Yeast Species, Strains, and Growth Media Mediate Attraction of Drosophila suzukii (Diptera: Drosophilidae)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Colony
2.2. Yeast Isolation from Macerated Blackberry and Raspberry Fruits
2.3. Yeast Identification
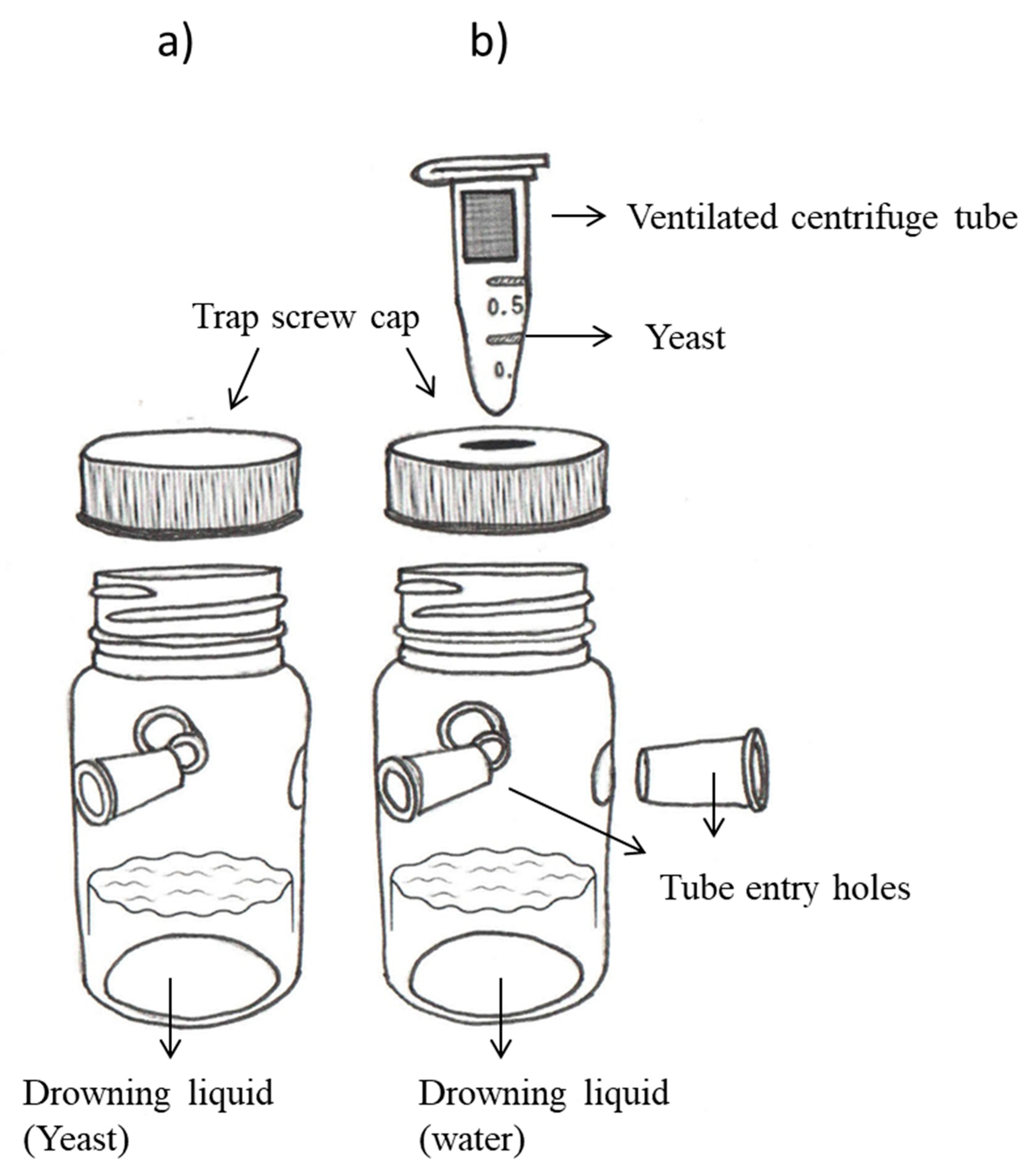
2.4. Attraction of D. suzukii to Yeast Isolates from Blackberry and Raspberry Fruit
2.4.1. Initial Evaluation of Isolates
2.4.2. Additional Evaluation of RiM2-Hu Isolate from Raspberry
2.5. Attraction of D. suzukii to Other Yeast Species
2.6. Statistical Analyses
3. Results
3.1. Yeast Identification
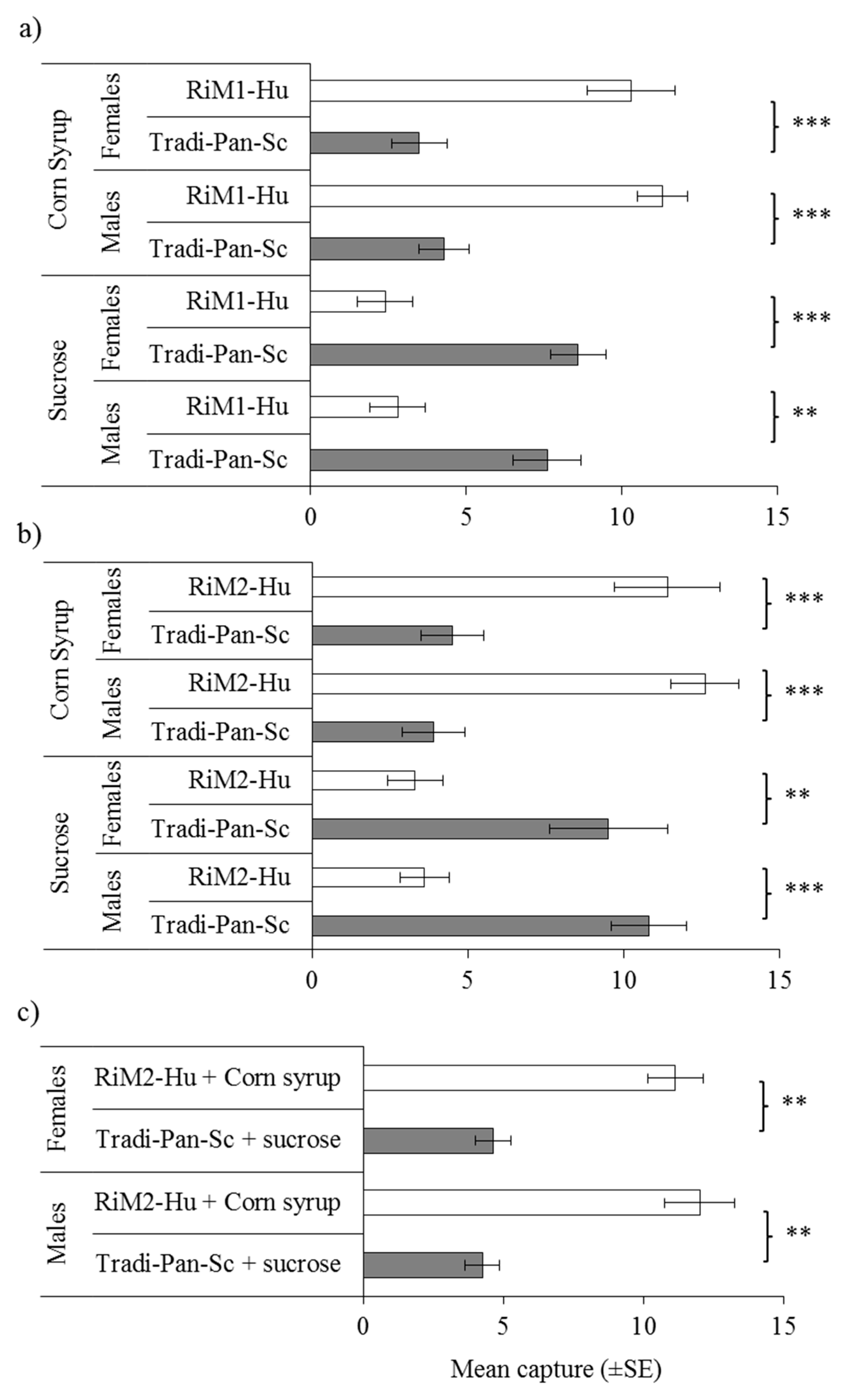
3.2. Attraction of D. suzukii to Yeasts Isolated from Blackberry and Raspberry Fruits
3.3. Attraction of D. suzukii to Species of Candida, Pichia, and Hanseniaspora
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.; Witzgall, P. Combining mutualistic yeast and pathogenic virus: A novel method for codling moth control. J. Chem. Ecol. 2013, 39, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.; Basoalto, E.; Yee, W.; Hilton, R.; Kurtzman, C. Adding yeasts with sugar to increase the number of effective insecticide classes to manage Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in cherry. Pest Manag. Sci. 2015, 72, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Hamby, K.A.; Becher, P.G. Current knowledge of interactions between Drosophila suzukii and microbes, and their potential utility for pest management. J. Pest Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Begon, M. Yeasts and Drosophila. In Genetics and Biology of Drosophila; Ashburner, M., Carson, H.L., Thomson, J.N., Jr., Eds.; Academic Press: London, UK, 1982; pp. 345–384. [Google Scholar]
- Hamby, K.A.; Hernández, A.; Boundy-Mills, K.; Zalom, F. Associations of yeasts with spotted-wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in cherries and raspberries. Appl. Environ. Microbiol. 2012, 78, 4869–4873. [Google Scholar] [CrossRef] [PubMed]
