Antennal Morphology and Sexual Dimorphism of Antennal Sensilla in Callitettix versicolor (Fabricius) (Hemiptera: Cercopidae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collecting
2.2. Image Processing and Morphometric Measurement
3. Results
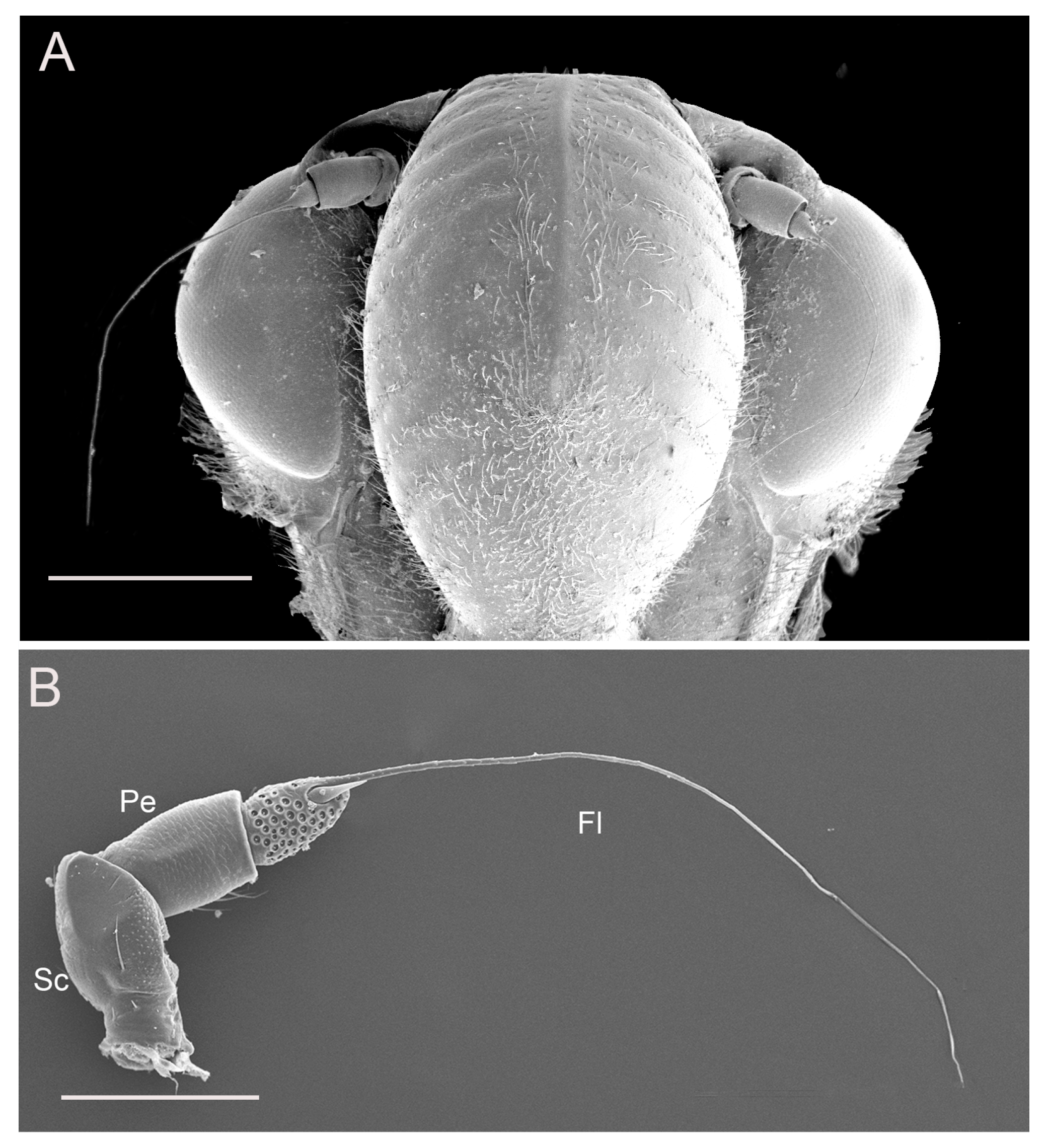
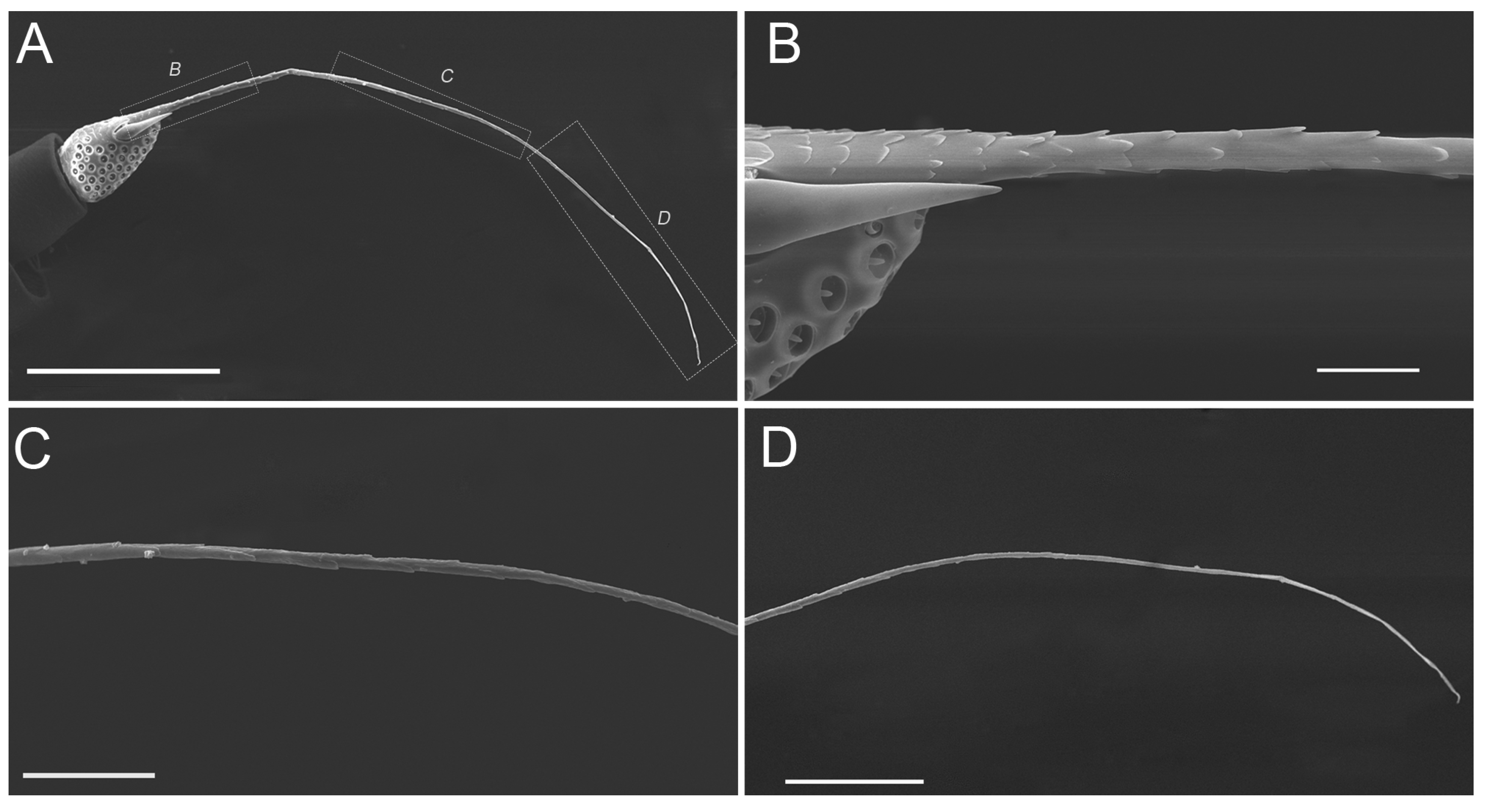
3.1. Gross Morphology of Antennae
3.2. Sensilla Types
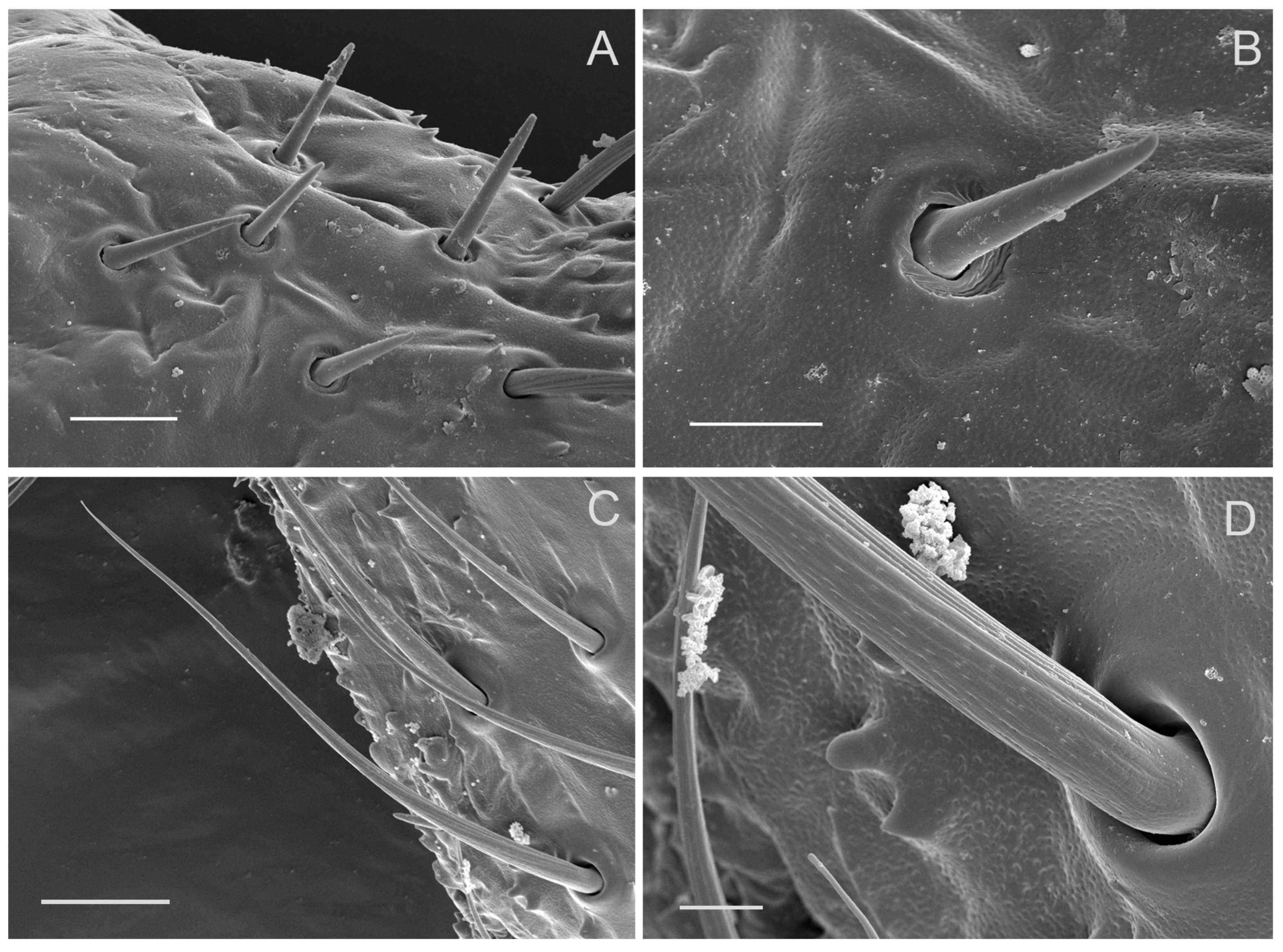
