]
- Ali, S.; Ahmed, M.Z.; Li, N.; Ali, S.A.I.; Wang, M.-Q. Functional characteristics of chemosensory proteins in the sawyer beetle Monochamus alternatus Hope. Bull. Entomol. Res. 2019, 109, 34–42. [Google Scholar] [CrossRef]
- Ji, R.; Yu, H.; Fu, Q.; Chen, H.; Ye, W.; Li, S.; Lou, Y. Comparative transcriptome analysis of salivary glands of two populations of rice brown planthopper, Nilaparvata lugens, that differ in virulence. PLoS ONE 2013, 8, e79612. [Google Scholar] [CrossRef]
- Boulain, H.; Legeai, F.; Jaquiéry, J.; Guy, E.; Morlière, S.; Simon, J.-C.; Sugio, A. Differential expression of candidate salivary effector genes in pea aphid biotypes with distinct host plant specificity. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Nicholson, S.J.; Hartson, S.D.; Puterka, G.J. Proteomic analysis of secreted saliva from Russian wheat aphid (Diuraphis noxia Kurd.) biotypes that differ in virulence to wheat. J. Proteom. 2012, 75, 2252–2268. [Google Scholar] [CrossRef]
- Nicholson, S.J.; Puterka, G.J. Variation in the salivary proteomes of differentially virulent greenbug (Schizaphis graminum Rondani) biotypes. J. Proteom. 2014, 105, 186–203. [Google Scholar] [CrossRef]
- Al-Khodor, S.; Price, C.T.; Kalia, A.; Abu Kwaik, Y. Functional diversity of ankyrin repeats in microbial proteins. Trends Microbiol. 2010, 18, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Nam, J.; Boyes, D.C.; Holt, B.F.; Hubert, D.A.; Wiig, A.; Dangl, J.L. A duplicated pair of Arabidopsis RING-finger E3 ligases contribute to the RPM1- and RPS2-mediated hypersensitive response. Plant J. Cell Mol. Biol. 2005, 44, 258–270. [Google Scholar] [CrossRef]
- Abramovitch, R.B.; Janjusevic, R.; Stebbins, C.E.; Martin, G.B. Type III effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 2851–2856. [Google Scholar] [CrossRef] [Green Version]
- Nabity, P.D. Insect-induced plant phenotypes: Revealing mechanisms through comparative genomics of galling insects and their hosts. Am. J. Bot. 2016, 103, 979–981. [Google Scholar] [CrossRef] [Green Version]
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host-plant. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef]
- Will, T. Function of aphid saliva in aphid-plant interaction. Biol. Ecol. Aphids 2016, 109, 221–237. [Google Scholar]
- Knoblauch, M.; Froelich, D.R.; Pickard, W.F.; Peters, W.S. SEORious business: Structural proteins in sieve tubes and their involvement in sieve element occlusion. J. Exp. Bot. 2014, 65, 1879–1893. [Google Scholar] [CrossRef]
- Guiguet, A.; Dubreuil, G.; Harris, M.O.; Appel, H.M.; Schultz, J.C.; Pereira, M.H.; Giron, D. Shared weapons of blood- and plant-feeding insects: Surprising commonalities for manipulating hosts. J. Insect Physiol. 2016, 84, 4–21. [Google Scholar] [CrossRef]
- Hassan, S.; Behm, C.A.; Mathesius, U. Effectors of plant parasitic nematodes that re-program root cell development. Funct. Plant Biol. 2010, 37, 933–942. [Google Scholar] [CrossRef]
- Jaouannet, M.; Magliano, M.; Arguel, M.J.; Gourgues, M.; Evangelisti, E.; Abad, P.; Rosso, M.N. The root-knot nematode calreticulin Mi-CRT is a key effector in plant defense suppression. Mol. Plant Microbe Interact. 2012, 26, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhang, F.; He, Q.; He, G. Molecular dynamics of detoxification and toxin-tolerance genes in brown planthopper (Nilaparvata lugens Stål., Homoptera: Delphacidae) feeding on resistant rice plants. Arch. Insect Biochem. Physiol. 2005, 59, 59–66. [Google Scholar] [CrossRef]