- Markow, T.A.; O’Grady, P. Reproductive ecology of Drosophila. Func. Ecol. 2008, 22, 747–759. [Google Scholar] [CrossRef]
- Reuter, M.; Bell, G.; Greig, D. Increased outbreeding in yeast in response to dispersal by an insect vector. Curr. Biol. 2007, 17, R81–R83. [Google Scholar] [CrossRef] [Green Version]
- Bellutti, N.; Gallmetzer, A.; Innerebner, G.; Schmidt, S.; Zelger, R.; Koschier, E.H. Dietary yeast affects preference and performance in Drosophila suzukii. J. Pest Sci. 2018, 91, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.S.; Goddard, M.R. Do yeasts and Drosophila interact just by chance? Fungal Ecol. 2019, 38, 37–43. [Google Scholar] [CrossRef]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron. 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Palanca, L.; Gaskett, A.C.; Günther, C.S.; Newcomb, R.D.; Goddard, M.R. Quantifying variation in the ability of yeast to attract Drosophila melanogaster. PLoS ONE 2013, 8, e75332. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.S.; Goddard, M.R.; Newcomb, R.D.; Buser, C.C. The context of chemical communication driving a mutualism. J. Chem. Ecol. 2015, 41, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.S.; Knight, S.J.; Jones, R.; Goddard, M.R. Are Drosophila preferences for yeasts stable or contextual? Ecol. Evol. 2019. [Google Scholar] [CrossRef]
- Starmer, W.T.; Fogleman, J.C. Coadaptation of Drosophila and yeasts in their natural habitat. J. Chem. Ecol. 1986, 12, 1037–1055. [Google Scholar] [CrossRef]
- Chandler, J.A.; Eisen, J.A.; Kopp, A. Yeast communities of diverse Drosophila species: Comparison of two symbiont groups in the same hosts. Appl. Environ. Microbiol. 2012, 78, 7327–7336. [Google Scholar] [CrossRef]
