Pest Management Challenges and Control Practices in Codling Moth: A Review

 , ,
, ,  , and
, and
Abstract
:1. Introduction
Origin and Biology of the Codling Moth, Cydia pomonella
2. Insecticides Resistance
3. Present Strategies in Codling Moth Suppression
3.1. Mechanical Control
3.2. Chemical Control
3.3. Biological Control
3.4. Population Genetic Monitoring
3.5. Area-Wide Integrated Pest Management
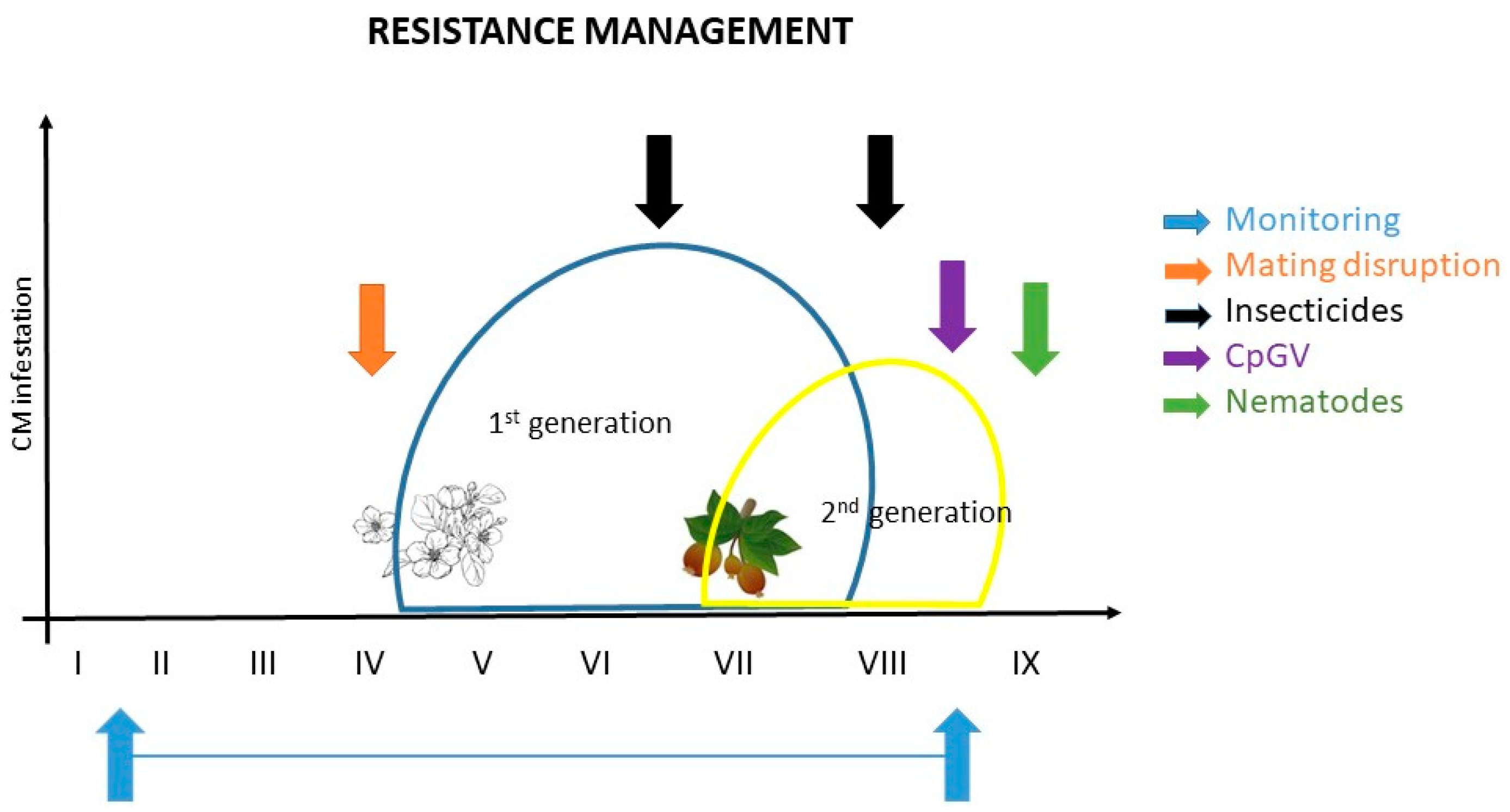
4. Resistance Management Strategies
5. Perspectives in Codling Moth Resistance Detection
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciglar, I. Integrirana Zaštita Voćaka i Vinove Loze, 1st ed.; Zrinski: Čakovec, Croatia, 1998; pp. 82–87. [Google Scholar]
- Balachowsky, A.; Mesnil, L. Les Insectes Nuisibles Aux Plantes Cultivées; Ministère de L’ Agriculture: Paris, France, 1935; pp. 130–158.