- Urbanska, A.; Tjallingii, W.F.; Dixon, A.F.G.; Leszczynski, B. Phenol oxidising enzymes in the grain aphid’s saliva. Entomol. Exp. Appl. 1998, 86, 197–203. [Google Scholar] [CrossRef]
- Giordanengo, P.; Brunissen, L.; Rusterucci, C.; Vincent, C.; van Bel, A.; Dinant, S.; Girousse, C.; Faucher, M.; Bonnemain, J.-L. Compatible plant-aphid interactions: How aphids manipulate plant responses. C. R. Biol. 2010, 333, 516–523. [Google Scholar] [CrossRef]
- Burger, B.V.; de Klerk, C.A.; Morr, M.; Burger, W.J.G. Identification, synthesis, and field tests of the sex pheromone of Margarodes prieskaensis (Jakubski). J. Chem. Ecol. 2017, 43, 94–105. [Google Scholar] [CrossRef]
- Du, Y.-P.; Jiang, E.-S.; Wang, F.-P.; Zhang, S.-Z.; Zhai, H. Gene expression profiling of rootstock ‘140Ru’ and Vitis vinifera L. cv. ‘Crimson Seedless’ grape roots infected with grape phylloxera. Plant Growth Regul. 2014, 73, 1–8. [Google Scholar] [CrossRef]
- Macedo, M.R.; Das Graças Machado Freire, M. Insect digestive enzymes as a target for pest control. Invertebr. Surviv. J. 2011, 8, 190–198. [Google Scholar]
- Jongsma, M.A.; Beekwilder, J. Co-evolution of insect proteases and plant protease inhibitors. Curr. Protein Pept. Sci. 2011, 12, 437–447. [Google Scholar] [CrossRef]
- Hartl, M.; Giri, A.P.; Kaur, H.; Baldwin, I.T. The multiple functions of plant serine protease inhibitors: Defense against herbivores and beyond. Plant Signal. Behav. 2011, 6, 1009–1011. [Google Scholar] [CrossRef] [Green Version]
- Bayés, A.; de la Vega, M.R.; Vendrell, J.; Aviles, F.X.; Jongsma, M.A.; Beekwilder, J. Response of the digestive system of Helicoverpa zea to ingestion of potato carboxypeptidase inhibitor and characterization of an uninhibited carboxypeptidase B. Insect Biochem. Mol. Biol. 2006, 36, 654–664. [Google Scholar] [CrossRef]
- Srinivasan, A.; Giri, A.P.; Gupta, V.S. Structural and functional diversities in lepidopteran serine proteases. Cell. Mol. Biol. Lett. 2006, 11, 132–154. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, A.; Sharma, R.; Gakhar, S.K. Identification, characterization and analysis of expression of gene encoding carboxypeptidase A in Anopheles culicifacies A (Diptera: Culicidae). Acta Trop. 2014, 139, 123–130. [Google Scholar] [CrossRef]
- Schultz, J.C.; Appel, H.M.; Ferrieri, A.; Arnold, T.M. Flexible resource allocation during plant defense responses. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Divol, F.; Vilaine, F.; Thibivilliers, S.; Amselem, J.; Palauqui, J.-C.; Kusiak, C.; Dinant, S. Systemic response to aphid infestation by Myzus persicae in the phloem of Apium graveolens. Plant Mol. Biol. 2005, 57, 517–540. [Google Scholar] [CrossRef]
- Andreas, P.; Kisiala, A.; Emery, R.J.N.; De Clerck-Floate, R.; Tooker, J.F.; Price, P.W.; Miller, D.G., III; Chen, M.-S.; Connor, E.F. Cytokinins are abundant and widespread among insect species. Plants 2020, 9, 208. [Google Scholar] [CrossRef] [Green Version]
- De Meutter, J.; Tytgat, T.; Prinsen, E.; Gheysen, G.; Van Onckelen, H.; Gheysen, G. Production of auxin and related compounds by the plant parasitic nematodes Heterodera schachtii and Meloidogyne incognita. Commun. Agric. Appl. Biol. Sci. 2005, 70, 51–60. [Google Scholar]
