Neo Sex Chromosomes, Colour Polymorphism and Male-Killing in the African Queen Butterfly, Danaus chrysippus (L.)

 and
and
Abstract
:1. Introduction
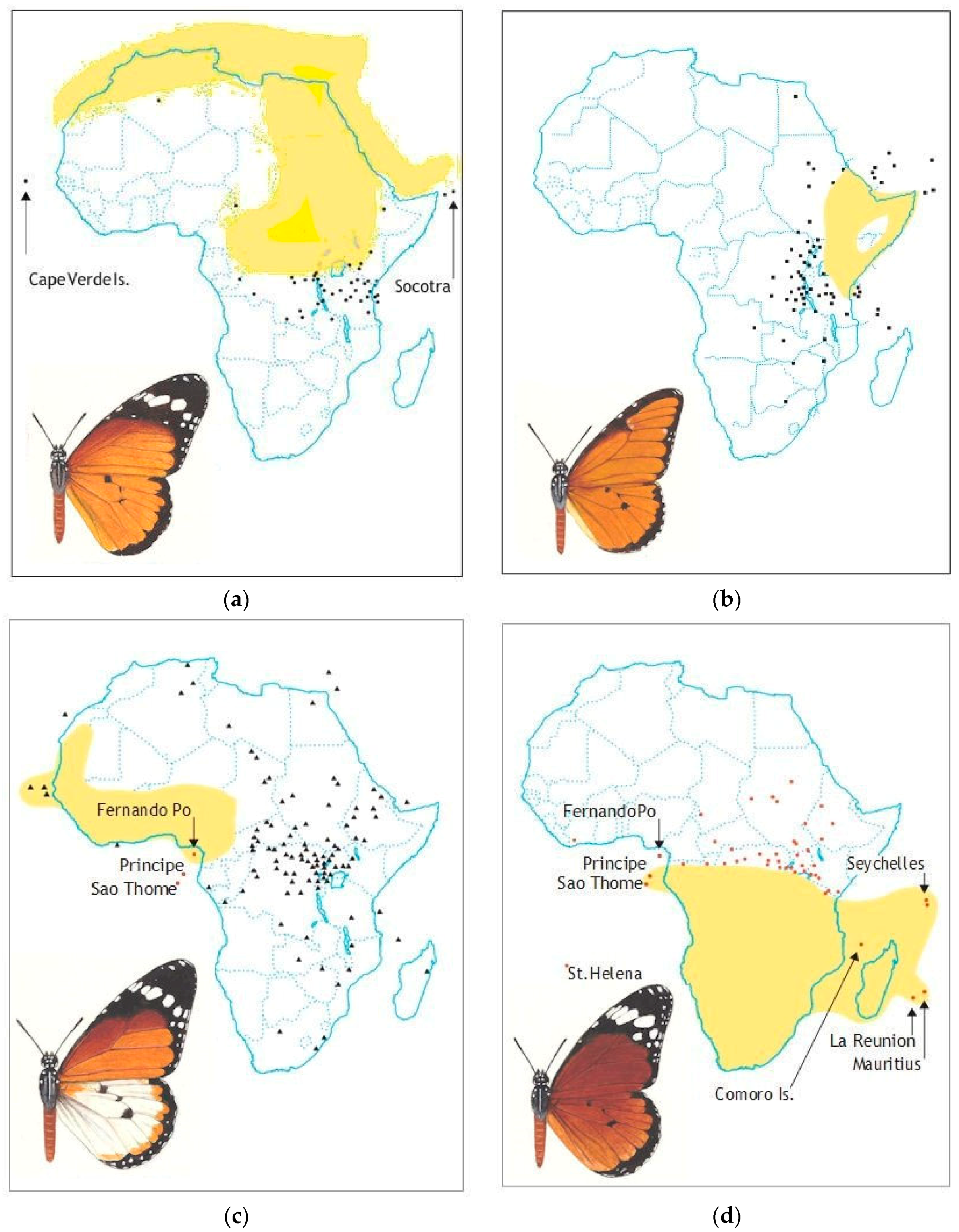
2. The Record
2.1. Host Plants
2.2. Cardenolide Glycosides
2.3. Pyrrolizidine Alkaloids
2.4. Courtship and Copulation
2.5. Defence
2.6. ‘Polymorphism’
2.7. Genetic Control of Colour Patterns
2.8. All-Female Broods
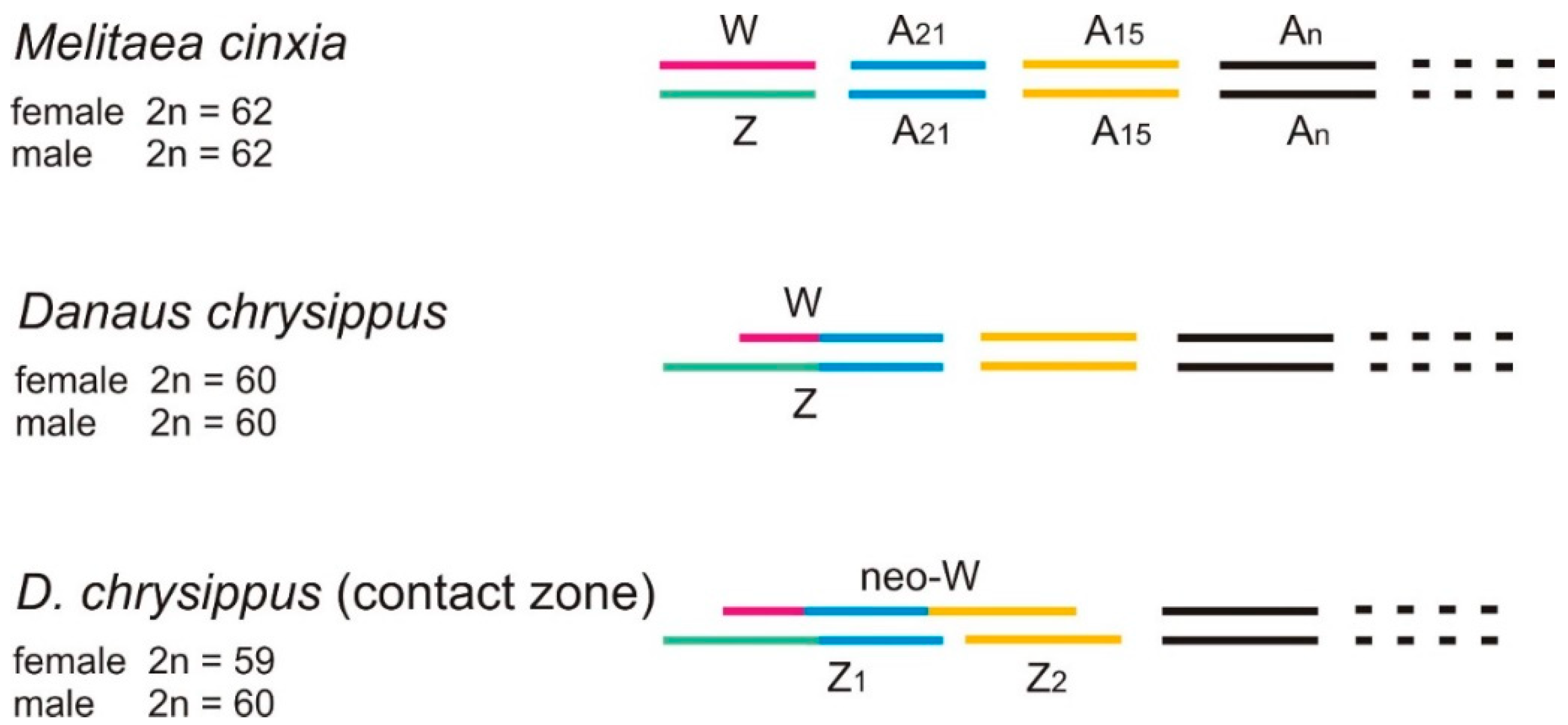
2.9. The neoW Chromosome
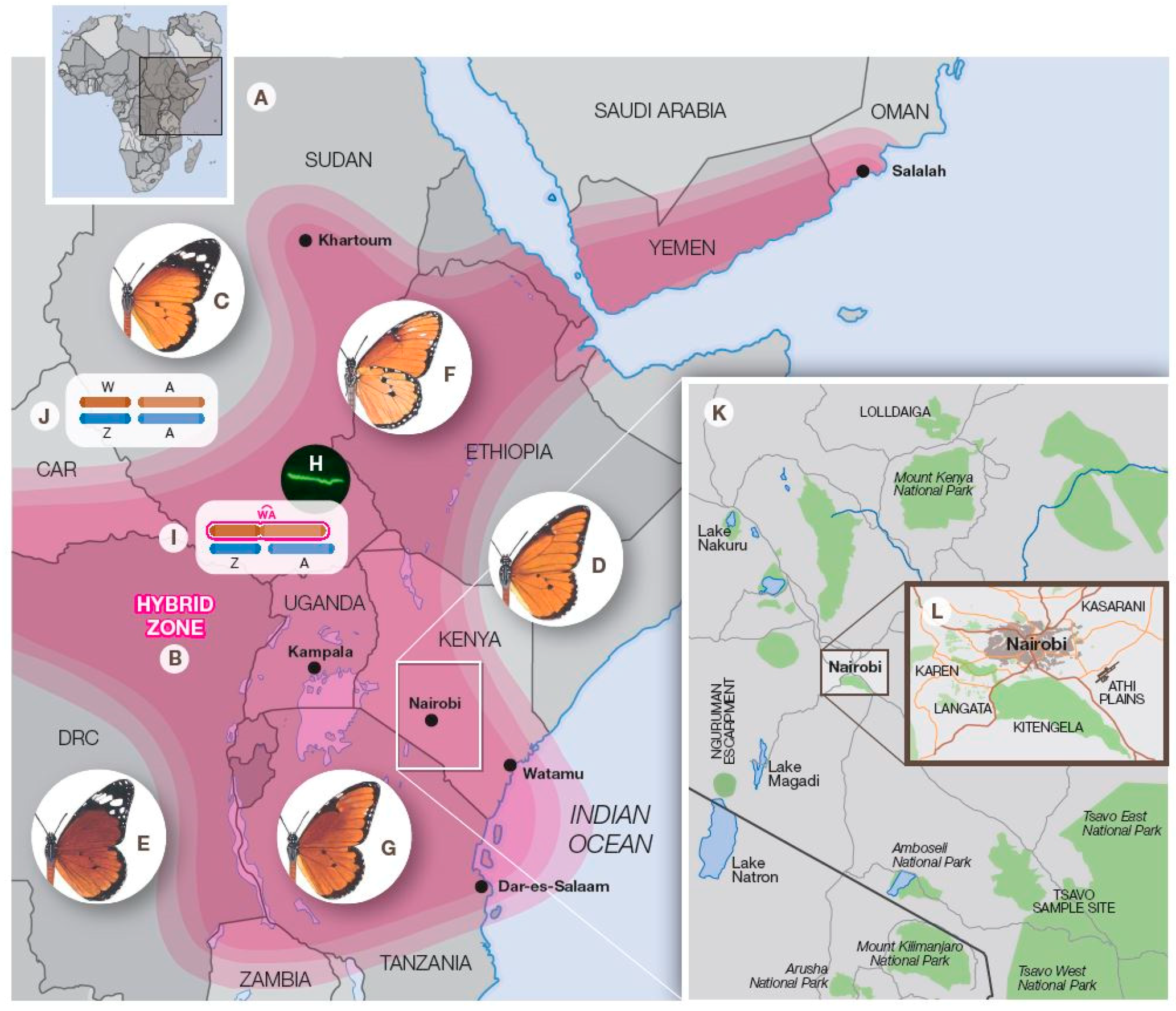
3. On-Going Hybrid Studies in the Nairobi Region, Kenya
3.1. Introduction
3.2. The Athi Plains
3.3. Kasarani
3.4. Kitengela
- (1)
- The population is ‘polymorphic’ for ssp. Dorippus (CC) and ssp. Chrysippus (cc). However, from the data in Table 4 it is clear, first, that the genotype arrays in two sexes differ sharply and indicate that they come predominantly from different source populations; secondly, much the most frequent female genotype is the hybrid form transiens (Cc).
