Feeding Behavior of Asian Citrus Psyllid [Diaphorina citri (Hemiptera: Liviidae)] Nymphs and Adults on Common Weeds Occurring in Cultivated Citrus Described Using Electrical Penetration Graph Recordings

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Plants
2.3. Electrical Penetration Graph Recordings of D. citri Nymphs and Adults
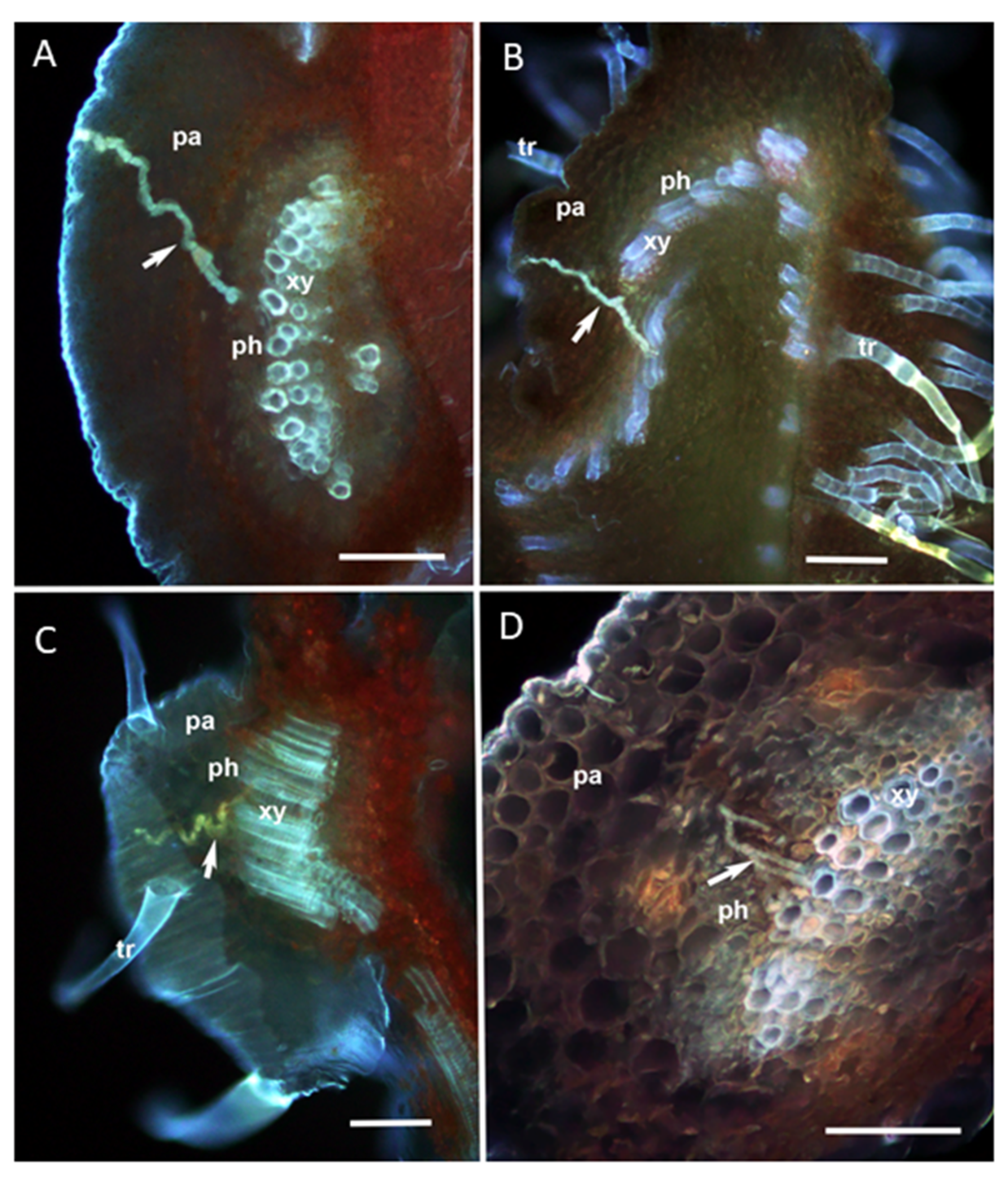
2.4. Visualization of Salivary Sheaths Produced by D. citri Adults
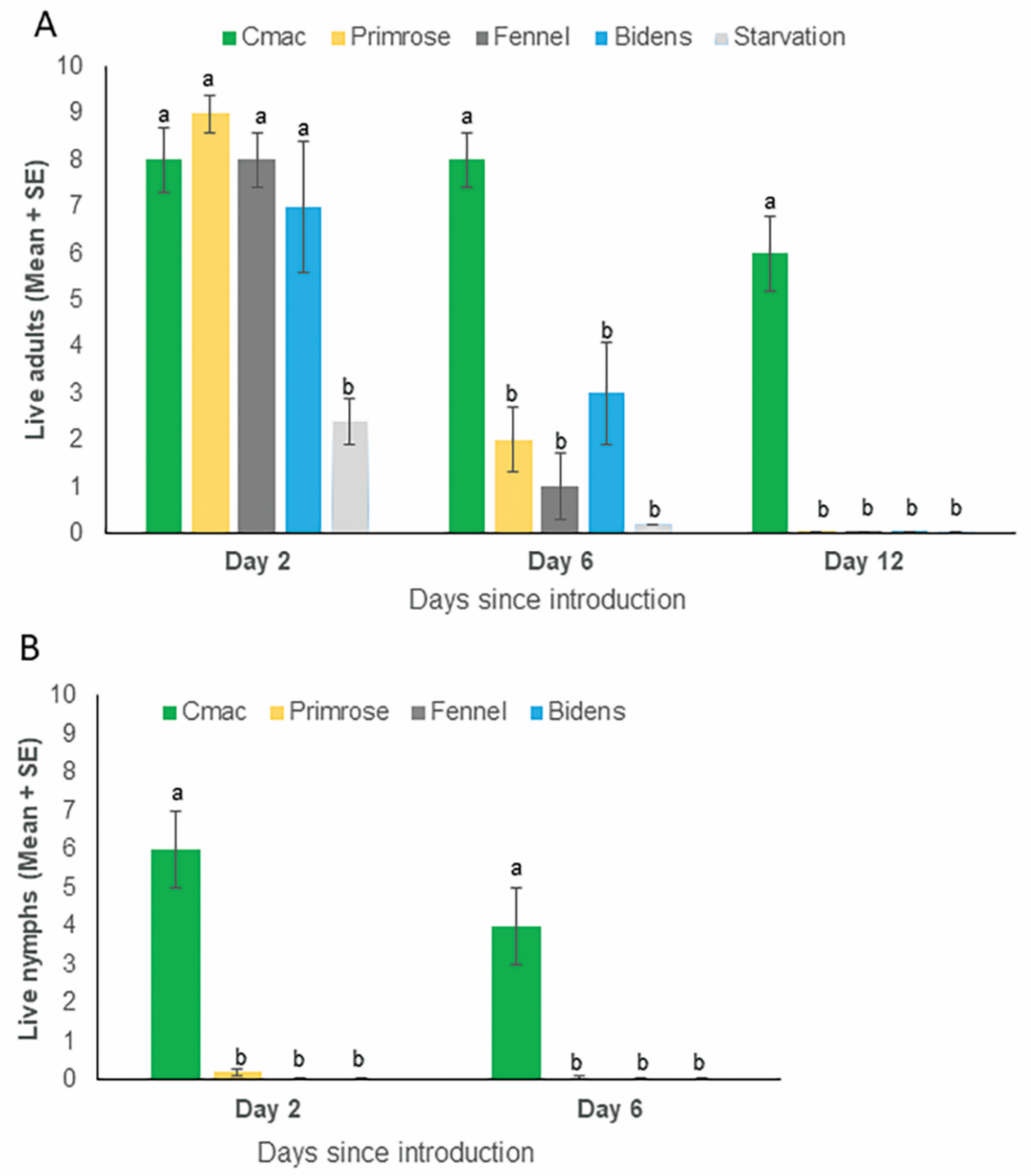
2.5. No Choice Assays to Study the Survival of D. citri Adults and Nymphs on Weed Plants
3. Results
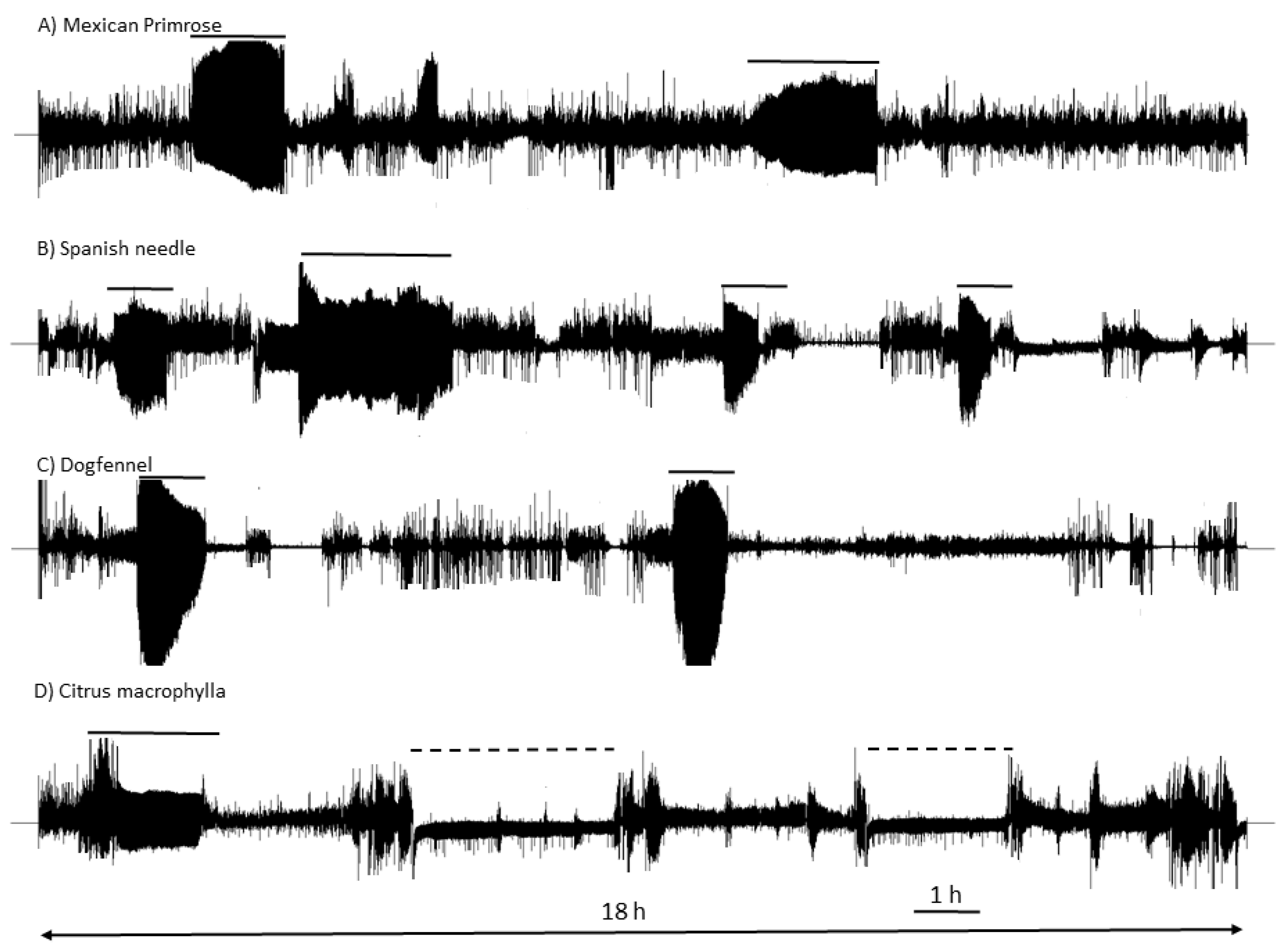
3.1. Electrical Penetration Graph Recordings of D. citri Nymphs and Adults
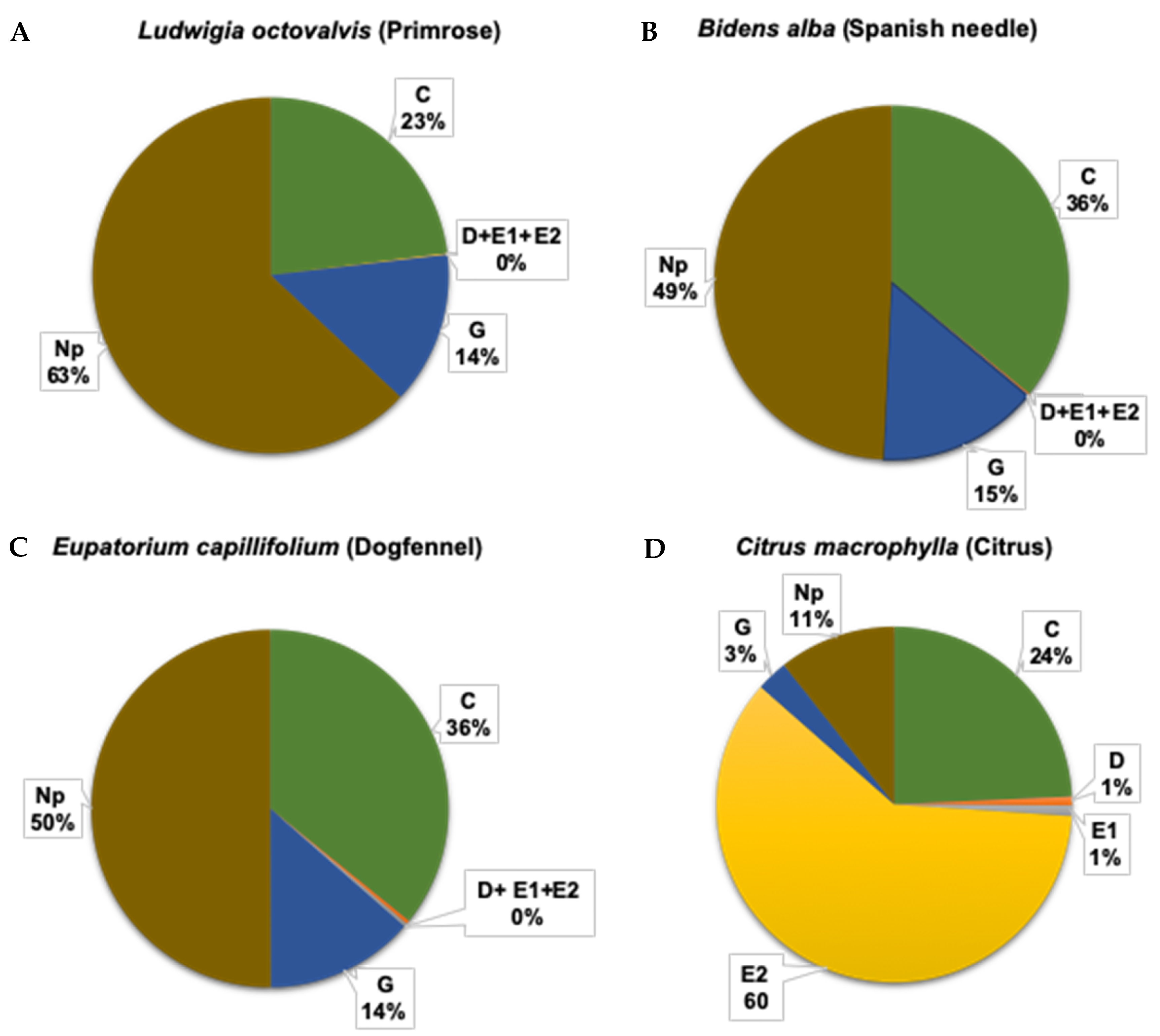
3.1.1. Frequency of Phloem Feeding Activities is Higher in Citrus than in Weeds
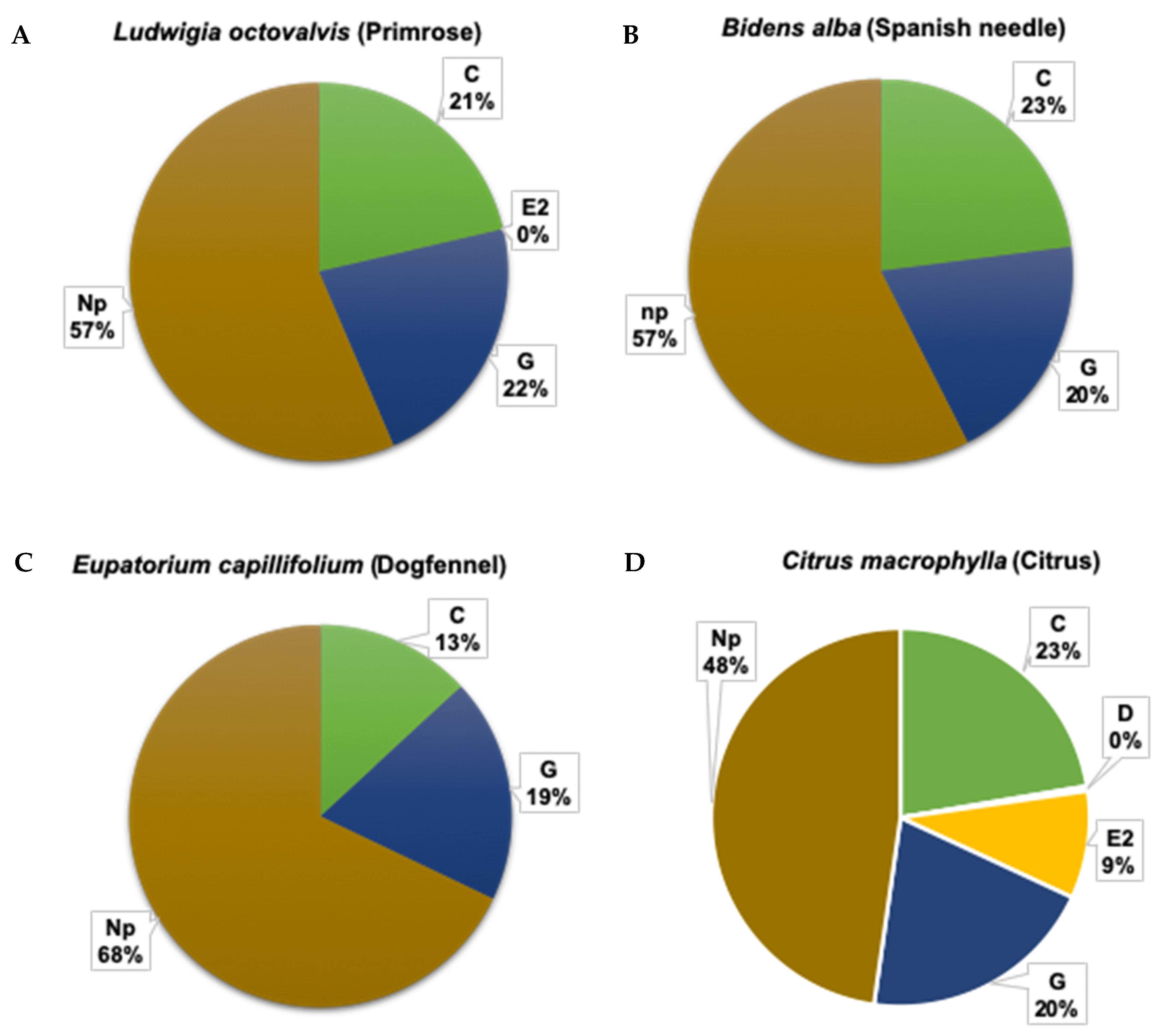