- Frewin, A.J.; Renquema, J.; Fraser, H.; Hallett, H. Evaluation of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2017, 110, 1156–1163. [Google Scholar] [CrossRef]
- Hampton, E.; Koski, C.; Barsoian, O.; Faubert, H.; Cowles, R.S.; Alm, S.R. Use of early ripening cultivars to avoid infestation and mass trapping to manage Drosophila suzukii (Diptera: Drosophilidae) in Vaccinium corymbosum (Ericales: Ericaceae). J. Econ. Entomol. 2014, 107, 1849–1857. [Google Scholar] [CrossRef]
- Iglesias, L.E.; Nyoike, T.W.; Liburd, O.E. Effect of trap design, bait type, and age on captures of Drosophila suzukii (Diptera: Drosophilidae) in berry crops. J. Econ. Entomol. 2014, 107, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Tonina, L.; Grassi, A.; Caruso, S.; Mori, N.; Gottardello, A.; Anfora, G.; Giomi, F.; Vaccari, G.; Ioriatti, C. Comparison of attractants for monitoring Drosophila suzukii in sweet cherry orchards in Italy. J. Appl. Entomol. 2018, 142, 18–25. [Google Scholar] [CrossRef]
- Scheidler, N.; Liu, C.; Hamby, K.; Zalom, F.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [PubMed]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, D.; Walton, V.; Shearer, P.; Walsh, D.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- NCBI GenBank. Available online: http://ncbi.nlm.nih.gov/ (accessed on 12 July 2019).
- Resende, L.V.; Pinheiro, L.K.; Miguel, M.G.D.C.P.; Ramos, C.L.; Vilela, D.M.; Schwan, R.F. Microbial community and physicochemical dynamics during the production of ‘Chicha’, a traditional beverage of indigenous people of Brazil. World J. Microbiol. Biotechnol. 2018, 34, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Jamovi. Jamovi Statistical Software v. 0.9. 2019. Available online: https://www.jamovi.org (accessed on 14 June 2019).
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Sicard, D.; Legras, J.C. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. Comp. Red. Biol. 2011, 344, 229–236. [Google Scholar] [CrossRef]
