Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Strains
2.2. Mosquito Rearing
2.3. Effect of Fitness Cost on Biological Parameters
2.3.1. Developmental Time and Viability of Mosquitoes
2.3.2. Size of Mosquitoes
2.3.3. Blood Feeding Capacity
2.3.4. Fecundity
2.3.5. Egg Viability after Storage
2.3.6. Adult Longevity
2.3.7. Larval Competition
2.3.8. Colony Competition
2.4. Statistical Analysis
3. Results
3.1. Development and Fecundity
3.2. Blood Feeding Capability
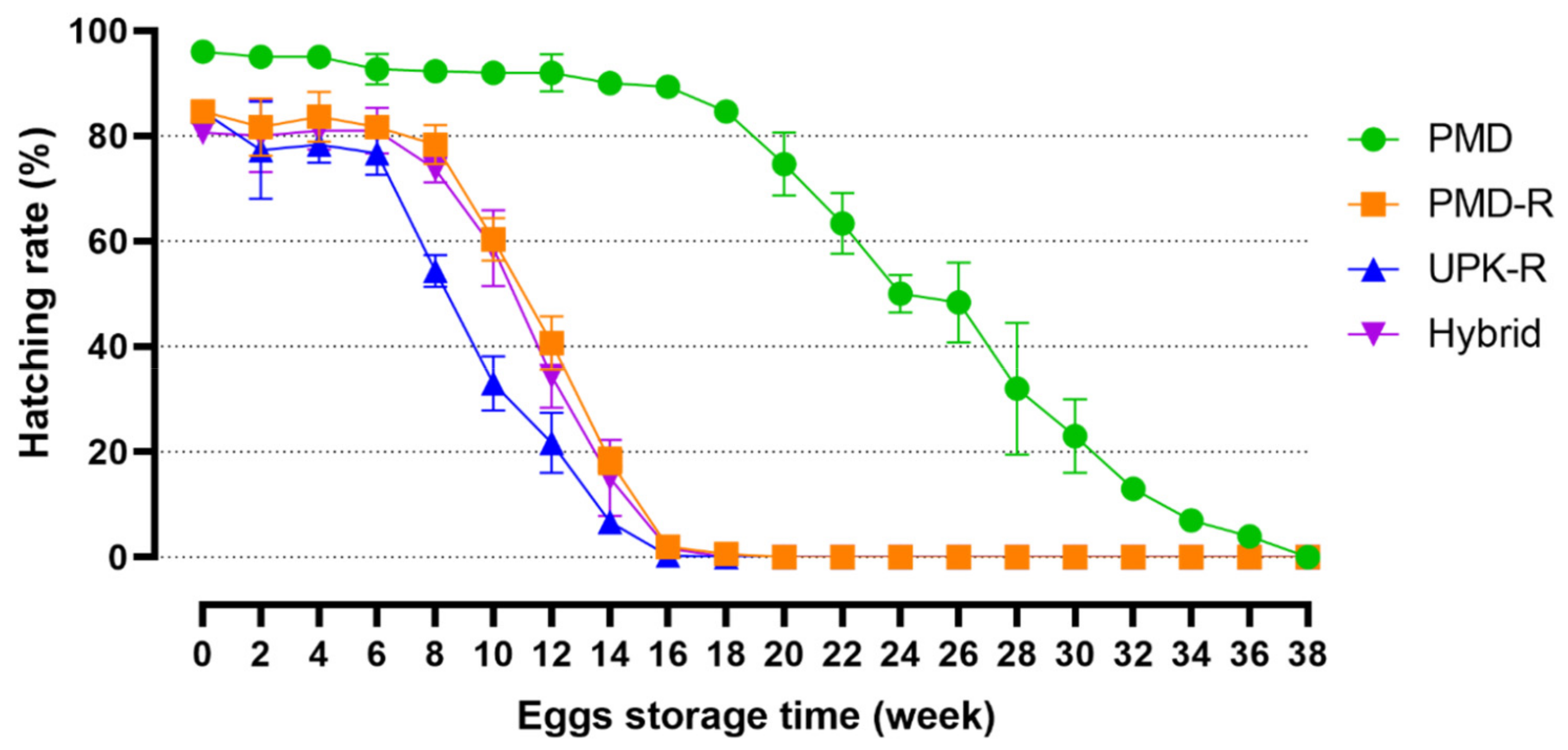