- Kovačević, Ž. Applied Entomology, 2nd ed.; University of Zagreb: Zagreb, Croatia, 1952; pp. 312–319. [Google Scholar]
- Slingerland, M.V. Codling moth in New England in 1750. N. Y. Agric. Exp. 1898, 142, 85–155. [Google Scholar]
- Zhang, X.Z. Taxonomic notes on the codling moth, Carpocapsa pomonella L. In Sinkiang. Acta Entomol. Sin. 1957, 7, 467–472. [Google Scholar]
- Franck, P.; Reyes, M.; Olivares, J.; Sauphanor, B. Genetic architecture in codling moth populations: Comparison between microsatellite and insecticide resistance markers. Mol. Ecol. 2007, 16, 3554–3564. [Google Scholar] [CrossRef] [PubMed]
- Thaler, R.; Brandstätter, A.; Meraner, A.; Chabicovski, M.; Parson, W.; Zelger, R.; Dalla Via, J.; Dallinger, R. Molecular phylogeny and population structure of the codling moth (Cydia pomonella) in Central Europe: II. AFLP analysis refl ects human-aided local adaptation of a global pest species. Mol. Phylogenet. Evol. 2008, 48, 838–849. [Google Scholar] [CrossRef]
- Maceljski, M. Jabučni savijač (Cydia/Laspeyresia, Carpocapsa, Grapholita/ pomonella L.). In Poljoprivredna Entomologija, 2nd ed.; Zrinski: Čakovec, Croatia, 2002; pp. 302–309. [Google Scholar]
- Alford, D.V. A Color Atlas of Fruit Pests Their Recognition, Biology, and Control; Wolfe Publishing: Prescott, AZ, USA, 1984. [Google Scholar]
- Wildbolz, T. Über Möglichkeiten der Prognose und Befallsüberwachung und Über Toleranzgrenzen bei der Integrierten Schädlingsbekämpfung im Obstbau. Entomophaga 1962, 7, 273–283. [Google Scholar] [CrossRef]
- Higbee, B.S.; Calkins, C.O.; Temple, C.A. Overwintering of codling moth (Lepidoptera: Tortricidae) larvae in apple harvest bins and subsequent moth emergence. J. Econ. Entomol. 2001, 94, 1511–1517. [Google Scholar] [CrossRef]
- Meraner, A.; Brandstätter, A.; Thaler, R.; Aray, B.; Unterlechner, M.; Niederstätter, H.; Parson, W.; Zelger, R.; Dalla Via, J.; Dallinger, R. Molecular phylogeny and population structure of the codling moth (Cydia pomonella) in Central Europe: I. Ancient clade splitting revealed by mitochondrial haplotype markers. Mol. Phylogenet. Evol. 2008, 48, 825–837. [Google Scholar] [CrossRef]
- Neven, L.G. Fate of codling moth (Lepidoptera: Tortricidae) in harvested apples held under short photoperiod. J. Econ. Entomol. 2012, 105, 297–303. [Google Scholar] [CrossRef]
- Neven, L.G. Effects of short photoperiod on codling moth diapause and survival. J. Econ. Entomol. 2013, 106, 520–523. [Google Scholar] [CrossRef]
- Pajač, I.; Barić, B.; Mikac, K.M.; Pejić, I. New insights into the biology and ecology of Cydia pomonella from apple orchards in Croatia. Bull. Insectol. 2012, 65, 185–193. [Google Scholar]
- Geier, P.W. The life history of codling moth, Cydia pomonella (L.) (Lepidoptera:Tortricidae), in the Australian capital territory. Aust. J. Zool. 1963, 11, 323–367. [Google Scholar] [CrossRef]
- Pajač, I.; Barić, B. The behaviour of codling moth (Lepidoptera: Tortricidae) in the Croatian apple orchards. IOBC/WPRS Bull. 2012, 74, 79–82. [Google Scholar]
- Sauphanor, B.; Brosse, V.; Bouvier, J.C.; Speich, P.; Micoud, A.; Martinet, C. Monitoring resistance to diflubenzuron and deltamethrin in French codling moth populations (Cydia pomonella). Pest Manag. Sci. 2000, 56, 74–82. [Google Scholar] [CrossRef]
- Boivin, T.; Chabert D’Hières, C.; Bouvier, J.C.; Beslay, D.; Sauphanor, B. Pleiotropy of insecticide resistance in the codling moth, Cydia pomonella. Entomol. Exp. Appl. 2001, 99, 381–386. [Google Scholar] [CrossRef]
- Bouvier, J.C.; Buès, R.; Boivin, T.; Boudinhon, L.; Beslay, D.; Sauphanor, B. Deltamethrin resistance in the codling moth (Lepidoptera: Tortricidae): Inheritance and number of genes involved. Heredity 2001, 87, 456–462. [Google Scholar] [CrossRef]
- May, R.M.; Dobson, A.P. Population dynamics and the rate of evolution of pesticide resistance. In Pesticide Resistance: Strategies and Tactics for Management; National Academy Press: Washington, DC, USA, 1986; pp. 170–193. [Google Scholar]
- Hough, W.S. Relative resistance to arsenical poisoning of two codling moth strains. J. Econ. Entomol. 1928, 21, 325–329. [Google Scholar] [CrossRef]
- Thwaite, W.G.; Williams, D.G.; Hately, A.M. Extent and significance of azinphos-methyl resistance in codling moth in Australia. Pest Control. Sustain. Agric. 1993, 93, 166–168. [Google Scholar]
- Sauphanor, B.; Bouvier, J.C.; Brosse, V. Spectrum of insecticide resistance in Cydia pomonella (Lepidoptera: Tortricidae) in South-eastern France. J. Econ. Entomol. 1998, 91, 1225–1231. [Google Scholar] [CrossRef]
- Reuveny, H.; Cohen, E. Resistance of the codling moth Cydia pomonella (L.) (Lep Tortricidae) to pesticides in Israel. J. Appl. Entomol. 2004, 128, 645–651. [Google Scholar] [CrossRef]
- Sauphanor, B.; Cuany, A.; Bouvier, J.C.; Brosse, V.; Amichot, M.; Berge, J.B. Mechanism of resistance to deltametrin in Cydia pomonella (L.) (Lepidoptera: Tortricidae). Pestic. Biochem. Physiol. 1997, 58, 109–117. [Google Scholar] [CrossRef]
- Stara, J.; Naďova, K.; Kocourek, F. Insecticide resistance in the codling moth (Cydia pomonella). J. Fruit Ornam. Plant Res. 2006, 14, 99–106. [Google Scholar]
- Reyes, M.; Franck, P.; Charmillot, P.J.; Ioriatti, C.; Olivares, J.; Pasqualini, E.; Sauphanor, B. Diversity of insecticide resistance mechanisms and spectrum in European populations of the codling moth, Cydia pomonella. Pest Manag. Sci. 2007, 63, 890–902. [Google Scholar] [CrossRef] [PubMed]