- Dobzhansky, T.; Cooper, D.M.; Phaff, H.J.; Knapp, E.P.; Carson, H.L. Studies on the ecology of the Drosophila in the Yosemite region of California: IV. Differential attraction of species of Drosophila to different species of yeasts. Ecology 1956, 37, 544–550. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef]
- Yarrow, D. Methods for the isolation, maintenance, classification and identification of yeasts. In the Yeasts, a Taxonomic Study, 4th ed.; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 77–100. [Google Scholar]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. How yeasts are classified. In Yeasts: Characteristics and Identification, 3rd ed.; Barnett, J.A., Payne, R.W., Yarrow, D., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 15–22. [Google Scholar]
- Gethins, L.; Guneser, O.; Demirkol, A.; Rea, M.C.; Stanton, C.; Ross, R.P.; Yuceer, Y.; Morrissey, J.P. Influence of carbon and nitrogen source on production of volatile fragrance and flavour metabolites by the yeast Kluyveromyces marxianus. Yeast 2015, 32, 67–76. [Google Scholar] [PubMed]
- Crépin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by Saccharomyces cerevisiae during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef] [PubMed]
- Jiranek, V.; Langridge, P.; Henschke, P.A. Amino acid and ammonium utilization by Saccharomyces cerevisiae wine yeasts from a chemically defined medium. Am. J. Enol. Vitic. 1995, 46, 75–83. [Google Scholar]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The impact of single amino acids on growth and volatile aroma production by Saccharomyces cerevisiae strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [PubMed]
- Marcus, M. Bait and Trap Design Preferences for Drosophila suzukii. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2014. Available online: http://hdl.handle.net/1957/52499 (accessed on 12 July 2019).