3.3. Viability of Eggs after Storage
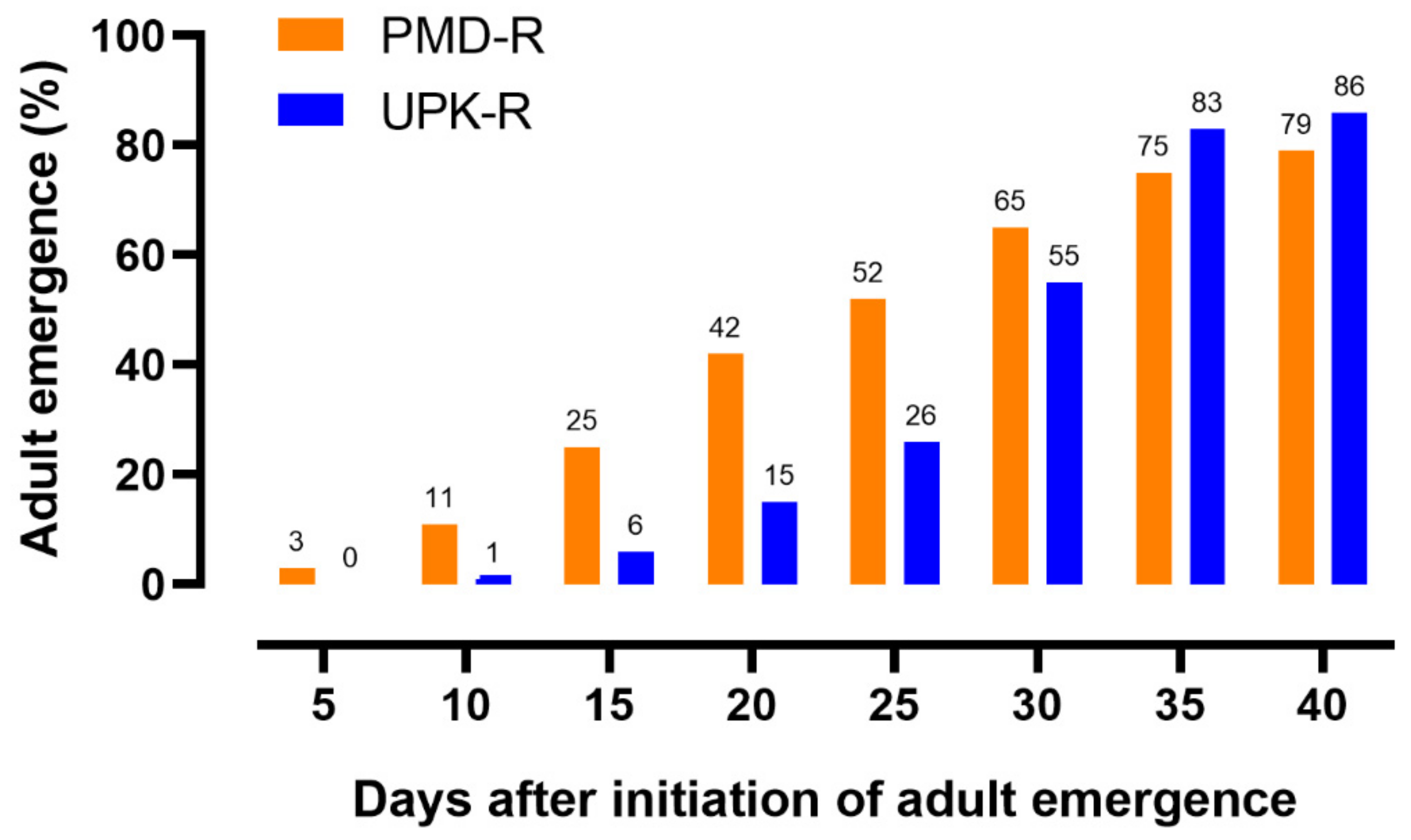
3.4. Adult Longevity
3.5. PMD-R and UPK-R Competition
3.5.1. Larval Competition
3.5.2. Colony Competition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Monitoring and Managing Insecticide Resistance in Aedes Aegypti Populations, Interim Guidance for Entomologist; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- World Health Organization. Zika Virus. 2016. Available online: http://www.who.int/mediacentre/factsheets/zika/en/ (accessed on 20 July 2018).