- Reyes, M.; Barros-Parada, W.; Ramírez, C.C.; Fuentes-Contreras, E. Organophosphate resistance and its main mechanism in populations of codling moth (Lepidoptera: Tortricidae) from Central Chile. J. Econ. Entomol. 2015, 108, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Bush, M.R.; Abdel-All, Y.A.; Rock, G.C. Parathion resistance and esterase activity in codling moth (Lepidoptera: Tortricidae) from North Carolina. J. Econ. Entomol. 1993, 86, 660–666. [Google Scholar] [CrossRef]
- Brun-Barale, A.; Bouvier, J.C.; Pauron, D.; Bergé, J.B.; Sauphanor, B. Involvement of a sodium channel mutation in pyrethroid resistance in Cydia pomonella L., and development of a diagnostic test. Pest Manag. Sci. 2005, 61, 549–554. [Google Scholar] [CrossRef]
- Cassanelli, S.; Reyes, M.; Rault, M.; Manicardi, G.C.; Sauphanor, B. Acetylcholinesterase mutation in an insecticide-resistant population of the codling moth Cydia pomonella (L.). Insect Biochem. Mol. Biol. 2006, 36, 642–653. [Google Scholar] [CrossRef]
- Pajač, I.; Barić, B.; Šimon, S.; Mikac, K.M.; Pejić, I. An initial examination of the population genetic structure of Cydia pomonella (Lepidoptera: Tortricidae) in Croatian apple orchards. J. Food Agric. Environ. 2011, 9, 459–464. [Google Scholar]
- Bosch, D.; Rodríguez, M.A.; Avilla, J. Monitoring resistance of Cydia pomonella (L.) Spanish field populations to new chemical insecticides and the mechanisms involved. Pest Manag. Sci. 2018, 74, 933–943. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.Q.; Zhang, Y.L. Investigation of insecticide-resistance status of Cydia pomonella in Chinese populations. Bull. Entomol. Res. 2015, 105, 316–325. [Google Scholar] [CrossRef]
- Voudouris, C.C.; Sauphanor, B.; Franck, P.; Reyes, M.; Mamuris, Z.; Tsitsipis, J.A.; Vontas, J.; Margaritopoulos, J.T. Insecticide resistance status of the codling moth Cydia pomonella (Lepidoptera: Tortricidae) from Greece. Pestic. Biochem. Physiol. 2011, 100, 229–238. [Google Scholar] [CrossRef]
- Herniou, E.A.; Luque, T.; Chen, X.; Vlak, J.M.; Winstanley, D.; Cory, J.S.; O’Reilly, D.R. Use of whole genome sequence data to infer baculovirus phylogeny. J. Virol. 2001, 75, 8117–8126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.L.; Hoover, K. Baculoviruses and other occluded insect viruses. In Insect Pathology, 2nd ed.; Vega, F.E., Kaya, H.K., Eds.; Academic Press: London, UK, 2012; pp. 73–131. [Google Scholar]
- Falcon, L.A.; Kane, W.R.; Bethell, R.S. Preliminary evaluation of a granulosis virus for control of the codling moth. J. Econ. Entomol. 1968, 61, 1208–1213. [Google Scholar] [CrossRef]
- Huber, J.; Dickler, E. Codling moth granulosis virus: It’s efficiency in the field in comparison with organophosphorus insecticides. J. Econ. Entomol. 1977, 70, 557–561. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Jehle, J.A. Field resistance of codling moth against Cydia pomonella granulovirus (CpGV) is autosomal and incompletely dominant inherited. J. Invertebr. Pathol. 2006, 93, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zingg, D.; Undorf-Spahn, K.; Fritsch, E.; Huber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Zichová, T.; Stará, J.; Kundu, J.K.; Eberle, K.E.; Jehle, J.A. Resistance to Cydia pomonella granulovirus follows a geographically widely distributed inheritance type within Europe. Biocontrol 2013, 58, 525–534. [Google Scholar] [CrossRef]
- Schulze-Bopp, S.; Jehle, J.A. Development of a direct test of baculovirus resistance in wild codling moth populations. J. Appl. Entomol. 2013, 137, 153–160. [Google Scholar] [CrossRef]
- Sauer, A.J.; Fritsch, E.; Undorf-Spahn, K.; Nguyen, P.; Marec, F.; Heckel, D.G.; Jehle, J.A. Novel resistance to Cydia pomonella granulovirus (CpGV) in codling moth shows autosomal and dominant inheritance and confers cross-resistance to different CpGV genome groups. PLoS ONE 2017, 12, e0179157. [Google Scholar] [CrossRef]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A third type of resistance to Cydia pomonella granulovirus in codling moths shows a mixed Z-linked and autosomal inheritance pattern. Appl. Environ. Microbiol. 2017, 83, e01036-17. [Google Scholar] [CrossRef] [Green Version]
- Alaphilippe, A.; Capowiez, Y.; Severac, G.; Simon, S.; Saudreau, M.; Caruso, S.; Vergnani, S. Codling moth exclusion netting: An overview of French and Italian experiences. IOBC-WPRS Bull. 2016, 112, 31–35. [Google Scholar]
- Pajač Živković, I.; Jemrić, T.; Fruk, M.; Buhin, J.; Barić, B. Influence of different netting structures on codling moth and apple fruit damages in northwest Croatia. Agric. Conspec. Sci. 2016, 81, 99–102. [Google Scholar]
- Tasin, M.; Demaria, D.; Ryne, C.; Cesano, A.; Galliano, A.; Anfora, G.; Ioriatti, C.; Alma, A. Effect of anti-hail nets on Cydia pomonella behavior in apple orchards. Entomol. Exp. Appl. 2008, 129, 32–36. [Google Scholar] [CrossRef]
- Sauphanor, B.; Severac, G.; Maugin, S.; Toubon, J.F.; Capowiez, Y. Exclusion netting may alter reproduction of the codling moth (Cydia pomonella) and prevent associated fruit damage to apple orchards. Entomol. Exp. Appl. 2012, 145, 134–142. [Google Scholar] [CrossRef]
- Baiamonte, I.; Raffo, A.; Nardo, N.; Moneta, E.; Peparaio, M.; D’Aloise, A.; Kelderer, M.; Casera, C.; Paoletti, F. Effect of the use of anti-hail nets on codling moth (Cydia pomonella) and organoleptic quality of apple (cv. Braeburn) grown in Alto Adige Region (northern Italy). J. Sci. Food Agric. 2016, 96, 2025–2032. [Google Scholar] [CrossRef]
- Sauer, A.J. Novel Types of Resistance of Codling Moth to Cydia pomonella Granulovirus. Ph.D. Thesis, Technische Universität, Darmstadt, Germany, 2017. [Google Scholar]
- Insecticide Resistance Action Committee (IRAC). Codling Moth, Cydia pomonella. Available online: https://www.irac-online.org/pests/cydia-pomonella (accessed on 20 August 2019).