3.3. Sensilla Trichodea (ST)
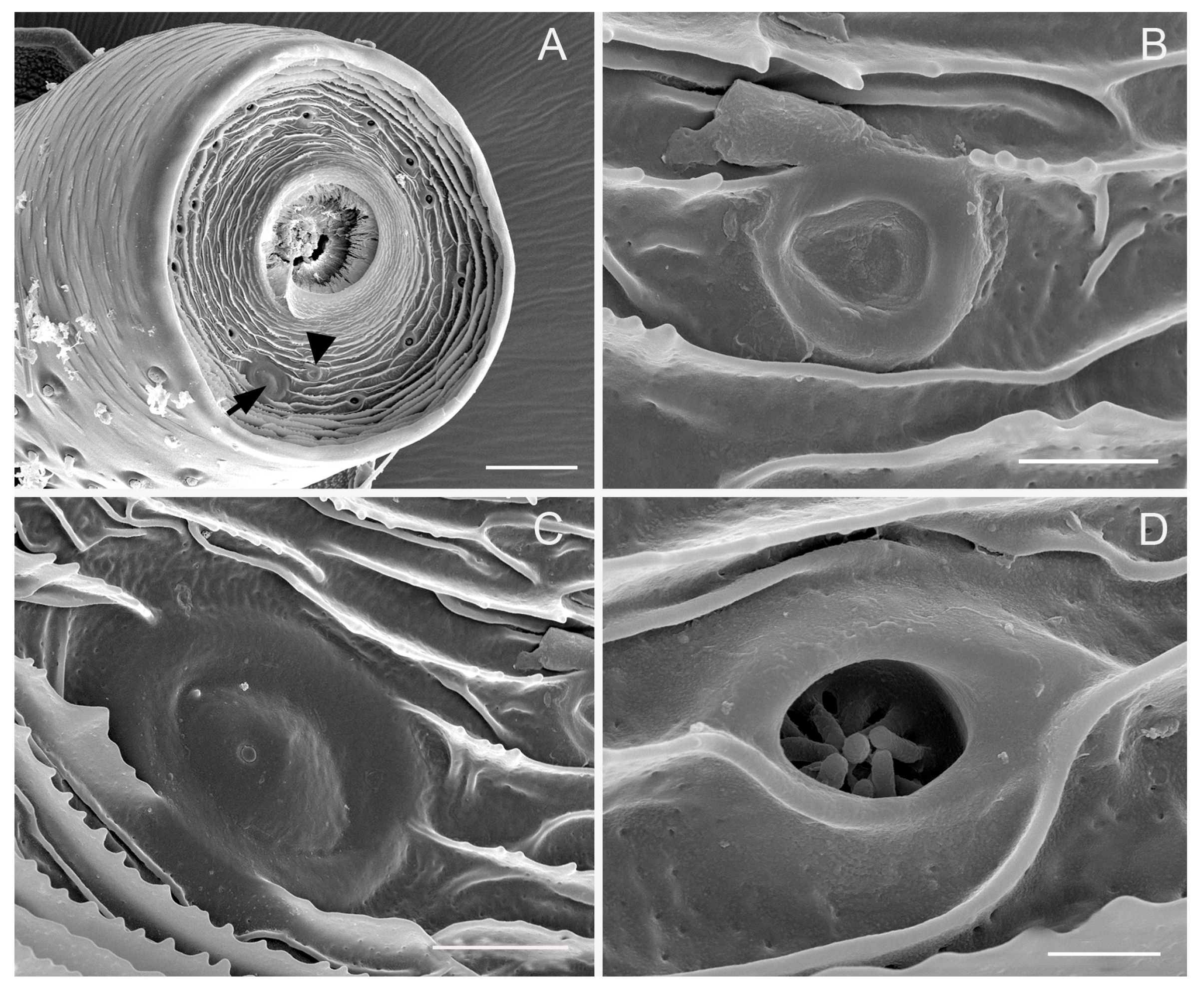
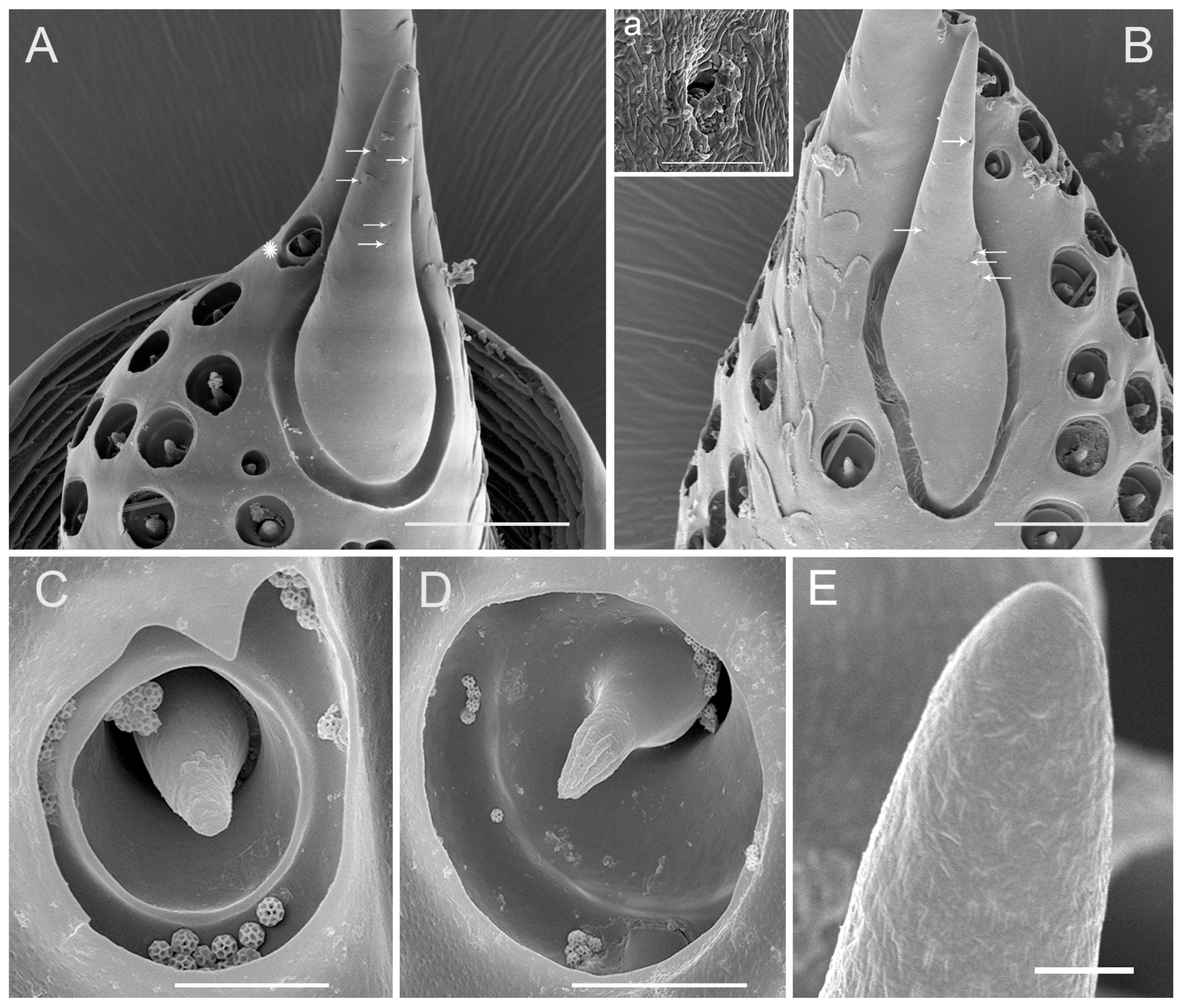
3.4. Sensilla Coeloconica (SCo)
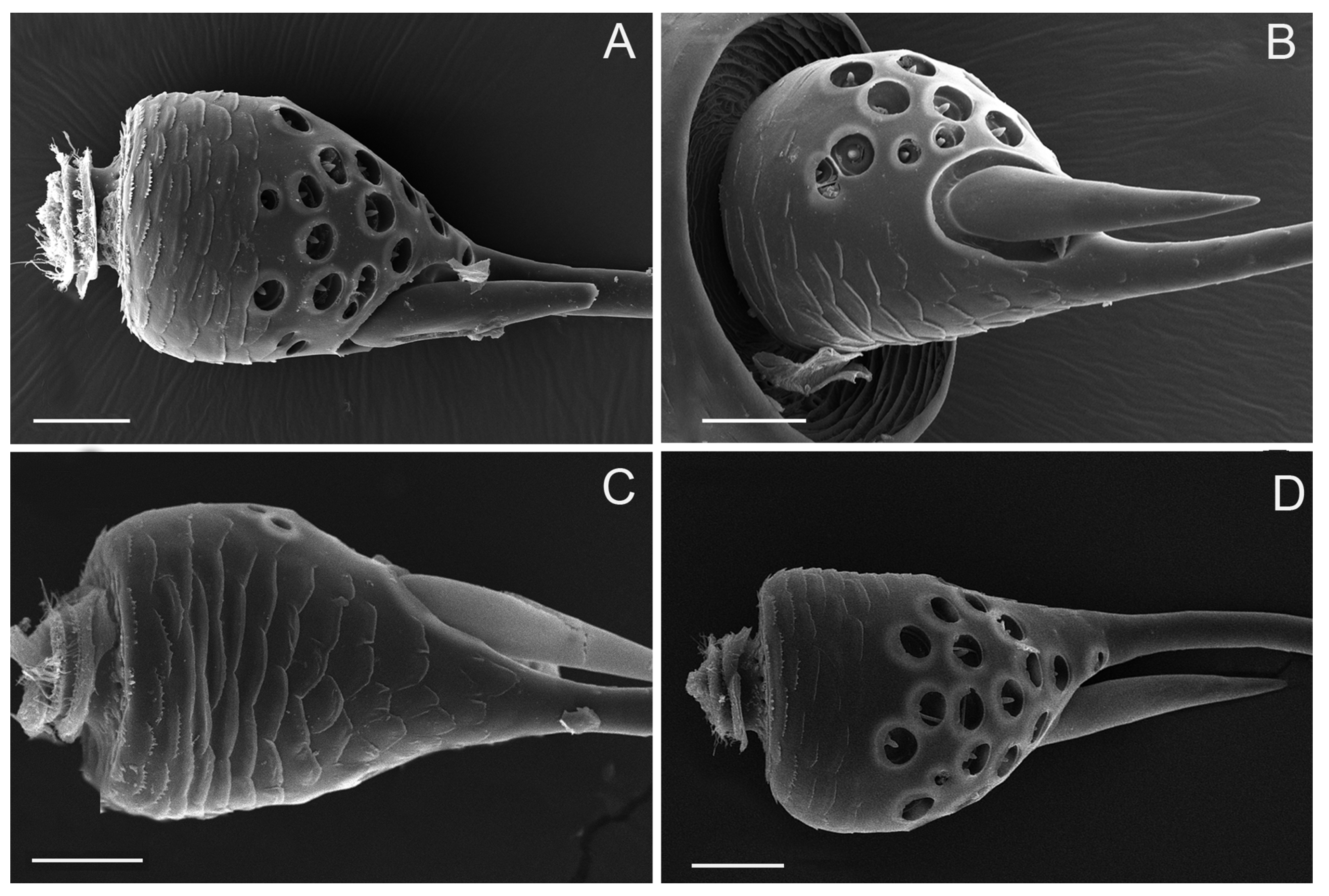
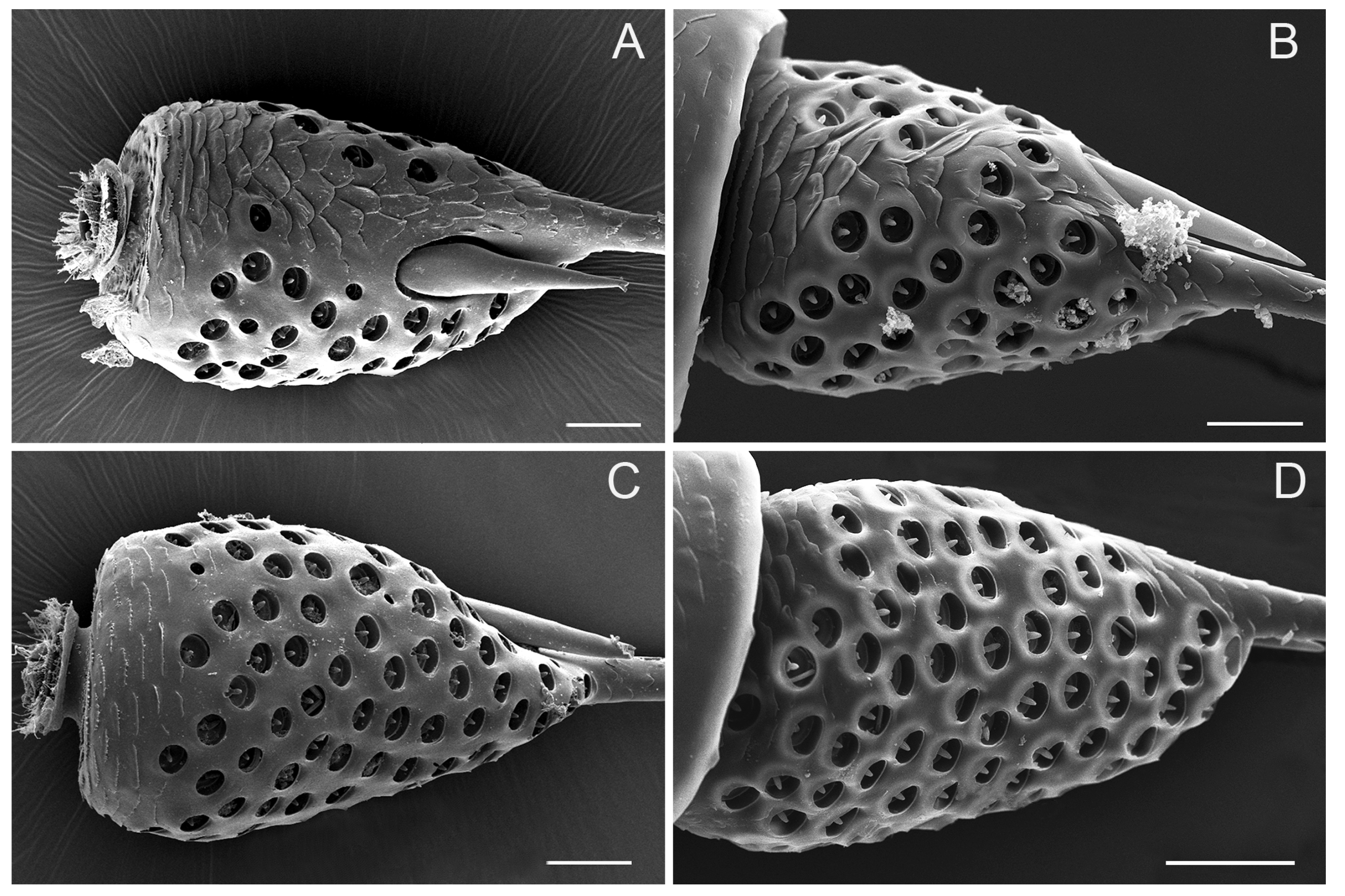
3.5. Sensilla Basiconica (SB)
3.6. Sensilla Campaniformia (SCa)
4. Discussion
4.1. Types and Functions of Antennal Sensilla
4.2. Morphological Characters