- Loxdale, H.D.; Balog, A. Aphid specialism as an example of ecological-evolutionary divergence. Biol. Rev. Camb. Philos. Soc. 2018, 93, 642–657. [Google Scholar] [CrossRef]
- Vorwerk, S.; Martinez-Torres, D.; Forneck, A. Pantoea agglomerans-associated bacteria in grape phylloxera (Daktulosphaira vitifoliae, Fitch). Agric. For. Entomol. 2007, 9, 57–64. [Google Scholar] [CrossRef]




| Secretory Prediction Pipeline | Biotype A | Biotype C | ||
| Up-regulated genes | 1133 | 100.0% | 789 | 100.0% |
| Transmembrane helices | 186 | 16.4% | 176 | 22.3% |
| GPI-anchors | 8 | 0.7% | 7 | 0.9% |
| Transcripts without structural retaining signals | 941 | 83.1% | 609 | 77.2% |
| Classical secretion pathway | 447 | 39.5% | 231 | 29.3% |
| Non-classical secretion pathway | 407 | 35.9% | 279 | 35.4% |
| Secreted proteins | 621 | 54.8% | 380 | 48.2% |
| Grape Phylloxera Effector Candidate | Effector Candidate Homologs | ||||||
|---|---|---|---|---|---|---|---|
| A. pisum | M. persicae | S. avenae | |||||
| Reference ID | e-Value | Reference ID | e-Value | Reference ID | e-Value | ||
| Grape phylloxera biotype A | |||||||
| Calmodulin | DV3017805 | Mp_O_17539_c0_seq4|m.16201 | 5 × 10−100 | ||||
| Carbonic anhydrase | DV3001696 | XP_003241827.1 | 8 × 10−88 | Mp_J_18423_c0_seq1|m.16524 | 10 × 10−64 | c14864_g1 | 7 × 10−88 |
| Glucose dehydrogenase | DV3020319 | XP_001946107.1 | 6 × 10−33 | Mp_J_18221_c0_seq1|m.16042 | 0 × 10−00 | c10172_g2 | 2 × 10−129 |
| Serine-threonine phosphatase | DV3006248 | XP_008186276.1 | 3 × 10−117 | ||||
| DV3009036 | XP_008186276.1 | 3 × 10−102 | |||||
| Grape phylloxera biotype C | |||||||
| Carboxylesterase | DV3000733.2 | XP_001947304.2 | 9 × 10−173 | Mp_O_42087_c0_seq1|m.342653| | 6 × 10−172 | c15304_g1 | 0 × 10−00 |
| DV3025271.1 | XP_001947304.2 | 2 × 10−157 | Mp_O_42087_c0_seq1|m.342653| | 1 × 10−156 | c15304_g1 | 0 × 10−00 | |
| Carboxypeptidase | DV3007270 | Mp_J_34059_c2_seq1|m.133176| | 0 × 10−00 | ||||
| Glucose dehydrogenase | DV3004487 | XP_001946107.1 | 0 × 10−00 | Mp_O_32734_c0_seq7|m.94545| | 0 × 10−00 | c10172_g2 | 0 × 10−00 |
| Peroxidase | DV3022019 | XP_008186762.1 | 3 × 10−170 | Mp_F_35394_c1_seq17|m.129188| | 2× 10−166 | c14243_g1 | 1 × 10−161 |
| Serine protease | DV3020836 | XP_016663805.1 | 6 × 10−129 | Mp_J_29613_c0_seq1|m.54328| | 6 × 10−66 | c13826_g1 | 4 × 10−109 |
| DV3020837 | XP_016663805.1 | 2 × 10−110 | Mp_J_29613_c0_seq1|m.54328| | 3 × 10−62 | c13826_g1 | 1 × 10−93 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savoi, S.; Eitle, M.W.; Berger, H.; Curto, M.; Meimberg, H.; Griesser, M.; Forneck, A. Comparative Transcriptome Analysis of Two Root-Feeding Grape Phylloxera (D. vitifoliae) Lineages Feeding on a Rootstock and V. vinifera. Insects 2020, 11, 691. https://doi.org/10.3390/insects11100691
Savoi S, Eitle MW, Berger H, Curto M, Meimberg H, Griesser M, Forneck A. Comparative Transcriptome Analysis of Two Root-Feeding Grape Phylloxera (D. vitifoliae) Lineages Feeding on a Rootstock and V. vinifera. Insects. 2020; 11(10):691. https://doi.org/10.3390/insects11100691
Chicago/Turabian StyleSavoi, Stefania, Markus W. Eitle, Harald Berger, Manuel Curto, Harald Meimberg, Michaela Griesser, and Astrid Forneck. 2020. "Comparative Transcriptome Analysis of Two Root-Feeding Grape Phylloxera (D. vitifoliae) Lineages Feeding on a Rootstock and V. vinifera" Insects 11, no. 10: 691. https://doi.org/10.3390/insects11100691