- (2)
- The occasional arrival of males in numbers following substantial rains, unaccompanied by females of the same genotype, suggests that the migratory behaviour of the two sexes differs sharply [96].
- (3)
- Mating preference for C locus genotype was absent in both sexes, probably because the majority of males were CC and females Cc; thus, choice was severely curtailed.
- (4)
- It follows from (1) and (3) that pairing is disassortative (negatively non-random) for genotype (Table S2, p < 0.00001) since shortage of males effectively eliminates female choice, while male choice is restricted because 83% of females are transiens (Cc). Thus, ‘choice’ is in practice denied to both sexes.
- (5)
- Spermatophores per female averaged 1.7 (n = 260), while only 7.3% of females were unmated. These data mean that all females eventually find a mate. In a 1:1 population from Ghana, free from MK, the spermatophore/female average was 3.5 (n = 20) [8].
- (6)
- Through a mark-recapture study in May–July 2015, 63.4% of all males caught were in copula compared to only 13.4% of females. The implication of the sex difference is that, if all females are eventually mated (see 5), the average male must mate 5 times, but to achieve 1.7 spermatophores per female, males must mate 8.5 times, hence, neoW females are likely to receive under-sized spermatophores from over-taxed males.
- (7)
- The butterfly is a permanent resident varying in density between a minimum of 5.4/ha in October 2014 after a long drought and 68.8/ha in May 2015 following heavy rain.
- (8)
- The mean sex ratio over three years was 84.1% female, varying between extremes of 100% female in the driest periods, up to six months duration, down to 72.3% female in May 2015. The mean sex ratio at Kitengela is the highest recorded for D. chrysippus anywhere and exceeds the long-term (30-year) average of 74.5% for the Nairobi region which includes places such as Athi Plains and Kasarani.
- (9)
- The Spiroplasma infection rate of females at Kitengela, assessed by PCR, was 87.9% (n = 72), an estimate that tallies well with sex ratios and male-killing frequencies throughout the Nairobi region and over the years.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, D.A.S.; Lushai, G.; Allen, J.A. A classification of Danaus butterflies (Lepidoptera: Nymphalidae) based upon data from morphology and DNA. Zool. J. Linn. Soc. 2005, 144, 191–212. [Google Scholar] [CrossRef]
- Poulton, E.B. Essays on Evolution 1889–1907; Clarendon Press: Oxford, UK, 1908. [Google Scholar]
- Larson, T.B. Il y a trios millénaires et demi que Danaus chrysippus (L.) est connu en haute-Egypte (Lepidoptera: Danaidae). Linn. Belg. 1977, 7, 55–58. [Google Scholar]
- Ackery, P.R.; Vane-Wright, R.I. Milkweed Butterflies: Their Cladistics and Biology; British Museum (Natural History): London, UK, 1984. [Google Scholar]
- Lushai, G.; Zalucki, M.P.; Smith, D.A.S.; Goulson, D.; Daniels, G. The lesser wanderer butterfly, Danaus petilia (Stoll 1790) stat. rev. (Lepidoptera: Danainae), reinstated as a species. Aust. J. Entomol. 2005, 44, 6–14. [Google Scholar] [CrossRef]
- Braby, M.F.; Farias-Quipildor, G.E.; Vane-Wright, R.I.; Lohman, D.J. Morphological and molecular evidence supports the recognition of Danaus petilia (Stoll, 1790) (Lepidoptera: Nymphalidae) as a species distinct from Danaus chrysippus (Linnaeus, 1758). Syst. Biodivers. 2015, 13, 386–402. [Google Scholar] [CrossRef]
- Moore, F. A monograph of the Limnaina and Euploina, two groups of diurnal Lepidoptera belonging to the subfamily Euploeinae, with descriptions of new genera and species. Proc. Zool. Soc. Lond. 1883, 201–324. [Google Scholar] [CrossRef]
- Smith, D.A.S.; Gordon, I.J.; Traut, W.; Herren, J.; Collins, S.; Martins, D.J.; Saitoti, K.; Ireri, P.; Ffrench-Constant, R. A neo-W chromosome in a tropical butterfly links colour pattern, male-killing and speciation. Proc. R. Soc. B 2016, 283, 20160821. [Google Scholar] [CrossRef]
- Smith, D.A.S. African Queens and Their Kin: A Darwinian Odyssey; Brambleby Books: Taunton, UK, 2014. [Google Scholar]
- Owen, D.F.; Chanter, D.O. Population biology of tropical African butterflies. 2. Sex ratio and polymorphism in Danaus chrysippus L. Rev. Zool. Bot. Afr. 1968, 78, 81–97. [Google Scholar]
- Smith, D.A.S. Genetics of some polymorphic forms of the African butterfly Danaus chrysippus L. (Lepidoptera: Danaidae). Insect Syst. Evol. 1975, 6, 134–144. [Google Scholar] [CrossRef]
- Ford, E.B. Polymorphism and taxonomy. In The New Systematics; Huxley, J., Ed.; Clarendon Press: Oxford, UK, 1940; pp. 493–513. [Google Scholar]