- Beers, E.H.; Stuckling, D.M.; Prokopy, R.J.; Avila, J. Ecology and management of apple arthropod pests. In Apples: Botany, Production and Uses, 1st ed.; Ferree, D.C., Warrington, I.J., Eds.; CABI Publishing: Wallingford, UK, 2003; pp. 489–514. [Google Scholar]
- Arthropod Pesticide Resistance Database (APRD). Cydia pomonella-Shown Resistance to Active Ingredient(s). Available online: https://www.pesticideresistance.org/display.php?page=species&arId=407 (accessed on 30 August 2019).
- Gonzalez, D.C. Cydia pomonella (L.) Behavior and Responses to Host Volatiles. Ph.D. Thesis, Department de Quimica, Universitat de Lleida, Lleida, Spain, 2007. [Google Scholar]
- Lacey, L.A.; Thomson, D.; Vincent, C.; Arthurs, S.P. Codling moth granulovirus: A comprehensive review. Biocontrol Sci. Technol. 2008, 18, 639–663. [Google Scholar] [CrossRef]
- Czaja, K.; Góralczyk, K.; Struciński, P.; Hernik, A.; Korcz, W.; Minorczyk, M.; Łyczewska, M.; Ludwicki, J.K. Biopesticides-towards increased consumer safety in the European Union. Pest Manag. Sci. 2015, 71, 3–6. [Google Scholar] [CrossRef]
- Gerwick, B.C.; Sparks, T.C. Natural products for pest control: An analysis of their role, value and future. Pest Manag. Sci. 2014, 70, 1169–1185. [Google Scholar] [CrossRef]
- Bassi, A.; Rison, J.L.; Wiles, J.A. Chlorantraniliprole (DPX-E2Y45, Rynaxypyr®, Coragen®), a new diamide insecticide for control of codling moth (Cydia pomonella), Colorado potato beetle (Leptinotarsa decemlineata) and European grapevine moth (Lobesia botrana). Nova Goric. 2009, 4, 39–45. [Google Scholar]
- Pajač Živković, I.; Barić, B. Rezistentnost jabukova savijača na insekticidne pripravke. Glas. Biljn. Zaštite 2017, 17, 469–479. [Google Scholar]
- Lacey, L.A.; Unruh, T.R. Biological control of codling moth (Cydia pomonella, Lepidoptera: Tortricidae) and its role in integrated pest management, with emphasis on entomopathogens. Vedalia 2005, 12, 33–60. [Google Scholar]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Contr. 2001, 21, 230–248. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Arthurs, S.P.; Headrick, H. Comparative activity of the codling moth granulovirus against Grapholita molesta and Cydia pomonella (Lepidoptera: Tortricidae). J. Entomol. Soc. Br. Columbia 2005, 102, 79–80. [Google Scholar]
- Arthurs, S.P.; Lacey, L.A. Field evaluation of commercial formulations of the codling moth granulovirus (CpGV): Persistence of activity and success of seasonal applications against natural infestations in the Pacific Northwest. Biol. Control 2004, 31, 388–397. [Google Scholar] [CrossRef]
- Lacey, L.A.; Arthurs, S.P.; Knight, A.; Becker, K.; Headrick, H. Efficacy of codling moth granulovirus: Effect of adjuvants on persistence of activity and comparison with other larvicides in a Pacific Northwest apple orchard. J. Entomol. Sci. 2004, 39, 500–513. [Google Scholar] [CrossRef]
- Lacey, L.A.; Arthurs, S.P. New method for testing solar sensitivity of commercial formulations of the granulovirus of codling moth (Cydia pomonella, Tortricidae: Lepidoptera). J. Invertebr. Pathol. 2005, 90, 85–90. [Google Scholar] [CrossRef]
- Thorpe, P.T.; Pryke, J.S.; Samways, M.J. Review of ecological and conservation perspectives on future options for arthropod management in Cape Floristic Region pome fruit orchards. Afr. Entomol. 2016, 24, 279–306. [Google Scholar] [CrossRef]
- Georgis, R.; Koppenhöfer, A.; Lacey, L.; Belair, G.; Duncan, L.; Grewal, P.; Samish, M.; Torr, P.; van Tol, R. Successes and failures of entomopathogenic nematodes. Biol. Contr. 2006, 38, 103–123. [Google Scholar] [CrossRef]
- Lacey, L.A.; Unruh, T.R. Entomopathogenic nematodes for control of codling moth: Effect of nematode species, dosage, temperature and humidity under laboratory and simulated field conditions. Biol. Contr. 1998, 13, 190–197. [Google Scholar] [CrossRef]
- Lacey, L.A.; Granatstein, D.; Arthurs, S.P.; Headrick, H.; Fritts, J.R. Use of entomopathogenic nematodes (Steinernematidae) in conjunction with mulches for control of overwintering codling moth (Lepidoptera: Tortricidae). J. Entomol. Sci. 2006, 41, 107–119. [Google Scholar] [CrossRef]
- Glen, D.M. The effects of predators on the eggs of codling moth Cydia pomonella, in a cider apple orchard in Southwest England. Ann. Appl. Biol. 1975, 80, 115–119. [Google Scholar] [CrossRef]
- Wearing, C.H. Integrated control of apple pests in New Zealand: 10 Population dynamics of codling moth in Nelson. N. Z. J. Zool. 1979, 6, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Hogan, B.C. A Study of Bat Foraging Activity and Its Relation to Codling Moth Activity on Four Yolo County Walnut Orchards. Master’s Thesis, California State University, Sacramento, CA, USA, 2000. [Google Scholar]
- Riddick, E.W.; Mills, N.J. Potential of adult carabids (Coleoptera: Carabidae) as predators of fifth-instar codling moth (Lepidoptera: Tortricidae) in apple orchards in California. Environ. Entomol. 1994, 23, 1338–1345. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Altieri, M.A. Ecological engineering: A new direction for agricultural pest management. AFBM J. 2004, 1, 25–31. [Google Scholar]
- Granatstein, D.; Mullinix, K. Mulching options for Northwest organic and conventional orchards. HortScience 2008, 43, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Shapiro-Ilan, D.I.; Glenn, G.M. Post-application of anti-desiccant agents improves efficacy of entomopathogenic nematodes in formulated host cadavers or aqueous suspension against diapausing codling moth larvae (Lepidoptera: Tortricidae). Biocontrol Sci. Technol. 2010, 20, 909–921. [Google Scholar] [CrossRef]
- Navaneethan, T.; Strauch, O.; Besse, S.; Bonhomme, A.; Ehlers, R.U. Influence of humidity and a surfactant-polymer-formulation on the control potential of the entomopathogenic nematode Steinernema feltiae against diapausing codling moth larvae (Cydia pomonella L.) (Lepidoptera: Tortricidae). BioControl 2010, 55, 777–788. [Google Scholar] [CrossRef]