- World Health Organization. Dengue and Severe Dengue. 2017. Available online: http://www.who.int/mediacentre/factsheets/fs117/en/ (accessed on 15 April 2019).
- Ministry of Public Health. Morbidity and Mortality Weekly Report, Week No. 52nd. 2019. Available online: https://ddc.moph.go.th/th/site/newsview/view/696 (accessed on 8 July 2019).
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Kidwell, M.G.; Ribeiro, J.M. Can transposable elements be used to drive disease refractoriness genes into vector populations? Parasitol. Today 1992, 8, 325–329. [Google Scholar] [CrossRef]
- Speranca, M.A.; Capurro, M.L. Perspectives in the control of infectious diseases by transgenic mosquitoes in the post-genomic era: A review. Mem. Inst. Oswaldo Cruz 2007, 102, 425–433. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Strategy for Dengue Prevention and Control 2012–2020; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Chareonviriyaphap, T.; Bangs, M.J.; Suwonkerd, W.; Kongmee, M.; Corbel, V.; Ngoen-Klan, R. Review of insecticide resistance and behavioral avoidance of vectors of human diseases in Thailand. Parasit. Vectors 2013, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, H.; Zaim, M.; Yadav, R.S.; Soares, A.; Ameneshewa, B.; Mnzava, A.; Hii, J.; Dash, A.P.; Ejov, M. Global trends in the use of insecticides to control vector-borne diseases. Environ. Health Perspect. 2012, 120, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.G.; Field, L.M.; Usherwood, P.N.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neely, J.M. Insecticide-resistance studies on Aedes aegypti in Thailand. Bull. World Health Organ. 1966, 35, 91. [Google Scholar] [PubMed]
- Somboon, P.; Prapanthadara, L.A.; Suwonkerd, W. Insecticide susceptibility tests of Anopheles minimus s.l., Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus in northern Thailand. Southeast Asian J. Trop. Med. Public Health 2003, 34, 87–93. [Google Scholar] [PubMed]
- Jirakanjanakit, N.; Rongnoparut, P.; Saengtharatip, S.; Chareonviriyaphap, T.; Duchon, S.; Bellec, C.; Yoksan, S. Insecticide susceptible/resistance status in Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Thailand during 2003–2005. J. Econ. Entomol. 2007, 100, 545–550. [Google Scholar] [PubMed]
- Paeporn, P.; Ya-umphan, P.; Supaphathom, K.; Savanpanyalert, P.; Wattanachai, P.; Patimaprakorn, R. Insecticide susceptibility and selection for resistance in a population of Aedes aegypti from Ratchaburi province, Thailand. Trop Biomed 2004, 21, 1–6. [Google Scholar] [PubMed]
- Pethuan, S.; Jirakanjanakit, N.; Saengtharatip, S.; Chareonviriyaphap, T.; Kaewpa, D.; Rongnoparut, P. Biochemical studies of insecticide resistance in Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Thailand. Trop Biomed 2007, 24, 7–15. [Google Scholar] [PubMed]
- Ponlawat, A.; Scott, J.G.; Harrington, L.C. Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J. Med. Entomol 2005, 42, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef] [PubMed]
- David, J.P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R Soc. B Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Molec. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M.; Knipple, D.C. The molecular biology of knockdown resistance to pyrethroid insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef]
- Kasai, S.; Komagata, O.; Itokawa, K.; Shono, T.; Ng, L.C.; Kobayashi, M.; Tomita, T. Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: Target site insensitivity, penetration, and metabolism. PLoS Negl. Trop. Dis. 2014, 8, e2948. [Google Scholar] [CrossRef]