- Smith, D.A.S.; Owen, D.F.; Gordon, I.J.; Lowis, N.K. The butterfly Danaus chrysippus (L.) in East Africa: Polymorphism and morph-ratio clines within a complex, extensive and dynamic hybrid zone. Zool. J. Linn. Soc. 1997, 120, 51–78. [Google Scholar] [CrossRef]
- Smith, D.A.S.; Gordon, I.J.; Depew, L.A.; Owen, D.F. Genetics of the butterfly Danaus chrysippus (L.) in a broad hybrid zone, with special reference to sex ratio, polymorphism and intragenomic conflict. Biol. J. Linn. Soc. 1998, 65, 1–40. [Google Scholar] [CrossRef]
- Bates, H.W. Contributions to an insect fauna of the Amazon Valley. Lepidoptera-Nymphalinae. Trans. Linn. Soc. Lond. 1862, 23, 495–566. [Google Scholar] [CrossRef]
- Müller, F. Notes on Brazilian Entomology. Trans. Entomol. Soc. Lond. 1878, 26, 211–233. [Google Scholar] [CrossRef]
- Swailem, S.M.; Ishmail, I.I. Biological studies on Danaus chrysippus L. (Lepidoptera: Danaidae). Bull. Société Entomol. d’Egypte 1971, 55, 211–218. [Google Scholar]
- Mebs, D.; Reuss, E.; Schneider, M. Studies on the cardenolide sequestration in African milkweed butterflies (Danaidae). Toxicon 2005, 45, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Brower, L.P. Ecological chemistry. Sci. Am. 1969, 220, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.; Kellet, D.N. Reactions of various predators to insects storing heart poisons (cardiac glycosides) in their tissues. J. Entomol. A 1972, 46, 103–110. [Google Scholar] [CrossRef]
- Dobler, S.; Dalla, S.; Wagschal, V.; Agrawal, A.A. Community-wide convergent evolution in insect adaptation to toxic cardenolides by substitutions in the Na, K-ATPase. Proc. Natl. Acad. Sci. USA 2012, 109, 13040–13045. [Google Scholar] [CrossRef]
- Aardema, M.L.; Zhen, Y.; Andolfatto, P. The evolution of cardenolide-resistant forms of NA+, K+-ATPase in Danainae butterflies. Mol. Ecol. 2011, 21, 340–349. [Google Scholar] [CrossRef]
- Rothschild, M.; von Euw, J.; Reichstein, J.; Smith, D.A.S.; Pierre, J. Cardenolide storage in Danaus chrysippus with additional notes on D. plexippus. Proc. R. Soc. B 1975, 190, 1–31. [Google Scholar]
- Bernays, E.A.; Singer, M.S.; Rodrigues, D. Trenching behaviour by caterpillars of the Euphorbia specialist, Pygarctia roseicapitis: A field study. J. Insect Behav. 2004, 17, 41–52. [Google Scholar] [CrossRef]
- Singer, M.S.; Stireman, J.O., III. Does anti-parasitoid defense explain host-plant selection by a polyphagous caterpillar? Oikos 2003, 100, 554–562. [Google Scholar] [CrossRef]
- Singer, M.S.; Mace, K.C.; Bernays, E.A. Self-medication as adaptive plasticity: Increased ingestion of plant toxins by parasitized caterpillars. PLoS ONE 2009, 4, e4796. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.; Reichstein, R.T.; Von Euw, J.; Apin, R.T.; Harmann, R.R.M. Toxic Lepidoptera. Toxicon 1970, 8, 293–299. [Google Scholar] [CrossRef]
- Pliske, T.E. Attraction of Lepidoptera to plants containing pyrrolizidine alkaloids. Environ. Entomol. 1975, 4, 474–479. [Google Scholar] [CrossRef]
- Owen, D.F. Tropical Butterflies; Clarendon Press: Oxford, UK, 1971. [Google Scholar]
- Edgar, J.A.; Culvenor, C.C.J. Pyrrolizidine ester alkaloid in danaid butterflies. Nature 1974, 248, 614–616. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.S., Jr. Adult-obtained pyrrolizidine alkaloids defend ithomiine butterflies against a spider predator. Nature 1984, 309, 707–709. [Google Scholar] [CrossRef]
- Brown, K.S., Jr. Chemical ecology of dehydropyrrolizidine alkaloids in adult Ithomiinae (Lepidoptera: Nymphalidae). Rev. Bras. Biol. 1984, 44, 435–460. [Google Scholar]
- Edgar, J.A.; Culvenor, C.C.J.; Smith, L.W. Dihydropyrrolizidine derivatives in the hair pencil secretion of danaid butterflies. Experientia 1971, 27, 761–762. [Google Scholar] [CrossRef]
- Schulz, S.; Boppré, M.; Vane-Wright, R.I. Specific mixtures of secretions from male scent organs of African milkweed butterflies (Danainae). Philos. Trans. R. Soc. B 1993, 342, 161–181. [Google Scholar]
- Edgar, J.A.; Boppré, M.; Schneider, D. Pyrrolizidine alkaloid storage in African and Australian danaid butterflies. Experientia 1979, 35, 1447–1448. [Google Scholar] [CrossRef]
- Dussourd, D.E.; Harvis, C.A.; Meinwald, J.; Eisner, T. Paternal allocation of sequestered plant pyrrolizidine alkaloid to eggs in the danaine butterfly Danaus gilippus. Cell. Mol. Life Sci. 1989, 45, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Eisner, T.; Eisner, M. Unpalatability of the pyrrolizidine-containing moth Utetheisa ornatrix and its larva to wolf spiders. Psyche 1991, 98, 111–118. [Google Scholar] [CrossRef]