- De Waal, J.Y.; Malan, A.P.; Addison, M.F. Evaluating mulches together with Heterorhabditis zealandica (Rhabditida: Heterorhabditidae) for the control of diapausing codling moth larvae, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Biocontrol Sci. Technol. 2011, 21, 255–270. [Google Scholar] [CrossRef]
- Franck, P.; Timm, A.E. Population genetic structure of Cydia pomonella: A review and case study comparing spatiotemporal variation. J. Appl. Entomol. 2010, 134, 191–200. [Google Scholar] [CrossRef]
- Blommers, L.H. Integrated pest management in European apple orchards. Annu. Rev. Entomol. 1994, 39, 213–241. [Google Scholar] [CrossRef]
- Fuentes-Contreras, E.; Espinoza, J.L.; Lavandero, B.; Ramírez, C.C. Population genetic structure of codling moth (Lepidoptera: Tortricidae) from apple orchards in central Chile. J. Econ. Entomol. 2008, 101, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Roderick, G.K. Geographic structure of insect populations: Gene flow, phylogeography, and their uses. Annu. Rev. Entomol. 1996, 41, 325–352. [Google Scholar] [CrossRef] [PubMed]
- Keil, S.; Gu, H.; Dorn, S. Response of Cydia pomonella to selection on mobility: Laboratory evaluation and field verifi caion. Ecol. Entomol. 2001, 26, 495–501. [Google Scholar] [CrossRef]
- Dorn, S.; Schumacher, P.; Abivardy, C.; Meyhofer, R. Global and regional pest insects and their antagonists in orchards: Spatial dynamics. Agric. Ecosyst. Environ. 1999, 73, 111–118. [Google Scholar] [CrossRef]
- Pashley, D.P.; Bush, G.L. The use of allozymes in studying insect movement with special reference to the codling moth, Laspeyresia pomonella. In Movement of Highly Mobile Insects: Concepts and Methodology in Research, 1st ed.; Rabb, R.L., Kennedy, G.G., Eds.; North Carolina State University Press: Raleigh, NC, USA, 1979; pp. 333–341. [Google Scholar]
- Bues, R.; Toubon, J.F.; Poitout, H.S. Variabilite ecophysiologique et enzymatique de Cydia pomonella L. en fonction de l’origine geographique et de la plante hote. Agronomie 1995, 15, 221–231. [Google Scholar] [CrossRef]
- Timm, A.E.; Geertsema, H.; Warnich, L. Gene flow among Cydia pomonella (Lepidoptera: Tortricidae) geographic and host populations in South Africa. J. Econ. Entomol. 2006, 99, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Gu, H.N.; Dorn, S. Isolation of microsatellite loci in the codling moth Cydia pomonella (Lepidoptera: Tortricidae). Mol. Ecol. Notes 2005, 5, 226–227. [Google Scholar] [CrossRef]
- Franck, P.; Guérin, F.; Loiseau, A.; Sauphanor, B. Isolation and characterization of microsatellite loci in the codling moth Cydia pomonella (Lepidoptera: Tortricidae). Mol. Ecol. Notes 2005, 5, 99–102. [Google Scholar] [CrossRef]
- Chen, M.H.; Dorn, S. Microsatellites reveal genetic differentiation among populations in an insect species with high genetic variability in dispersal, the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2010, 100, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Contreras, E.; Basoalto, E.; Franck, P.; Lavandero, B.; Knight, A.L.; Ramírez, C.C. Measuring local genetic variability in populations of codling moth (Lepidoptera: Tortricidae) across an unmanaged and commercial orchard interface. Environ. Entomol. 2014, 43, 520–527. [Google Scholar] [CrossRef]
- Men, Q.L.; Chen, M.H.; Zhang, Y.L.; Feng, J.N. Genetic structure and diversity of a newly invasive species, the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae) in China. Biol. Invasions 2013, 15, 447–458. [Google Scholar] [CrossRef]
- Voudouris, C.; Franck, P.; Olivares, J.; Sauphanor, B.; Mamuris, Z.; Tsitsipis, J.; Margaritopoulos, J. Comparing the genetic structure of codling moth Cydia pomonella (L.) from Greece and France: Long distance gene-flow in a sedentary pest species. Bull. Entomol. Res. 2012, 102, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Margaritopoulos, J.T.; Voudouris, C.C.; Olivares, J.; Sauphanor, B.; Mamuris, Z.; Tsitsipis, J.A.; Franck, P. Dispersal ability in codling moth: Mark-release-recapture experiments and kinship analysis. Agric. For. Entomol. 2012, 14, 399–407. [Google Scholar] [CrossRef]
- Knight, A.L. Codling moth areawide integrated pest management. In Areawide Pest Management: Theory and Implementation, 1st ed.; Koul, O., Cuperus, G., Elliott, N., Eds.; CAB International: Oxfordshire, UK, 2008; pp. 159–190. [Google Scholar]
- Witzgall, P.; Stelinski, L.; Gut, L.; Thomson, D. Codling moth management and chemical ecology. Annu. Rev. Entomol. 2008, 53, 503–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Bengtsson, M.; Witzgall, P. Behavioral response of female Codling Moths, Cydia Pomonella, to apple volatiles. J. Chem. Ecol. 1999, 25, 1343–1351. [Google Scholar] [CrossRef]
- Roelofs, W.; Comeau, A.; Hill, A.; Milicevic, G. Sex attractant of the codling moth: Characterization with electroantennogram technique. Science 1971, 174, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L.; Light, D.M. Timing of egg hatch by early-season codling moth (Lepidoptera: Tortricidae) predicted by moth catch in pear ester-and codlemone-baited traps. Can. Entomol. 2005, 137, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Reynolds, K.M.; Buttery, R.G.; Merrill, G.; Roitman, J.; et al. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Naturwissenschaften 2001, 88, 333–338. [Google Scholar] [CrossRef]
- Knight, A.L.; Hilton, R.; Light, D.M. Monitoring codling moth (Lepidoptera: Tortricidae) in apple with blends of ethyl (E,Z)-2,4-decadienoate and codlemone. Environ. Entomol. 2005, 34, 598–603. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, V.J.; Manning, L.A.; Cole, L.; Suckling, D.M.; El-Sayed, A.M. Efficacy of the pear ester as a monitoring tool for codling moth Cydia pomonella (Lepidoptera: Tortricidae) in New Zealand apple orchards. Pest Manag. Sci. 2008, 64, 209–214. [Google Scholar] [CrossRef]