- Wood, O.; Hanrahan, S.; Coetzee, M.; Koekemoer, L.; Brooke, B. Cuticle thickening associated with pyrethroid resistance in the major malaria vector Anopheles funestus. Parasit. Vectors 2010, 3, 67. [Google Scholar] [CrossRef]
- Stenhouse, S.A.; Plernsub, S.; Yanola, J.; Lumjuan, N.; Dantrakool, A.; Choochote, W.; Somboon, P. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasit. Vectors 2013, 6, 253. [Google Scholar] [CrossRef]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Prapanthadara, L.-A. A novel F1552/C1552 point mutation in the Aedes aegypti voltage-gated sodium channel gene associated with permethrin resistance. Pestic. Biochem. Phys. 2010, 96, 127–131. [Google Scholar] [CrossRef]
- Lumjuan, N.; Wicheer, J.; Leelapat, P.; Choochote, W.; Somboon, P. Identification and characterisation of Aedes aegypti aldehyde dehydrogenases involved in pyrethroid metabolism. PLoS ONE 2014, 9, e102746. [Google Scholar] [CrossRef] [PubMed]
- Plernsub, S.; Saingamsook, J.; Yanola, J.; Lumjuan, N.; Tippawangkosol, P.; Sukontason, K.; Walton, C.; Somboon, P. Additive effect of knockdown resistance mutations, S989P, V1016G and F1534C, in a heterozygous genotype conferring pyrethroid resistance in Aedes aegypti in Thailand. Parasit. Vectors 2016, 9, 417. [Google Scholar] [CrossRef] [PubMed]
- Prapanthadara, L.; Promtet, N.; Koottathep, S.; Somboon, P.; Suwonkerd, W.; McCarroll, L.; Hemingway, J. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull. 2002, 26, 185–189. [Google Scholar]
- Somwang, P.; Yanola, J.; Suwan, W.; Walton, C.; Lumjuan, N.; Prapanthadara, L.A.; Somboon, P. Enzymes-based resistant mechanism in pyrethroid resistant and susceptible Aedes aegypti strains from northern Thailand. Parasitol. Res. 2011, 109, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; McCarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003, 17, 87–94. [Google Scholar] [CrossRef]
- Srisawat, R.; Komalamisra, N.; Eshita, Y.; Zheng, M.; Ono, K.; Itoh, T.Q.; Matsumoto, A.; Petmitr, S.; Rongsriyam, Y. Point mutations in domain II of the voltage-gated sodium channel gene in deltamethrin-resistant Aedes aegypti (Diptera: Culicidae). Appl. Entomol. Zool. 2010, 45, 275–282. [Google Scholar] [CrossRef]
- Kawada, H.; Oo, S.Z.; Thaung, S.; Kawashima, E.; Maung, Y.N.; Thu, H.M.; Thant, K.Z.; Minakawa, N. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl. Trop. Dis. 2014, 8, e3032. [Google Scholar] [CrossRef]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L.A. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef]
- Harris, A.F.; Rajatileka, S.; Ranson, H. Pyrethroid resistance in Aedes aegypti from Grand Cayman. Am. J. Trop. Med. Hyg. 2010, 83, 277–284. [Google Scholar] [CrossRef]
- Linss, J.G.; Brito, L.P.; Garcia, G.A.; Araki, A.S.; Bruno, R.V.; Lima, J.B.; Valle, D.; Martins, A.J. Distribution and dissemination of the Val1016Ile and Phe1534Cys Kdr mutations in Aedes aegypti Brazilian natural populations. Parasit. Vectors 2014, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Plernsub, S.; Saingamsook, J.; Yanola, J.; Lumjuan, N.; Tippawangkosol, P.; Walton, C.; Somboon, P. Temporal frequency of knockdown resistance mutations, F1534C and V1016G, in Aedes aegypti in Chiang Mai city, Thailand and the impact of the mutations on the efficiency of thermal fogging spray with pyrethroids. Acta Trop. 2016, 162, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Belinato, T.A.; Martins, A.J.; Valle, D. Fitness evaluation of two Brazilian Aedes aegypti field populations with distinct levels of resistance to the organophosphate temephos. Mem. Inst. Oswaldo Cruz 2012, 107, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.P.; Linss, J.G.; Lima-Camara, T.N.; Belinato, T.A.; Peixoto, A.A.; Lima, J.B.; Valle, D.; Martins, A.J. Assessing the effects of Aedes aegypti kdr mutations on pyrethroid resistance and its fitness cost. PLoS ONE 2013, 8, e60878. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.F.; de Melo-Santos, M.A.; Santos, E.M.; Beserra, E.B.; Helvecio, E.; de Carvalho-Leandro, D.; dos Santos, B.S.; de Menezes Lima, V.L.; Ayres, C.F. Fitness cost in field and laboratory Aedes aegypti populations associated with resistance to the insecticide temephos. Parasit. Vectors 2015, 8, 662. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Thomas, A.; Samuel, T.; Sahgal, A.; Verma, A.; Pillai, M.K. Diminished reproductive fitness associated with the deltamethrin resistance in an Indian strain of dengue vector mosquito, Aedes aegypti L. Trop Biomed 2009, 26, 155–164. [Google Scholar] [PubMed]
- Martins, A.J.; Ribeiro, C.D.; Bellinato, D.F.; Peixoto, A.A.; Valle, D.; Lima, J.B. Effect of insecticide resistance on development, longevity and reproduction of field or laboratory selected Aedes aegypti populations. PLoS ONE 2012, 7, e31889. [Google Scholar] [CrossRef]
- Mebrahtu, Y.B.; Norem, J.; Taylor, M. Inheritance of larval resistance to permethrin in Aedes aegypti and association with sex ratio distortion and life history variation. Am. J. Trop. Med. Hyg. 1997, 56, 456–465. [Google Scholar] [CrossRef]
- Amin, A.M.; White, G.B. Relative fitness of organophosphate-resistant and susceptible strains of Culex quinquefasciatus Say (Diptera: Culicidae). Bull. Entomol. Res. 2009, 74, 591–598. [Google Scholar] [CrossRef]
- Hardstone, M.C.; Lazzaro, B.P.; Scott, J.G. The effect of three environmental conditions on the fitness of cytochrome P450 monooxygenase-mediated permethrin resistance in Culex pipiens quinquefasciatus. BMC Evol. Biol. 2009, 9, 42. [Google Scholar] [CrossRef]
- Kumar, S.; Pillai, M.K. Correlation between the reproductive potential and the pyrethroid resistance in an Indian strain of filarial vector, Culex quinquefasciatus Say (Diptera: Culicidae). Bull. Entomol. Res. 2011, 101, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, L.; Sun, L.; Zhu, C. Biotic characteristics in the deltamethrin-susceptible and resistant strains of Culex pipiens pallens (Diptera: Culicidae) in China. Appl. Entomol. Zool. 2002, 37, 305–308. [Google Scholar] [CrossRef]
- Otali, D.; Novak, R.J.; Wan, W.; Bu, S.; Moellering, D.R.; De Luca, M. Increased production of mitochondrial reactive oxygen species and reduced adult life span in an insecticide-resistant strain of Anopheles gambiae. Bull. Entomol. Res. 2014, 104, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Rowland, M. Behaviour and fitness of gamma HCH/dieldrin resistant and susceptible female Anopheles gambiae and An. stephensi mosquitoes in the absence of insecticide. Med. Vet. Entomol. 1991, 5, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Belinato, T.A.; Valle, D. The impact of selection with diflubenzuron, a chitin synthesis inhibitor, on the fitness of two Brazilian Aedes aegypti field populations. PLoS ONE 2015, 10, e0130719. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, O.N.; Fonseca-Gonzalez, I.; Chaverra-Rodriguez, D. Geometric morphometrics of nine field isolates of Aedes aegypti with different resistance levels to lambda-cyhalothrin and relative fitness of one artificially selected for resistance. PLoS ONE 2014, 9, e96379. [Google Scholar] [CrossRef] [PubMed]