3.1.2. Duration of Xylem Feeding Activities is Similar on Weed Species Compared to Citrus
3.1.3. Total Duration (Frequency × Mean Duration) of Phloem and Xylem Feeding Activities
3.2. Visualization of Salivary Sheaths Produced by D. citri Adults
3.3. Survival of Psyllid Adults and Nymphs on Weed Species and Citrus in No Choice Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bove, J.M. Huanglongbing: A destructive, newly emerging, century-old disease of citrus. J. Plant. Pathol. 2006, 88, 7–37. [Google Scholar]
- Gottwald, T.R. Current epidemiological understanding of citrus huanglongbing. Ann. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.G.; Richardson, M.L.; Ammar, E.-D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae), vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Stansly, P.A.; Arevalo, H.A.; Qureshi, J.A.; Jones, M.M.; Hendricks, K.; Roberts, P.D.; Roka, F.M. Vector control and foliar nutrition to maintain economic sustainability of bearing citrus in Florida groves affected by huanglongbing. Pest. Manag. Sci. 2014, 70, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, J.A.; Kostyk, B.C.; Stansly, P.A. Insecticidal suppression of Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) vector of Huanglongbing pathogens. PLoS ONE 2014, 9, e112331. [Google Scholar] [CrossRef] [Green Version]
- Monzo, C.; Stansly, P.A. Thresholds for vector control and compatibility with beneficial fauna in citrus with high incidence of Huanglongbing. Acta Hortic. 2015, 1065, 1137–1143. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. A Review of the Citrus Greening Research and Development Efforts Supported by the Citrus Research and Development Foundation: Fighting a Ravaging Disease; The National Academies Press: Washington, DC, USA, 2018. [Google Scholar] [CrossRef]