- Bull, L.B.; Culvenor, C.C.G.; Dick, A.T. The Pyrrolizidine Alkaloids, Their Chemistry, Pathogenicity and Other Biological Properties; Frontiers of Biology: Amsterdam, The Netherlands, 1968. [Google Scholar]
- Bate-Smith, E.C. Attractants and repellents in higher animals. In Phytochemical Ecology; Harborne, J.B., Ed.; Academic Press: London, UK, 1972; pp. 45–56. [Google Scholar]
- Kassarov, L. Are birds able to taste and reject butterflies based on ‘beak mark tasting’? A different point of view. Behaviour 1999, 136, 965–981. [Google Scholar] [CrossRef]
- Birkhead, T. Bird Sense: What It’s Like to Be a Bird; Bloomsbury: London, UK, 2012. [Google Scholar]
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme. Proc. R. Soc. B 1979, 205, 581–598. [Google Scholar]
- Koonin, E.V. Splendor and misery of adaptation, or the importance of neutral null for understanding evolution. BMC Biol. 2016, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Boppré, M. Leaf scratching—A specialized behaviour of danaine butterflies for gathering secondary plant substances. Oecologia 1983, 59, 414–416. [Google Scholar] [CrossRef]
- Seibt, U.; Schneider, D.; Eisner, T. Duftpinsel, Flügeltaschen und Balz das Tagfalters Danaus chrysippus. Beih. Z. Tierppsychol. 1972, 31, 513–530. [Google Scholar] [CrossRef]
- Brower, L.P.; Brower, J.V.Z.; Cranston, F.P. Courtship behaviour of the queen butterfly Danaus gilippus berenice. Zool. N. Y. 1965, 50, 1–39. [Google Scholar]
- Smith, D.A.S.; Gordon, I.J.; Lushai, G.; Goulson, D.; Allen, J.A.; Maclean, N. Hybrid queen butterflies from a cross Danaus chrysippus (L.) × D. gilippus (Cramer): Confirmation of species status for the parents and further support for Haldane’s Rule. Biol. J. Linn. Soc. 2002, 76, 535–544. [Google Scholar] [CrossRef]
- Edgar, J.A.; Cockrum, C.C.J.; Frahn, J.L. Pyrrolizidine alkaloids in Danaus plexippus L. and Danaus chrysippus L. Experientia 1976, 32, 1535–1537. [Google Scholar] [CrossRef]
- Sakai, H.; Oshima, H.; Yuri, H.; Gotoh, H.; Daimon, T.; Yaginuma, T.; Sahara, K.; Niimi, T. Dimorphic sperm formation by Sex-lethal. Proc. Natl. Acad. Sci. USA 2019, 116, 10412–10417. [Google Scholar] [CrossRef]
- Trimen, R. On some remarkable mimetic analogies among African butterflies. Trans. Linn. Soc. 1869, 26, 497–522. [Google Scholar] [CrossRef]
- Wallace, A.R. The Malay Archipelago: The Land of the Orang-Utan and the Bird of Paradise. A Narrative of Travel with Studies of Man and Nature; 2 vols; Macmillan: London, UK, 1869. [Google Scholar]
- Darwin, C. The Descent of Man and Selection in Relation to Sex, 2nd ed.; John Murray: London, UK, 1874. [Google Scholar]
- Swynnerton, C.F.M. Experiments and observations bearing on the explanation of form and colouring, 1908–1913. J. Linn. Soc. Zool. 1919, 33, 203–385. [Google Scholar] [CrossRef]
- Parsons, J.A. A digitalis-like toxin in the monarch butterfly, Danaus plexippus. J. Physiol. 1965, 178, 290–303. [Google Scholar] [CrossRef]
- Poulton, E.B. The Colours of Animals: Their Meaning and Use, Especially Considered in the Case of Insects; Kegan Paul: London, UK, 1890. [Google Scholar]
- Brower, L.P.; Brower, J.V.Z. Birds, butterflies and plant poisons: A study in ecological chemistry. Zool. N.Y. 1964, 49, 137–149. [Google Scholar]
- Carpenter, G.D.H. The relative frequency of beak marks on butterflies of different edibility to birds. Proc. Zool. Soc. Lond. 1941, 111, 223–231. [Google Scholar] [CrossRef]
- Smith, D.A.S. The significance of beak marks on the wings of an aposematic and distasteful butterfly. Nature 1979, 281, 215–216. [Google Scholar] [CrossRef]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar]
- Smith, D.A.S. Mate selection in butterflies: Competition, coyness, choice and chauvinism. In The Biology of Butterflies; Vane-Wright, R.I., Ackery, P.R., Eds.; Academic Press: London, UK, 1984; pp. 225–244. [Google Scholar]
- Owen, D.F.; Smith, D.A.S. Danaus chrysippus and its polymorphic Müllerian mimics in tropical Africa. Trop. Lepid. 1993, 4, 77–81. [Google Scholar]
- Smith, D.A.S. Batesian mimicry between Danaus chrysippus and Hypolimnas misippus (Lepidoptera) in Tanzania. Nature 1973, 242, 129–131. [Google Scholar] [CrossRef]
- Smith, D.A.S. Phenotypic diversity, mimicry and natural selection in the African butterfly Hypolimnas misippus L. (Lepidoptera: Nymphalidae). Biol. J. Linn. Soc. 1976, 8, 183–204. [Google Scholar] [CrossRef]