- Okoye, P.N.; Brooke, B.D.; Hunt, R.H.; Coetzee, M. Relative developmental and reproductive fitness associated with pyrethroid resistance in the major southern African malaria vector, Anopheles funestus. Bull. Entomol. Res. 2007, 97, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Prapanthadara, L.; Reunkum, W.; Leelepat, P.; Suwan, W.; Yanola, J.; Somboon, P. Glutathione-S-transferase isoenzymes and the DDTase activity in two DDT-resistant strains of Aedes aegypti. Dengue Bull. 2005, 29, 183–191. [Google Scholar]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.A.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Choovattanapakorn, N.; Yanola, J.; Lumjuan, N.; Saingamsook, J.; Somboon, P. Characterization of metabolic detoxifying enzymes in an insecticide resistant strain of Aedes aegypti harboring homozygous S989P and V1016G kdr mutations. Med. Entomol. Zool. 2017, 68, 19–26. [Google Scholar] [CrossRef]
- Son-un, P.; Choovattanapakorn, N.; Saingamsook, J.; Somboon, P.; Yanola, J.; Lumjuan, N.; Walton, C. Effect of Relaxation of Deltamethrin Pressure on Metabolic Resistance in a Pyrethroid-Resistant Aedes aegypti (Diptera: Culicidae) Strain Harboring Fixed P989P and G1016G kdr Alleles. J. Med. Entomol. 2018, 55, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Saingamsook, J.; Saeung, A.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. A multiplex PCR for detection of knockdown resistance mutations, V1016G and F1534C, in pyrethroid-resistant Aedes. aegypti. Parasit. Vectors 2017, 10, 465. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, C.; Saingamsook, J.; Somboon, P. A simple and affordable membrane-feeding method for Aedes aegpyti and Anopheles minimus (Diptera: Culicidae). Acta Trop. 2015, 152, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Rattanarithikul, R.; Harbach, R.E.; Harrison, B.A.; Panthusiri, P.; Coleman, R.E.; Richardson, J.H. Illustrated keys to the mosquitoes of Thailand. VI. Tribe Aedini. Southeast Asian J. Trop. Med. Public Health 2010, 41, 1–225. [Google Scholar]
- Platt, N.; Kwiatkowska, R.M.; Irving, H.; Diabaté, A.; Dabire, R.; Wondji, C.S. Target-site resistance mutations (kdr and RDL), but not metabolic resistance, negatively impact male mating competiveness in the malaria vector Anopheles gambiae. Heredity 2015, 115, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Bourguet, D.; Guillemaud, T.; Chevillon, C.; Raymond, M. Fitness costs of insecticide resistance in natural breeding sites of the mosquito Culex pipiens. Evolution 2004, 58, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Van Uitregt, V.O.; Hurst, T.P.; Wilson, R.S. Reduced size and starvation resistance in adult mosquitoes, Aedes notoscriptus, exposed to predation cues as larvae. J. Anim. Ecol. 2012, 81, 108–115. [Google Scholar] [CrossRef]
- Plernsub, S.; Stenhouse, S.A.; Tippawangkosol, P.; Lumjuan, N.; Yanola, J.; Choochote, W.; Somboon, P. Relative developmental and reproductive fitness associated with F1534C homozygous knockdown resistant gene in Aedes aegypti from Thailand. Trop Biomed. 2013, 30, 621–630. [Google Scholar]
- Christopher, S.R. (Ed.) Aedes Aegypti: The Yellow Fever Mosquito; Cambridge University Press: London, UK, 1960; p. 739. [Google Scholar]
- Scott, T.W.; Naksathit, A.; Day, J.F.; Kittayapong, P.; Edman, J.D. A fitness advantage for Aedes aegypti and the viruses it transmits when females feed only on human blood. Am. J. Trop. Med. Hyg. 1997, 57, 235–239. [Google Scholar] [CrossRef]