- Lam, S.S.; Howell, K.S. Drosophila-associated yeast species in vineyard ecosystems. FEMS Microbiol. Lett. 2015, 362, 170. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Code | Species | Source a | Origen | Accession No. |
|---|---|---|---|---|
| Tradi-Pan | Saccharomyces cerevisiae | Dry baker’s yeast | Safmex SA de CV | MN153007 |
| RfM1 | Saccharomyces cerevisiae | Blackberry | Isolated | MN153005 |
| RfM2 | Hanseniaspora uvarum | Blackberry | Isolated | MN153006 |
| RfM3 | Hanseniaspora uvarum | Blackberry | Isolated | MN153004 |
| RiM1 | Hanseniaspora uvarum | Raspberry | Isolated | MN153010 |
| RiM2 | Hanseniaspora uvarum | Raspberry | Isolated | MN153008 |
| 5HL | Hanseniaspora lachancei | Mescal | Collection CIATEJ | - |
| 1CB | Candida boidinii | Tejuino | Collection CIATEJ | - |
| 2CG | Candida glabrata | Tejuino | Collection CIATEJ | - |
| 3CH | Candida humilis | Tejuino | Collection CIATEJ | - |
| 4 CT | Candida tropicalis | Tejuino | Collection CIATEJ | - |
| 6PA | Pichia anomala | Mescal | Collection CIATEJ | - |
| 7PKL | Pichia kluyveri | Mezcal | Collection CIATEJ | - |
| 8PKU | Pichia kudriavzevii | Cacao | Collection CIATEJ | - |
| 9PMA | Pichia manshurica | Mescal | Collection CIATEJ | - |
| 10PME | Pichia membranifaciens | Mescal | Collection CIATEJ | - |
| Species and Code | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| 1. Saccharomyces cerevisiae KC881066.1 * | - | 85.7 | 100 | 100 | 85.9 | 85.9 | 85.7 | 85.7 |
| 2. Hanseniaspora uvarum KY107844.1 * | - | 85.7 | 85.7 | 99.8 | 99.8 | 100 | 100 | |
| 3. S. cerevisiae Tradi-pan | - | 100 | 85.9 | 85.9 | 85.7 | 85.7 | ||
| 4. S. cerevisiae RfM1 | - | 85.9 | 85.9 | 85.7 | 85.7 | |||
| 5. H. uvarum RfM2 | - | 100 | 99.8 | 99.8 | ||||
| 6. H. uvarum RfM3 | — | 99.8 | 99.8 | |||||
| 7. H. uvarum RiM1 | — | 100 | ||||||
| 8. H. uvarum RiM2 | — |
| Mean ± SE | ||||
|---|---|---|---|---|
| Yeasts | Males | Females | ||
| Saccharomyces cerevisiae | 8.3 ± 1.6 | NS | 7.3 ± 1.5 | NS |
| Candida boidinii | 6.9 ± 1.6 | 6.8 ± 1.6 | ||
| S. cerevisiae | 7.3 ± 1.0 | NS | 7.9 ± 1.3 | NS |
| Candida glabrata | 4.1 ± 1.6 | 4.1 ± 1.3 | ||
| S. cerevisiae | 4.0 ± 1.1 | NS | 4.6 ± 0.9 | NS |
| Candida humilis | 5.3 ± 0.9 | 5.8 ± 1.2 | ||
| S. cerevisiae | 4.9 ± 0.9 | NS | 6.0 ± 1.0 | NS |
| Candida tropicalis | 8.1 ± 1.2 | 5.8 ± 1.1 | ||
| S. cerevisiae | 3.6 ± 0.9 | NS | 4.5 ± 0.9 | NS |
| Hanseniaspora lachancei | 3.0 ± 0.5 | 3.0 ± 0.8 | ||