- Gordon, I.J. Natural selection for rare and mimetic colour pattern combinations in wild populations of the diadem butterfly Hypolimnas misippus (L.). Biol. J. Linn. Soc. 1987, 31, 1–23. [Google Scholar] [CrossRef]
- Gordon, I.J.; Smith, D.A.S. Diversity in mimicry. Trends Ecol. Evol. 1999, 13, 150–151. [Google Scholar] [CrossRef]
- Gordon, I.J.; Edmunds, M.; Edgar, J.A.; Lawrence, J.; Smith, D.A.S. Linkage disequilibrium and natural selection for mimicry in the Batesian mimic Hypolimnas misippus (L.) in the Afrotropics. Biol. J. Linn. Soc. 2010, 100, 180–194. [Google Scholar] [CrossRef]
- Poulton, P.B. Danaida chrysippus L. and D. dorippus Klug, proved by breeding to be two forms of the same species. Proc. Entomol. Soc. Lond. 1924, 1925, cxix–cxxiii. [Google Scholar]
- Gordon, I.J. Polymorphism of the tropical butterfly Danaus chrysippus L. in Africa. Heredity 1984, 53, 583–593. [Google Scholar] [CrossRef]
- Clarke, C.A.; Sheppard, P.M.; Smith, A.G. The genetics of fore and hindwing colour in crosses between Danaus chrysippus from Australia and Sierra Leone (Danaidae). J. Lepid. Soc. 1973, 27, 73–77. [Google Scholar]
- Sturtevant, A.H. Linkage in the silkworm moth. Am. Nat. 1915, 28, 315–317. [Google Scholar] [CrossRef]
- Suomalainen, E.; Cook, L.M.; Turner, J.R.G. Achiasmate oogenesis in the Heliconiine butterflies. Hereditas 1973, 74, 302–304. [Google Scholar] [CrossRef]
- Turner, J.R.G.; Sheppard, P.M. Absence of crossing-over in female butterflies (Heliconius). Heredity 1975, 34, 265–269. [Google Scholar] [CrossRef]
- Martin, S.H.; Singh, K.S.; Gordon, I.J.; Omufwoko, K.S.; Collins, S.; Warren, I.A.; Munby, H.; Brattström, O.; Traut, W.; Martins, D.J.; et al. Whole-chromosome hitchhiking driven by a male-killing endosymbiont. BioRxiv 2019. [Google Scholar] [CrossRef]
- Hurst, G.G.D.; Hurst, L.D.; Majerus, M. Cytoplasmic sex ratio distorters. In Influential Passengers; O’Neil, S.L., Hoffman, A.A., Werren, J.H., Eds.; Oxford University Press: Oxford, UK, 1997; pp. 125–154. [Google Scholar]
- Hamilton, W.D. Extraordinary sex ratios. Science 1967, 156, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.S. All-female broods in Danaus chrysippus L. and their ecological significance. Heredity 1975, 34, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M.; Hurst, G.D.D.; Jiggins, C.D.; Schulenburg, J.H.G.V.D.; Majerus, M.E.N. The butterfly Danaus chrysippus is infected by a male-killing Spiroplasma bacterium. Parasitology 2000, 120, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Herren, J.; Gordon, I.J.; Holland, P.W.H.; Smith, D.A.S. The butterfly Danaus chrysippus (L.) in Kenya is variably infected with respect to genotype and body size by a maternally transmitted male-killing endosymbiont (Spiroplasma). Int. J. Trop. Insect Sci. 2007, 27, 62–69. [Google Scholar] [CrossRef]
- Hassan, S.S.M.; Idris, E.; Majerus, M.E.N. Male-killer dynamics in Danaus chrysippus (L.) (Lepidoptera: Nymphalidae) in East Africa. Afr. J. Ecol. 2012, 50, 489–499. [Google Scholar] [CrossRef]
- Smith, D.A.S.; Gordon, I.J.; Allen, J.A. Reinforcement in hybrids among once isolated semispecies of Danaus chrysippus (L.) and evidence for sex chromosome evolution. Ecol. Entomol. 2010, 35, 77–89. [Google Scholar] [CrossRef]
- Smith, D.A.S. Heterosis, epistasis and linkage disequilibrium in a wild population of the polymorphic butterfly Danaus chrysippus L. Zool. J. Linn. Soc. 1980, 69, 87–109. [Google Scholar] [CrossRef]
- Gordon, I.J.; Ireri, P.; Smith, D.A.S. Hologenomic speciation: Synergy between a male-killing bacterium and sex-linkage creates a ‘magic trait’ in a butterfly hybrid zone. Biol. J. Linn. Soc. 2014, 100, 180–194. [Google Scholar] [CrossRef]
- Traut, W.; Ahola, V.; Smith, D.A.S.; Gordon, I.J.; Ffrench-Constant, R.H. Karyotypes versus Genomes: The Nymphalid Butterflies Melitaea cinxia, Danaus plexippus and D. chrysippus. Cytogenet. Genome Res. 2017, 153, 46–53. [Google Scholar] [CrossRef]
- White, M.J.D. Modes of Speciation; Freeman & Co.: San Francisco, CA, USA, 1978. [Google Scholar]
- Gazarve, E.; Catalan, J.; da Graca Ramalino, M.; da Luz Mathias, M.; Nunes, A.C.; Brittan-Davidian, J.; Auffray, J.C. Do chromosomal hybrids necessarily suffer from developmental instability? Biol. J. Linn. Soc. 2006, 88, 33–43. [Google Scholar] [CrossRef]
- Smith, D.A.S. Negative non-random mating in the polymorphic butterfly Danaus chrysippus in Tanzania. Nature 1973, 242, 131–132. [Google Scholar] [CrossRef]
- Gupta, Y. Chromosome studies of some Indian Lepidoptera. Chromosoma 1964, 15, 540–561. [Google Scholar] [CrossRef]