- Clements, A.N. The Biology of Mosquitoes, Volume 1: Development, Nutrition, and Reproduction; Chapman & Hall: London, UK, 1992; p. 509. [Google Scholar]
- Helinski, M.E.; Harrington, L.C. Male mating history and body size influence female fecundity and longevity of the dengue vector Aedes aegypti. J. Med. Entomol. 2011, 48, 202–211. [Google Scholar] [CrossRef]
- Ponlawat, A.; Harrington, L.C. Age and body size influence male sperm capacity of the dengue vector Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 422–426. [Google Scholar] [CrossRef]
- Ponlawat, A.; Harrington, L.C. Factors associated with male mating success of the dengue vector mosquito, Aedes aegypti. Am. J. Trop. Med. Hyg. 2009, 80, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Berticat, C.; Boquien, G.; Raymond, M.; Chevillon, C. Insecticide resistance genes induce a mating competition cost in Culex pipiens mosquitoes. Genet. Res. 2002, 79, 41–47. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Strain (Mean ± SE) * | |||
|---|---|---|---|---|
| PMD | PMD-R | UPK-R | Hybrid | |
| Pupation time (day) 1 | 11.6 ± 0.1 a | 11.5 ± 0.1 a | 12.6 ± 0.1 b | 11.8 ± 0.1 a |
| Pupation rate (%) 2 | 89.5 ± 4.0 a | 85.0 ± 2.5 a | 89.0 ± 1.3 a | 95.0 ± 1.7 a |
| Adult emergence time (day) 3 | 14.9 ± 0.1 a | 14.6 ± 0.1 a | 15.8 ± 0.1 b | 14.9 ± 0.1 a |
| Adult emergence rate (%) 4 | 89.0 ± 3.8 a | 85.0 ± 2.5 a | 87.5 ± 1.0 a | 95.0 ± 1.7 a |
| Wing length (mm) | 3.29 ± 0.02 a | 2.98 ± 0.03 b | 3.14 ± 0.02 c | 3.08 ± 0.01 c |
| Eggs/female (egg) | 94.8 ± 2.6 a | 74.7 ± 3.8 b | 91.9 ± 1.7 a | 87.4 ± 1.6 a |
| Eggs hatching rate (%) | 96.0 ± 0.6 a | 84.7 ± 0.3 b | 84.7 ± 0.3 b | 80.7 ± 0.9 c |
| Parameter | PMD | PMD-R | UPK-R | Hybrid |
|---|---|---|---|---|
| Blood feeding rate (%) | 94.0 ± 0.6 a | 93.0 ± 0.6 a | 95.0 ± 1.2 a | 95.7 ± 0.3 a |
| No. of pool 1 | 6 | 6 | 6 | 7 |
| Before blood meal (mg) 2 | 12.58 ± 0.23 a | 9.90 ± 0.61 b | 10.75 ± 0.12 b | 11.54 ± 0.31 a |
| After blood meal (mg) 2 | 27.55 ± 0.45 a | 20.37 ± 0.73 b | 22.18 ± 0.28 b,c | 23.83 ± 0.43 c |
| Weight of blood ingested (mg) 2 | 14.97 ± 0.25 a | 10.47 ± 0.76 b | 11.43 ± 0.31 b | 12.29 ± 0.43 b |
| Weight increase ratio | 2.19 ± 0.02 a | 2.09 ± 0.13 a | 2.07 ± 0.03 a | 2.07 ± 0.06 a |
| Life-Trait Parameter | PMD-R | UPK-R | Hybrid |
|---|---|---|---|
| Developmental time (larval to adult) | equal | longer | equal |
| Wing length | shorter | shorter | shorter |
| Blood feeding rate | equal | equal | equal |
| Blood ingestion volume | smaller | smaller | equal |
| Eggs/female | lower | equal | equal |
| Egg hatchability | lower | lower | lower |
| Egg viability duration | shorter | shorter | shorter |
| Adult longevity | shorter | shorter | shorter |
| Kdr Alleles | No. of Mosquito (%) | Total | |
|---|---|---|---|
| Cage | |||
| A | B | ||
| 1016V/V + 1534C/C | 5 (16.7) | 4 (13.4) | 9 (15.0) |
| 1016G/G + 1534F/F | 10 (33.3) | 13 (43.3) | 23 (38.3) |
| 1016V/G + 1534F/C | 15 (50.0) | 13 (43.3) | 28 (46.7) |
| Total | 30 (100.0) | 30 (100.0) | 60 (100.0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saingamsook, J.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand. Insects 2019, 10, 265. https://doi.org/10.3390/insects10090265
Saingamsook J, Yanola J, Lumjuan N, Walton C, Somboon P. Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand. Insects. 2019; 10(9):265. https://doi.org/10.3390/insects10090265
Chicago/Turabian StyleSaingamsook, Jassada, Jintana Yanola, Nongkran Lumjuan, Catherine Walton, and Pradya Somboon. 2019. "Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand" Insects 10, no. 9: 265. https://doi.org/10.3390/insects10090265