| S. cerevisiae | 7.6 ± 1.0 | * | 8.9 ± 1.5 | NS |
| Pichia anomala | 3.5 ± 0.6 | 5.3 ± 1.0 | ||
| S. cerevisiae | 9.6 ± 0.8 | *** | 8.6 ± 0.7 | *** |
| Pichia kluyveri | 1.1 ± 0.6 | 2.1 ± 0.7 | ||
| S. cerevisiae | 3.6 ± 0.9 | NS | 6.6 ± 0.4 | NS |
| Pichia kudriavzevii | 4.4 ± 0.9 | 4.3 ± 1.2 | ||
| S. cerevisiae | 6.8 ± 1.4 | NS | 7.9 ± 1.2 | * |
| Pichia manshurica | 3.1 ± 0.5 | 3.0 ± 0.6 | ||
| S. cerevisiae | 6.1 ± 0.8 | NS | 7.4 ± 1.3 | NS |
| Pichia membranifaciens | 5.0 ± 0.4 | 3.4 ± 1.0 | ||
| Mean ± SE | ||||
|---|---|---|---|---|
| Yeasts | Males | Females | ||
| Saccharomyces cerevisiae | 8.3 ± 1.6 | NS | 7.3 ± 1.5 | NS |
| Candida boidinii | 6.9 ± 1.6 | 6.8 ± 1.6 | ||
| S. cerevisiae | 5.9 ± 1.0 | NS | 5.3 ± 0.8 | NS |
| Candida glabrata | 6.3 ± 0.6 | 4.9 ± 1.1 | ||
| S. cerevisiae | 5.3 ± 1.4 | NS | 5.1 ± 0.7 | NS |
| Candida humilis | 4.9 ± 1.2 | 5.0 ± 1.2 | ||
| S. cerevisiae | 5.1 ± 0.9 | NS | 4.6 ± 1.4 | NS |
| Candida tropicalis | 7.5 ± 1.7 | 6.1 ± 1.5 | ||
| S. cerevisiae | 7.3 ± 1.4 | NS | 6.8 ± 0.5 | NS |
| Hanseniaspora lachancei | 4.6 ± 0.9 | 5.4 ± 0.8 | ||
| S. cerevisiae | 6.6 ± 1.2 | NS | 9.9 ± 0.6 | * |
| Pichia anomala | 4.6 ± 0.8 | 5.5 ± 0.8 | ||
| S. cerevisiae | 9.1 ± 0.7 | * | 9.3 ± 0.9 | * |
| Pichia kluyveri | 3.6 ± 0.8 | 2.9 ± 0.6 | ||
| S. cerevisiae | 4.8 ± 1.0 | NS | 6.5 ± 0.8 | NS |
| Pichia kudriavzevii | 3.6 ± 0.7 | 4.5 ± 0.9 | ||
| S. cerevisiae | 7.5 ± 1.2 | NS | 10.5 ± 0.9 | *** |
| Pichia manshurica | 3.4 ± 1.0 | 3.0 ± 0.6 | ||
| S. cerevisiae | 7.0 ± 1.3 | NS | 8.5 ± 1.3 | * |
| Pichia membranifaciens | 2.5 ± 0.8 | 2.6 ± 0.9 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lasa, R.; Navarro-de-la-Fuente, L.; Gschaedler-Mathis, A.C.; Kirchmayr, M.R.; Williams, T. Yeast Species, Strains, and Growth Media Mediate Attraction of Drosophila suzukii (Diptera: Drosophilidae). Insects 2019, 10, 228. https://doi.org/10.3390/insects10080228
Lasa R, Navarro-de-la-Fuente L, Gschaedler-Mathis AC, Kirchmayr MR, Williams T. Yeast Species, Strains, and Growth Media Mediate Attraction of Drosophila suzukii (Diptera: Drosophilidae). Insects. 2019; 10(8):228. https://doi.org/10.3390/insects10080228
Chicago/Turabian StyleLasa, Rodrigo, Laura Navarro-de-la-Fuente, Anne C. Gschaedler-Mathis, Manuel R. Kirchmayr, and Trevor Williams. 2019. "Yeast Species, Strains, and Growth Media Mediate Attraction of Drosophila suzukii (Diptera: Drosophilidae)" Insects 10, no. 8: 228. https://doi.org/10.3390/insects10080228