- Joshi, N.K.; Hull, L.A.; Rajotte, E.G.; Krawczyk, G.; Bohnenblust, E. Evaluating sex-pheromone-and kairomone-based lures for attracting codling moth adults in mating disruption versus conventionally managed apple orchards in Pennsylvania. Pest Manag. Sci. 2011, 67, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Suckling, D.M.; Judd, G.J.R. Positive interaction of a feeding attractant and a host kairomone for trapping the codling moth, Cydia pomonella (L.). J. Chem. Ecol. 2007, 33, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L.; Light, D.M.; Trimble, R.M. Identifying (E)-4,8-dimethyl-1,3,7-nonatriene plus acetic acid as a new lure for male and female codling moth (Lepidoptera: Tortricidae). Environ. Entomol. 2011, 40, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Fernández, D.E.; Cichón, L.; Garrido, S.; Ribes-Dasi, M.; Avilla, J. Comparison of lures loaded with codlemone and pear ester for capturing codling moths, Cydia pomonella, in apple and pear orchards using mating disruption. J. Insect Sci. 2010, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Thomson, D.; Jenkins, J. Successes with area-wide mating disruption: Moving from crisis management to sustainable pheromone-based pest management. In Proceedings of the IOBC/WPRS Working Group Pheromones and Other Semiochemicals in Integrated Production, Bursa, Turkey, 1–5 October 2012; Tasin, M., Kovanci, O.B., Eds.; International Organization for Biological and Integrated Control of Noxious Animals and Plants (OIBC/OILB). West Palaearctic Regional Section (WPRS/SROP): Dijon, France, 2014. [Google Scholar]
- Casado, D.; Cave, F.; Welter, S. (Puffer®-CM Dispensers for mating disruption of codling moth: Area of influence and impacts on trap finding success by males. Puffer®-CM Dispensers for mating disruption of codling moth: Area of influence and impacts on trap finding success by males. IOBC/WPRS Bull. 2014, 99, 25–31. [Google Scholar]
- Ciglar, I.; Barić, B.; Tomšić, T.; Šubić, M. Control of codling moth (Cydia pomonella) by mating disruption technique. Agron. Glas. 2000, 63, 85–93. [Google Scholar]
- Barić, B.; Pajač Živković, I. The efficacy of mating disruption in the control of codling moth in Croatia, with special reference to the costs. Pomol. Croat. Glas. Hrvat. Agron. Društva 2017, 21, 125–132. [Google Scholar]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [Green Version]
- Lösel, P.M.; Penners, G.; Potting, R.P.; Ebbinghaus, D.; Elbert, A.; Scherkenbeck, J. Laboratory and field experiments towards the development of an attract and kill strategy for the control of the codling moth, Cydia pomonella. Entomol. Exp. Appl. 2000, 95, 39–46. [Google Scholar] [CrossRef]
- Charmillot, P.J.; Pasquier, D.; Scalco, A.; Hofer, D. Studies on the control of the codling moth Cydia pomonella L. using attractant-insecticide. Mitt. Schweiz. Entomol. Ges. 1996, 69, 431–439. [Google Scholar]
- Mansour, M. Attract and kill for codling moth Cydia pomonella (Linnaeus) (Lepidoptera: Tortricidae) control in Syria. J. Appl. Entomol. 2010, 134, 234–242. [Google Scholar] [CrossRef]
- Damos, P.; Colomar, L.A.; Ioriatti, C. Integrated fruit production and pest management in Europe: The apple case study and how far we are from the original concept? Insects 2015, 6, 626–657. [Google Scholar] [CrossRef] [PubMed]
- Klassen, W. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-wide Integrated Pest Management, 1st ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–68. [Google Scholar]
- Vreysen, M.J.B.; Robinson, A.S.; Hendrichs, J. Areawide control of insect pests. From research to field implementation. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 1st ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 351–353. [Google Scholar]
- Vreysen, M.J.B.; Carpenter, J.E.; Marec, F. Improvement of the sterile insect technique for codling moth Cydia pomonella (Linnaeus) (Lepidoptera Tortricidae) to facilitate expansion of field application. J. Appl. Entomol. 2010, 134, 165–181. [Google Scholar] [CrossRef]
- Bloem, K.A.; Bloem, S.; Carpenter, J.E. Impact of moth suppression/eradication programmes using the sterile insect technique or inherited sterility. In Sterile Insect Technique. PRINCIPLES and Practice in Area-Wide Integrated Pest Management, 1st ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 677–700. [Google Scholar]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Public Relations and Political Support in Area-Wide Integrated Pest Management Programmes that Integrate the Sterile Insect Technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 1st ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 524–545. [Google Scholar]
- Joshi, N.K.; Rajotte, E.G.; Naithani, K.J.; Krawczyk, G.; Hull, L.A. Population dynamics and flight phenology model of codling moth differ between commercial and abandoned apple orchard ecosystems. Front. Physiol. 2016, 7, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onstad, D. Major Issues in Insect Resistance Management. In Insect Resistance Management, Biology, Economics and Prediction, 2nd ed.; Onstad, D., Ed.; Academic Press: Cambridge, MA, USA, 2007; pp. 1–16. [Google Scholar]
- Yu, F.L.; Wu, G.; Liu, T.J.; Zhai, B.P.; Chen, F.J. Effects of irrigation on the performance of cotton bollworm, Helicoverpa armigera (Hübner) during different pupal stages. Int. J. Pest Manag. 2008, 54, 137–142. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Insecticide Resistance Action Committee (IRAC). Resistance Management for Sustainable Agriculture and Improved Public Health, 2nd ed. 2010. Available online: http://www.irac-online.org/wp-content/uploads/2009/09/VM-Layout-v2.6_LR.pdf (accessed on 20 August 2019).