- De Lesse, H.; Condamin, M. Formules chromosomiques de quelques Lépidoptères Rhopalocères du Sénégal. Bull. l’Institut Fondam. d’Afrique Noire Dakar 1962, 24, 464–473. [Google Scholar]
- Servedio, M.R.; Van Doorn, G.S.; Kopp, M.; Frame, A.M.; Nosil, P. Magic traits in speciation: ‘magic’ but not rare? Trends Ecol. Evol. 2011, 26, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Haller, B.C.; Luis, F.; Rolshausen, G.; Gotanda, K.M.; Hendry, A.P. Magic traits: Distinguishing the important from the trivial. Trends Ecol. Evol. 2013, 27, 4–5. [Google Scholar] [CrossRef]
- Williams, C.B. Insect Migration; Oliver and Boyd: Edinburgh, UK, 1958. [Google Scholar]
- Smith, D.A.S.; Owen, D.F. Colour genes as markers for migratory activity: The butterfly Danaus chrysippus (L.) in Africa. Oikos 1997, 78, 127–135. [Google Scholar] [CrossRef]
- Lushai, G.; Smith, D.A.S.; Gordon, I.J.; Goulson, D.; Allen, J.A.; Maclean, N. Incomplete sexual isolation in sympatry between subspecies of the butterfly Danaus chrysippus (L.) and the creation of a hybrid zone. Heredity 2003, 90, 236–246. [Google Scholar] [CrossRef]
- Gordon, I.J.; Ireri, P.; Smith, D.A.S. Preference for isolated plants facilitates invasion of Danaus chrysippus (Linnaeus, 1758) (Lepidoptera: Nymphalidae) by a bacterial male-killer Spiroplasma. Austral Entomol. 2014, 54, 210–216. [Google Scholar] [CrossRef]
- Majerus, M.E.N. Sex Wars; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Zalucki, M.P. Sex around the milkweed patch. In The Biology and Conservation of the Monarch Butterfly; Malcolm, S.B., Zalucki, M.P., Eds.; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1993; pp. 69–76. [Google Scholar]
- Cane, M.A.; Molnar, P. Closing of the Indonesian seaway as a precursor to east African aridification around 3–4 million years ago. Nature 2001, 411, 157–162. [Google Scholar] [CrossRef]
- Fer, I.; Tietjen, B.; Jeltsch, F.; Trauth, M.H. Modelling vegetation change during Late Cenozoic uplift of the East African plateau. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 467, 120–130. [Google Scholar] [CrossRef]
- Vrba, E.S. African Bovidae: Evolutionary events since the Miocene. S. Afr. J. Sci. 1985, 81, 263–266. [Google Scholar]
- Leakey, M.G.; Spoor, F.; Brown, F.H.; Gathogo, P.N.; Kiairie, C.; Leakey, L.N.; McDougall, I. A new hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 2001, 410, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.E. The Bird Faunas of Africa and Its Islands; Academic Press: London, UK, 1966. [Google Scholar]
- Dartnell, L. Origins: How the Earth Made Us; The Bodley Head: London, UK, 2019. [Google Scholar]
- Maslin, M.A.; Brierley, C.M.; Milner, A.M.; Shultz, S.; Trauth, M.H.; Wilson, K.E. East African climate pulses and early human evolution. Quat. Sci. Rev. 2014, 101, 1–17. [Google Scholar] [CrossRef]
- Jung, G.; Prange, M.; Schulz, M. Influence of topography on tropical African vegetation coverage. Clim. Dyn. 2016, 46, 2535–2549. [Google Scholar] [CrossRef]
- Moreau, R.E. Vicissitudes of the African biomes in the Late Pleistocene. Proc. Zool. Soc. Lond. 1963, 141, 395–421. [Google Scholar] [CrossRef]
- Williams, M.A.J.; Faure, H. The Sahara and The Nile: Quaternary Environments and Prehistoric Occupation in Northern Africa; A.A. Balkema: Rotterdam, The Netherlands, 1980; pp. 207–224. [Google Scholar]
- Lancaster, N. Evidence for a widespread late Pleistocene humid period in the Kalahari. Nature 1979, 279, 145–146. [Google Scholar] [CrossRef]
- Alin, S.R.; Cohen, A.S. Lake level history of Lake Tanganyika, East Africa, for the past 2500 years based on Ostracod inferred water depth reconstruction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 199, 31–49. [Google Scholar] [CrossRef]
- Flenley, J.R. The Equatorial Rain Forest: A Geological History; Butterworths: London, UK, 1979. [Google Scholar]
- Maslin, M.A.; Christensen, B. Tectonics, orbital forcing, global climate change and human evolution in Africa: Introduction to the African paleoclimate special volume. J. Hum. Evol. 2007, 53, 443–464. [Google Scholar] [CrossRef]
- Trauth, M.H.; Maslin, M.A.; Deino, A.L.; Junginger, A.; Lesoloyia, E.O.; Odada, D.O.; Olaka, L.A.; Strecker, M.R.; Tiedermann, R. Human evolution in a variable environment: The amplifier Lakes of East Africa. Quat. Sci. Rev. 2010, 29, 2981–2988. [Google Scholar] [CrossRef]