- Johnston, N.J.; Stelinski, L.L.; Stansly, P.A. Dispersal patterns of Diaphorina citri (Kuwayama) as influenced by citrus grove management and abiotic factors. Fla. Entomol. 2018, 102, 168–173. [Google Scholar]
- Lewis-Rosenblum, H.; Martini, X.; Tiwari, S.; Stelinski, L.L. Seasonal movement patterns and long-range dispersal of Asian citrus psyllid in Florida citrus. J. Econ. Entomol. 2015, 108, 3–10. [Google Scholar] [CrossRef]
- Johnston, N.; Stansly, P.A.; Stelinski, L.L. Secondary hosts of the Asian citrus psyllid, Diaphorina citri (Kuwayama): Survivorship and preference. J. Appl. Entomol. 2019, 43, 921–928. [Google Scholar] [CrossRef]
- Futch, S.H.; Singh, M. Florida Citrus Pest Management Guide, Florida Citrus Pest Management Guide: Weeds; Rogers, M.E., Dewdney, M.M., Spann, T.M., Eds.; The University of Florida’s Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2010; pp. 125–137. [Google Scholar]
- USA Department of Agriculture–Natural Resources Conservation Service. The PLANTS Database; National Plant Data Team: Greensboro, NC, USA, 2010. Available online: http://plants.usda.gov (accessed on 3 November 2019).
- Buker, R.S. What You Should Know Before Planning Your Citrus Weed Management Program; Horticultural Sciences Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2005; Available online: http://edis.ifas.ufl.edu/pdffiles/HS/HS23500.pdf (accessed on 12 November 2019).
- Ramirez, A.H.M.; Amit, J.J.; Singh, M. Germination and Emergence Characteristics of Common Beggar’s-Tick (Bidens Alba). Weed Sci. 2012, 60, 374–378. [Google Scholar] [CrossRef]
- Hall, D.W.; Vandiver, V.V.; Ferell, J.A. Common Beggar’s-Tick (Hairy Beggar’s-Tick), Bidens alba (L.) D.C.; Agronomy Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2005; Available online: http://edis.ifas.ufl.edu/pdffiles/FW/FW00500.pdf (accessed on 23 October 2019).