- Hood, C.S. Geometric morphometric approaches to the study of sexual size dimorphism in mammals. Hystrix 2000, 11, 77–90. [Google Scholar]
- Levine, E.; Oloumi-Sadeghi, H. Western corn rootworm (Coleoptera: Chrysomelidae) larval injury to corn grown for seed production following soybeans grown for seed production. J. Econ. Entomol. 1996, 89, 1010–1016. [Google Scholar] [CrossRef]
- Bouyer, J.; Ravel, S.; Dujardin, J.P.; De Meeus, T.; Via, L.; Thévenon, S.; Guerrini, L.; Sidibé, I.; Solano, P. Population structuring of Glossina palpalis gambiensis (Diptera: Glossinidae) according to landscape fragmentation in the Mouhoun river, Burkina Faso. J. Med. Entomol. 2007, 44, 788–795. [Google Scholar] [CrossRef] [Green Version]
- Benitez, A.H.; Lemić, D.; Bažok, R.; Gallardo-Araya, M.C.; Mikac, M.K. Evolutionary Directional Asymmetry and Shape Variation in Diabrotica v. virgifera (Coleoptera: Chrysomelidae): An example using hind wings. Biol. J. Linn. Soc. 2014, 111, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Lemic, D.; Benitez, H.A.; Bazok, R. Intercontinental effect on sexual shape dimorphism and allometric relationships in the beetle pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Zool. Anz. 2014, 253, 203–206. [Google Scholar] [CrossRef]
- Mikac, K.M.; Douglas, J.; Spencer, J.L. Wing shape and size of the western corn rootworm (Coleoptera: Chrysomelidae) is related to sex and resistance to soybean-maize crop rotation. J. Econ. Entomol. 2013, 106, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Mikac, K.M.; Lemic, D.; Bažok, R.; Benítez, H.A. Wing shape changes: A morphological view of the Diabrotica virgifera virgifera European invasion. Biol. Invasions 2016, 18, 3401–3407. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Benítez, H.A.; Bažok, R. Changes in corn rootworm wing morphology are related to resistance development. J. Pest Sci. 2019, 92, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Pajač Živković, I.; Lemic, D.; Mešić, A.; Barić, B.; Órdenes, R.; Benítez, H.A. Effect of fruit host on wing morphology in Drosophila suzukii (Diptera: Drosophilidae): A first view using geometric morphometrics. Entomol. Res. 2018, 48, 262–268. [Google Scholar] [CrossRef]
- Benítez, H.A.; Lemic, D.; Püschel, T.A.; Gašparić, H.V.; Kos, T.; Barić, B.; Bažok, R.; Živković, I.P. Fluctuating asymmetry indicates levels of disturbance between agricultural productions: An example in Croatian population of Pterostichus melas melas (Coleptera: Carabidae). Zool. Anz. 2018, 276, 42–49. [Google Scholar] [CrossRef]
- Khaghaninia, S.; Mohammadi, S.A.; Sarafrazi, A.M.; Iraninejad, K.H.; Ebrahimi, E.; Zahiri, R. An analysis of seasonal dimorphism in codling moths, Cydia pomonella, from Iran using geometric morphometrics. Bull. Insectol. 2014, 67, 43–50. [Google Scholar]
- PajačŽivković, I.; Benitez, H.A.; Barić, B.; Bažok, R.; Drmić, Z.; Mrganić, M.; Lemić, D. Analysis of Croatian Cydia pomonella L. (Lepidoptera: Tortricidae) population variability by using geometric morphometrics. In Proceedings of the European Congress of Entomology, ECE, Napoli, Italy, 2–6 July 2018; Dallai, R., Ed.; Event Planet: Naples, Italy, 2018; pp. 180–181. [Google Scholar]
- Brumfield, R.T.; Beerli, P.; Nickerson, D.A.; Edwards, S.V. The utility of single nucleotide polymorphisms in inferences of population history. Trends Ecol. Evol. 2003, 18, 249–256. [Google Scholar] [CrossRef]
- Morin, P.A.; Luikart, G.; Wayne, R.K. SNPs in ecology, evolution and conservation. Trends Ecol. Evol. 2004, 19, 208–216. [Google Scholar] [CrossRef]
- Xing, C.; Schumacher, F.R.; Xing, G.; Lu, Q.; Wang, T.; Elston, R.C. Comparison of microsatellites, single-nucleotide polymorphisms (SNPs) and composite markers derived from SNPs in linkage analysis. BMC Genet. 2005, 6, S29. [Google Scholar] [CrossRef] [Green Version]
- Genissel, A.; Pastinen, T.; Dowell, A.; Mackay, T.F.; Long, A.D. No evidence for an association between common nonsynonymous polymorphisms in Delta and bristle number variation in natural and laboratory populations of Drosophila melanogaster. Genetics 2004, 166, 291–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coates, B.S.; Sumerford, D.V.; Miller, N.J.; Kim, K.S.; Sappington, T.W.; Siegfried, B.D.; Lewis, L.C. Comparative performance of single nucleotide polymorphism and microsatellite markers for population genetic analysis. J. Hered. 2009, 100, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsakiozi, P.; Evans, B.R.; Gloria-Soria, A.; Kamgang, B.; Mayanja, M.; Lutwama, J.; Le Goff, G.; Ayala, D.; Paupy, C.; Badolo, A.; et al. Population structure of a vector of human diseases: Aedes aegypti in its ancestral range, Africa. Ecol. Evol. 2018, 8, 7835–7848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saarman, N.P.; Opiro, R.; Hyseni, C.; Echodu, R.; Opiyo, E.A.; Dion, K.; Johnson, T.; Aksoy, S.; Caccone, A. The population genomics of multiple tsetse fly (Glossina fuscipes fuscipes) admixture zones in Uganda. Mol. Ecol. 2019, 28, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Francischini, F.J.; Cordeiro, E.M.; de Campos, J.B.; Alves-Pereira, A.; Viana, J.P.G.; Wu, X.; Wei, W.; Brown, P.; Joyce, A.; Murua, G.; et al. Diatraea saccharalis history of colonization in the Americas. The case for human-mediated dispersal. PLoS ONE 2019, 14, e0220031. [Google Scholar] [CrossRef]