- DeMenocal, P.B.; Ortiz, J.; Guilderson, T.; Adkins, J.; Sarntheim, M.; Baker, L.; Yarusinsky, M. Abrupt onset and termination of the African Humid Period: Rapid climate responses to gradual insolation forcing. Quat. Sci. Rev. 2000, 19, 347–361. [Google Scholar] [CrossRef]
- Gasse, F. Hydrological changes in the African tropics since the last glacial maximum. Quat. Sci. Rev. 2000, 19, 189–211. [Google Scholar] [CrossRef]
- Russell, J.M.; Johnson, T.C. A high-resolution geochemical record from Lake Edward, Uganda-Congo, and the timing and causes of tropical African drought during the late Holocene. Quat. Sci. Rev. 2005, 24, 1375–1389. [Google Scholar] [CrossRef]
- Russell, J.M.; Johnson, T.C. Little Ice Age drought in Equatorial Africa: ITCZ migrations and ENSO variability. Geology 2007, 35, 21–24. [Google Scholar] [CrossRef]
- Smith, D.A.S.; Owen, D.F.; Gordon, I.J.; Owiny, A.M. Polymorphism and evolution in the butterfly Danaus chrysippus (L.) in Africa. Heredity 1993, 71, 127–135. [Google Scholar] [CrossRef]
- Haldane, J.B.S. Sex ratio and unisexual sterility in hybrid animals. J. Genet. 1922, 12, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.A.S. Evidence for autosomal meiotic drive in the butterfly Danaus chrysippus L. Heredity 1976, 36, 139–142. [Google Scholar] [CrossRef]
- Idris, E.; Hassan, S.S.M. The Queen Butterfly, Danaus chrysippus (L.) (Lepidoptera: Nymphalidae) at Khartoum, Sudan. Egypt. Acad. J. Biol. Sci. 2012, 5, 95–102. [Google Scholar] [CrossRef]
- Charlat, S.; Hornett, E.A.; Fullard, J.H.; Davies, N.; Roderick, G.K.; Wedell, N.; Hurst, G.D.D. Extraordinary flux in sex ratio. Science 2007, 317, 214. [Google Scholar] [CrossRef]
- Hornett, E.A.; Moran, B.; Reynolds, L.A.; Charlat, S.; Tazzyman, S.; Wedell, N.; Jiggins, C.D.; Hurst, G.D.D. The Evolution of Sex Ratio Distorter Suppression Affects a 25cM Genomic Region in the Butterfly Hypolimnas bolina. PLoS Genet. 2014, 10, e1004822. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies & Forms | Original Status & Authority | Present Status & Range |
|---|---|---|
| chrysippus | Papilio chrysippus, L. 1758 | ssp, S. Europe, N. Africa, Asia |
| alcippus | Papilio alcippus, Cramer 1777 | ssp, W. Africa |
| dorippus | Euploea dorippus, Klug 1845 | ssp, NE Africa, Arabia |
| orientis | f. of Danaida chrysippus, Aurivillius 1909 | ssp, S. Africa, Indian Ocean |
| klugii | Limnas klugii, Butler 1886 | orientis × dorippus F1 hybrid |
| infumata | f. of Danaida chrysippus, Aurivillius 1898 | chrysippus × orientis F1 hybrid |
| transiens | f. of Danaida dorippus, Suffert 1900 | chrysippus × dorippus F1 hybrid |
| alcippoides 1 | f. of Danaus chrysippus, Moore 1883 | chrysippus or orientis × alcippus F1 hybrid |
| albinus | f. of Danaida dorippus, Lanz 1896 | alcippus × dorippus F2 hybrid |
| semialbinus | f. of Danaida dorippus, Strand 1910 | alcippus × dorippus F1 hybrid |
| Genotype | AA | Aa | aa |
|---|---|---|---|
| bc/bc | chrysippus | orange alcippoides orange alcippus | orange alcippus |
| Bc/Bc | orientis | brown alcippoides brown alcippus | brown alcippus |
| bC/bC | dorippus | semialbinus albinus | albinus |
| bC/bc | transiens | semialbinus-transiens albinus-transiens | albinus-transiens |
| Bc/bC | klugii | semialbinus-klugii albinus-klugii | albinus-klugii |
| Bc/bc | infumata | alcippoides-infumata alcippus-infumata | alcippus-infumata |
| Study Areas | Spiroplasma | ||
|---|---|---|---|
| + | − | n | |
| East Africa | 30 1 | 0 | 30 |
| Elsewhere | 2 2 | 12 3 | 14 |
| Totals | 32 | 12 | 44 |
| Sex | Genotype Frequencies | |||
|---|---|---|---|---|
| CC | Cc | cc | n | |
| Females | 0 | 801 | 118 | 919 |
| Males | 130 | 37 | 7 | 174 |
| Totals | 130 | 838 | 125 | 1093 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, D.A.S.; Traut, W.; Martin, S.H.; Ireri, P.; Omufwoko, K.S.; ffrench-Constant, R.; Gordon, I.J. Neo Sex Chromosomes, Colour Polymorphism and Male-Killing in the African Queen Butterfly, Danaus chrysippus (L.). Insects 2019, 10, 291. https://doi.org/10.3390/insects10090291
Smith DAS, Traut W, Martin SH, Ireri P, Omufwoko KS, ffrench-Constant R, Gordon IJ. Neo Sex Chromosomes, Colour Polymorphism and Male-Killing in the African Queen Butterfly, Danaus chrysippus (L.). Insects. 2019; 10(9):291. https://doi.org/10.3390/insects10090291
Chicago/Turabian StyleSmith, David A.S., Walther Traut, Simon H. Martin, Piera Ireri, Kennedy S. Omufwoko, Richard ffrench-Constant, and Ian J. Gordon. 2019. "Neo Sex Chromosomes, Colour Polymorphism and Male-Killing in the African Queen Butterfly, Danaus chrysippus (L.)" Insects 10, no. 9: 291. https://doi.org/10.3390/insects10090291