- Sellers, B.A.; Ferrell, J.A.; MacDonald, G.E.; Kline, W.N. Dogfennel (Eupatorium capillifolium) size at application affects herbicide efficacy. Weed Technol. 2009, 23, 247–250. [Google Scholar] [CrossRef]
- Mohankumar, B.; Alexander, D. Influence of water regimes on weed growth and yields of transplanted rice. Oryza 1989, 26, 103–105. [Google Scholar]
- Angeles, G. The periderm of flooded and non-flooded Ludwigia octovalvis (Onagraceae). IAWA Bull. 1992, 13, 195–200. [Google Scholar] [CrossRef]
- Tjallingii, W. Electronic recording of penetration behaviour by aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- Futch, S.H.; Hall, D.W. Identification of Broadleaf Weeds in Citrus; Horticultural Science Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2003; Available online: https://edis.ifas.ufl.edu/pdffiles/HS/HS15000.pdf (accessed on 23 October 2019).
- Skelley, L.H.; Hoy, M.A. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biol. Control 2004, 29, 14–23. [Google Scholar] [CrossRef]
- Li, W.B.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus Huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Luo, X.; Yen, A.L.; Powell, K.S.; Wu, F.; Wang, Y.; Xeng, L.; Yang, Y.; Cen, Y. Feeding behavior of Diaphorina citri (Hemiptera: Liviidae) and its acquisition of ‘Candidatus Liberibacter asiaticus’, on huanglongbing-infected Citrus reticulata leaves of several maturity stages. Fla. Entomol. 2015, 98, 186–192. [Google Scholar] [CrossRef]
- George, J.; Ammar, E.-D.; Hall, D.G.; Lapointe, S.L. Sclerenchymatous ring as a barrier to phloem feeding by Asian citrus psyllid: Evidence from electrical penetration graph and visualization of stylet pathways. PLoS ONE 2017, 12, e0173520. [Google Scholar] [CrossRef]
- George, J.; Shi, Q.; Stelinski, L.L.; Stover, E.; Lapointe, S.L. Formic acid induced release of citrus volatiles influence the host selection, oviposition and feeding behavior of Diaphorina citri. Front. Ecol. Evol. 2019, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- George, J.; Ammar, E.-D.; Hall, D.G.; Lapointe, S.L. Prolonged phloem feeding activities by Diaphorina citri nymphs may explain their greater acquisition of citrus greening pathogen. Sci. Rep. 2018, 8, 10352. [Google Scholar] [CrossRef]
- Bonani, J.P.; Fereres, A.; Garzo, E.; Miranda, M.P.; Appezzato-Da-Gloria, B. Characterization of electrical penetration graphs of the Asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomol. Exp. Appl. 2010, 134, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Ammar, E.-D.; Hall, D.G.; Shatters, R.G., Jr. Stylet morphometrics and citrus leaf vein structure in relation to feeding behavior of the Asian citrus psyllid Diaphorina citri, vector of citrus huanglongbing bacterium. PLoS ONE 2013, 8, e59914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, J.G.; Agrawal, A.A. Asymmetry of plant-mediated interactions between specialist aphids and caterpillars on two milkweeds. Funct. Ecol. 2014, 28, 1404–1412. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Natural enemy responses and pest control: Importance of local vegetation. Biol. Control 2010, 52, 160–166. [Google Scholar] [CrossRef]
- Altieri, M.A.; Whitcomb, W.H. The potential use of weeds in the manipulation of beneficial insects. HortScience 1979, 14, 12–18. [Google Scholar]
- Al-Doghairi, M.A.; Cranshaw, W.H. The effect of interplanting of nectariferous plants on the population density and parasitism of cabbage plants. Southwest. Entomol. 2004, 29, 61–68. [Google Scholar]
- Showler, A.T.; Greenberg, S.M. Effects of weeds on selected arthropod herbivore and natural enemy populations, and on cotton growth and yield. Environ. Entomol. 2003, 32, 39–50. [Google Scholar] [CrossRef]