- Yadav, S.; Stow, A.J.; Dudaniec, R.Y. Detection of environmental and morphological adaptation despite high landscape genetic connectivity in a pest grasshopper (Phaulacridium vittatum). Mol. Ecol. 2019, 8, 3395–3412. [Google Scholar] [CrossRef]


{kind=link}
| Insecticide Group | MoA [54] | Insecticide/Active Substance | Use Period (Approximate) | Resistance Development (Year of First Report/Region) |
|---|---|---|---|---|
| Inorganic/others | Arsenate | 1890s–1950s | 1928/USA | |
| Nicotine | 1960s | |||
| Chlorinated hydrocarbons | DDT | Mid 1940s–1970s | 1955/USA | |
| Thiodan/Endosulfan | 1960s–1970s | 1965/Syria | ||
| Organophosphates | 1B | Diazinon | 1950s–2000s | |
| Phosalone | 1960s–2000s | |||
| Azinphosmethyl | 1950s–present | 1991/USA | ||
| Chlorpyrifos-ethyl | 1960s–present | 2011/France, Spain | ||
| Chlorpyrifos-methyl | 1960s–present | |||
| Methidation | 1950s–1990s | |||
| Phosmet | 1970s–present | 1999/USA | ||
| Mevinphos | Mid 1950s–mid 1990s | |||
| Methomyl | 1970s–1990s | |||
| Oxamyl | Mid 1980s–1990s | |||
| Formetante hydrochloride | 1970s–1990s | |||
| Charbamates | 1A | Carbaryl | 1970s–present | 2012/Spain |
| Pyrethroids | 3A | Fenvalerate/Esfenvalerate | 1970s–present | |
| Permethrin | 1970s–present | |||
| Bifenthrin | 1980s–present | |||
| Deltametrin | 1970s–present | 2001/China | ||
| Flucythrinate | 1980s–present | |||
| Lambda-cyhalotrin | 1980s–present | 2008/USA | ||
| Gama-cyhalotrin | 1980s–present | |||
| Tau-fluvalinate | 1980s–present | |||
| Microbial insecticides | Bacillus thuringiensis sub sp. kurstaki | 1980s–present | ||
| Codling moth granulovirus (CpGV) | 1980s–present | 2007/Germany | ||
| Naturalites | 5 | Spinosad | 1990s–present | |
| Insect growth regulators | 15 | Benzonylureas (diflubenzuron, hexaflumuron, flufenoxuron, triflumuron, lufenuron, teflubenzuron) | 1970s–present | diflubenzuron/1988/USA triflumuron/1995/France teflubenzuron/1995/France flufenoxuron/2011/Spain |
| 7B | Fenoxycarb | 1980s–present | 2007/Czechoslovakia | |
| 18 | Tebufenozide | 1990s–present | 1995/France | |
| Methoxyfenozide | 1990s–present | 2008/USA | ||
| 7B | Pyriproxyfen | 2000–present | ||
| Nicotinoids | 4A | Acetamiprid | 1990s–present | 2010/USA |
| Thiacloprid | 2001–present | 2011/Spain | ||
| Thiamethoxam | 2001–present | |||
| Avermectins | 6 | Emamectin benzoate | 2000–present | |
| Anthranilic diamide insecticides | 28 | Chlorantraniliprole | 2007–present | |
| Spinosyns | 5 | Spinetoram | 2011–present |
| Natural Enemies | Organism/Family | Family/Species | CM Life Stage Attacked |
|---|---|---|---|
| Entomopathogenic organisms | Virus | Granulovirus (CpGV) | Neonate larvae |
| Bacteria | Bacillus thurigiensis | Neonate larvae | |
| Fungi | Beauveria bassiana | Cocooned overwintering larvae | |
| Nematodes | Steinernematidae | Cocooned overwintering larvae | |
| Heterorhabditidae | |||
| Predators | Anthocoridae | Orius insidiosus | Eggs and neonate larvae |
| Anthocoris musculus | |||
| Miridae | Hyaliodes harti | ||
| Phytocoris sp. | |||
| Diaphnidia sp. | |||
| Blepharidopterus angulatus | |||
| Deraeocoris spp. | |||
| Reduviidae | Mature larvae | ||
| Nabidae | |||
| Carabidae, Trogossitidae, Malachiidae, Staphylinidae, Cleridae, Cantharidae, Elateridae | Cocooned larvae | ||
| Formicidae | Mature larvae | ||
| Phlaeothripidae | Haplothrips faurei | Eggs | |
| Leptothrips mali | |||
| Dermaptera | Forficula auricularia | ||
| Parasitoids | Braconidae | Ascogaster quadridentata | Larvae |
| Microdes rufipes | |||
| Ichneumonidae | Mastrus ridibundus | Larvae and adults | |
| Liotryphon caudatus | |||
| Pimpla turionellae | Pupae | ||
| Trichogrammatidae | Trichogramma sp. | Eggs |
| 2000 | 2012 | 2017 | |
|---|---|---|---|
| No. of MoA available for codling moth control * | 8 | 10 | 11 |
| No. of individual insecticides available ** | High | Decreasing | Fewer |
| Use of semiochemicals (mating disruption) | Minor | Moderate | Increasing |
| Microbial insecticides | Minor | Moderate | Moderate |
| Biological control | Minor | Minor | Minor |
| Regulatory pressure | Low | High | Decreasing |
| Food chain pressure | Low | High | Decreasing |
| Field resistance issues **/*** | Moderate | Decreasing | Low |
| Resistance knowledge and investigation tools | Moderate | Increasing | High |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadoić Balaško, M.; Bažok, R.; Mikac, K.M.; Lemic, D.; Pajač Živković, I. Pest Management Challenges and Control Practices in Codling Moth: A Review. Insects 2020, 11, 38. https://doi.org/10.3390/insects11010038
Kadoić Balaško M, Bažok R, Mikac KM, Lemic D, Pajač Živković I. Pest Management Challenges and Control Practices in Codling Moth: A Review. Insects. 2020; 11(1):38. https://doi.org/10.3390/insects11010038
Chicago/Turabian StyleKadoić Balaško, Martina, Renata Bažok, Katarina M. Mikac, Darija Lemic, and Ivana Pajač Živković. 2020. "Pest Management Challenges and Control Practices in Codling Moth: A Review" Insects 11, no. 1: 38. https://doi.org/10.3390/insects11010038