- Colloff, M.J.; Lindsay, E.A.; Cook, D.C. Natural pest control in citrus as an ecosystem service: Integrating ecology, economics and management at the farm scale. Biol. Control 2013, 67, 170–177. [Google Scholar] [CrossRef]
- Ortega, A.L.D.; Villegas, A.M.; Ramírez, A.J.R.; Mendoza, E.E.G. Abundancia estacional de Diaphrina citri (Hemiptera: Liviidae) en plantaciones de cítricos en Cazones, Veracruz, México. Acta Zool. México 2013, 29, 317–333. [Google Scholar]
- Almaguer-Vargas, G.; Botello-Castillo, M.D.; Jacobo-Blas, L.; Mauricio-Perez, J.M.; Gonzalez-Hernandez, V.A.; Cruz-Huerta, N.; Ramirez, R.I.; Mondragon-Pederero, G.; De La Cruz, D.E. The contribution of weeds in reducing Diaphorina citri (Asian citrus psyllid) populations on mandarin trees, in Tuxpan, Veracruz, Mexico. Appl. Ecol. Environ. Res. 2018, 16, 7361–7375. [Google Scholar] [CrossRef]
- Capinera, J. Relationships between insect pests and weeds: An evolutionary perspective. Weed Sci. 2005, 53, 892–901. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Stelinski, L.L.; Hall, D.G. The roles of olfactory cues, visual cues, and mating status in orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to four different host plants. Environ. Entomol. 2009, 38, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostal, V.; Finch, S. Preference of the cabbage root fly, Delia radicum (L.), for colored traps: Influence of sex and physiological status of the flies, trap background and experimental design. Physiol. Entomol. 1996, 21, 123–130. [Google Scholar] [CrossRef]
- Sétamou, M.; Simpson, C.R.; Alabi, O.J.; Nelson, S.D.; Telagamsetty, S.; Jifon, J.L. Quality matters: Influence of citrus flush physiochemical characteristics on population dynamics of the Asian citrus psyllid (Hemiptera: Liviidae). PLoS ONE 2016, 11, e0168997. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Psyllid Life Stage | Plant Type | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Waveform | Effect | Nymph | Adult | Primrose | Spanish Needle | Dogfennel | Citrus macrophylla | F-Value | P > F |
| C Intercellular passage | Stage | 29 ± 2 | 29 ± 2 | 0.003 | 0.95 | ||||
| Plant type | 30 ± 3 | 32 ± 3 | 23 ± 3 | 32 ± 2 | 3.21 | 0.02 | |||
| Interaction | 3.90 | 0.01 | |||||||
| D Phloem penetration | Stage | 3.4 ± 0.4 | 0.7 ± 0.4 | 21.3 | <0.0001 | ||||
| Plant type | 0.09 ± 0.5 | 0.6 ± 0.5 | 1.5 ± 0.5 | 5.1 ± 0.4 | 21.2 | <0.0001 | |||
| Interaction | 4.6 | 0.004 | |||||||
| E1 Phloem salivation | Stage | 3.4 ± 0.4 | 0.7 ± 0.4 | 20.6 | <0.0001 | ||||
| Plant type | 0.09 ± 0.5 | 0.6 ± 0.5 | 1.5 ± 0.5 | 5.2 ± 0.4 | 21.4 | <0.0001 | |||
| Interaction | 6.1 | 0.006 | |||||||
| E2 Phloem ingestion | Stage | 1.4 ± 0.1 | 0.3 ± 0.1 | 22.64 | <0.0001 | ||||
| Plant type | 0.08 ± 0.2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 2.8 ± 0.2 | 58.6 | <0.0001 | |||
| Interaction | 21.9 | <0.0001 | |||||||
| G Xylem feeding | Stage | 6 ± 1 | 7 ± 1 | 0.77 | 0.38 | ||||
| Plant type | 6.9 ± 1 | 9.6 ± 1 | 6.8 ± 1 | 4.3 ± 1 | 5.1 | 0.002 | |||
| Interaction | 2.0 | 0.11 | |||||||
| Np Non-probing activities | Stage | 20 ± 1 | 22 ± 1 | 0.84 | 0.36 | ||||
| Plant type | 23 ± 2 | 22 ± 2 | 15 ± 2 | 23 ± 2 | 3.6 | 0.01 | |||
| Interaction | 4.4 | 0.005 | |||||||
| Psyllid Life Stage | Plant Type | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Waveform | Effect | Nymph | Adult | Primrose | Spanish Needle | Dogfennel | Citrus macrophylla | F-Value | P > F |
| C Intercellular passage | Stage | 12.4 ± 0.6 | 8.9 ± 0.6 | 16.1 | <0.001 | ||||
| Plant type | 9.3 ± 0.9 | 11.2 ± 0.9 | 12.3 ± 0.9 | 9.8 ± 0.8 | 2.06 | 0.10 | |||
| Interaction | 4.4 | 0.005 | |||||||
| D Phloem penetration | Stage | 0.8 ± 0.05 | 0.16 ± 0.05 | 80.7 | <0.0001 | ||||
| Plant type | 0.06 ± 0.07 | 0.4 ± 0.07 | 0.5 ± 0.07 | 0.8 ± 0.06 | 22.7 | <0.0001 | |||
| Interaction | 6.7 | 0.0003 | |||||||
| E1 Phloem salivation | Stage | 0.8 ± 0.1 | 0.26 ± 0.1 | 17.6 | <0.0001 | ||||
| Plant type | 0.05 ± 0.1 | 0.3 ± 0.1 | 0.4 ± 0.1 | 1.1 ± 0.1 | 13.6 | <0.0001 | |||
| Interaction | 1.3 | 0.26 | |||||||
| E2 Phloem ingestion | Stage | 67.9 ± 10 | 13.5 ± 10 | 8.8 | 0.004 | ||||
| Plant type | 0.2 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 133 ± 13 | 8.8 | <0.0001 | |||
| Interaction | 23.9 | <0.0001 | |||||||
| G Xylem feeding | Stage | 21.3 ± 4 | 39.5 ± 3.9 | 8.5 | 0.004 | ||||
| Plant type | 37.3 ± 5.8 | 32.9 ± 5.6 | 32.3 ± 5.9 | 22.8 ± 5.1 | 1.26 | 0.29 | |||
| Interaction | 2.09 | 0.10 | |||||||
| Np Non-probing activities | Stage | 30 ± 3 | 41 ± 3 | 5.10 | 0.03 | ||||
| Plant type | 41 ± 5 | 36.4 ± 5 | 54.6 ± 5 | 17.2 ± 4 | 11.85 | <0.0001 | |||
| Interaction | 3.84 | 0.01 | |||||||
| Psyllid Life Stage | Plant Type | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Waveform | Effect | Nymph | Adult | Primrose | Spanish Needle | Dogfennel | Citrus macrophylla | F-Value | P > F |
| C Intercellular passage | Stage | 316 ± 13 | 220 ± 12 | 32.37 | <0.0001 | ||||
| Plant type | 239 ± 19 | 308 ± 18 | 268 ± 19 | 252 ± 17 | 3.40 | 0.02 | |||
| Interaction | 8.35 | <0.0001 | |||||||
| D Phloem penetration | Stage | 4 ± 0.5 | 0.6 ± 0.4 | 24.6 | <0.0001 | ||||
| Plant type | 0.09 ± 0.6 | 0.7 ± 0.6 | 1.8 ± 0.6 | 5.4 ± 0.5 | 17.1 | <0.0001 | |||
| Interaction | 5.4 | 0.001 | |||||||
| E1 Phloem salivation | Stage | 4.3 ± 0.4 | 0.6 ± 0.4 | 28.6 | <0.0001 | ||||
| Plant type | 0.08 ± 0.6 | 0.5 ± 0.6 | 1.6 ± 0.6 | 6.3 ± 0.5 | 25.08 | <0.0001 | |||
| Interaction | 10.09 | <0.0001 | |||||||
| E2 Phloem ingestion | Stage | 202 ± 12 | 28 ± 12 | 64.1 | <0.0001 | ||||
| Plant type | 0.6 ± 0.1 | 0.00 ± 0.0 | 0.0 ± 0.0 | 376 ± 15 | 133.4 | <0.0001 | |||
| Interaction | 72.1 | <0.0001 | |||||||
| G Xylem feeding | Stage | 113 ± 19 | 218 ± 18 | 13.68 | 0.0003 | ||||
| Plant type | 197 ± 27 | 188 ± 26 | 174 ± 28 | 125 ± 24 | 1.53 | 0.20 | |||
| Interaction | 1.59 | 0.19 | |||||||
| Np Non-probing activities | Stage | 437 ± 21 | 611 ± 20 | 28.56 | <0.0001 | ||||
| Plant type | 642 ± 30 | 582 ± 29 | 633 ± 30 | 315 ± 26 | 32.74 | <0.0001 | |||
| Interaction | 12.61 | <0.0001 | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, J.; Kanissery, R.; Ammar, E.-D.; Cabral, I.; Markle, L.T.; Patt, J.M.; Stelinski, L.L. Feeding Behavior of Asian Citrus Psyllid [Diaphorina citri (Hemiptera: Liviidae)] Nymphs and Adults on Common Weeds Occurring in Cultivated Citrus Described Using Electrical Penetration Graph Recordings. Insects 2020, 11, 48. https://doi.org/10.3390/insects11010048
George J, Kanissery R, Ammar E-D, Cabral I, Markle LT, Patt JM, Stelinski LL. Feeding Behavior of Asian Citrus Psyllid [Diaphorina citri (Hemiptera: Liviidae)] Nymphs and Adults on Common Weeds Occurring in Cultivated Citrus Described Using Electrical Penetration Graph Recordings. Insects. 2020; 11(1):48. https://doi.org/10.3390/insects11010048
Chicago/Turabian StyleGeorge, Justin, Ramdas Kanissery, El-Desouky Ammar, Itze Cabral, Larry T. Markle, Joseph M. Patt, and Lukasz L. Stelinski. 2020. "Feeding Behavior of Asian Citrus Psyllid [Diaphorina citri (Hemiptera: Liviidae)] Nymphs and Adults on Common Weeds Occurring in Cultivated Citrus Described Using Electrical Penetration Graph Recordings" Insects 11, no. 1: 48. https://doi.org/10.3390/insects11010048