4.3. Sexual Dimorphism of Olfactory Sensilla
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liang, A.P.; Webb, M.D. New taxa and revisionary notes in Rhinaulacini spittlebugs from southern Asia (Homoptera: Cercopidae). J. Nat. Hist. 2002, 36, 729–756. [Google Scholar] [CrossRef]
- Chen, X.S.; Liang, A.P. Laboratory rearing of Callitettix versicolor (Hemiptera: Cicadomorpha: Cercopidae), with descriptions of the immature stages. Ann. Entomol. Soc. Am. 2012, 105, 664–670. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Antennae and sensilla. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; Volume 6, pp. 1–69. [Google Scholar]
- Chapman, R.F. Mechanoreception. Chemoreception. In The Insects: Structure and Function, 4th ed.; Chapman, R.F., Ed.; Cambridge University Press: New York, NY, USA, 1998; pp. 610–652. [Google Scholar]
- Ruschioni, S.; Riolo, P.; Verdolini, E.; Peri, E.; Guarino, S.; Colazza, S.; Romani, R.; Isidoro, N. Fine structure of antennal sensilla of Paysandisia archon and electrophysiological responses to volatile compounds associated with host palms. PLoS ONE 2015, 10, e0124607. [Google Scholar] [CrossRef] [PubMed]
- Bartlet, E.; Isidoro, N.; Williams, I. Antennal glands in Psylliodes chrysocephala and their possible role in reproductive behavior. Physiol. Entomol. 1994, 19, 241–250. [Google Scholar] [CrossRef]
- Romani, R.; Rosi, M.C.; Isidoro, N.; Bin, F. The role of the antennae during courtship behaviour in the parasitic wasp Trichopria drosophilae. J. Exp. Biol. 2008, 211, 2486–2491. [Google Scholar] [CrossRef]
- Hamilton, K.G.A. Revision of neotropical Aphrophorinae spittlebugs, part 1: Ptyelini (Hemiptera, Cercopoidea). Zootaxa 2012, 3497, 41–59. [Google Scholar]
- Wang, R.R.; Liu, J.J.; Li, X.Y.; Liang, A.P.; Bourgoin, T. Relating antennal sensilla diversity and possible species behaviour in the planthopper pest Lycorma delicatula (Hemiptera: Fulgoromorpha: Fulgoridae). PLoS ONE 2018, 13, e0194995. [Google Scholar] [CrossRef] [PubMed]
- Onagbola, E.O.; Meyer, W.L.; Boina, D.R.; Stelinski, L.L. Morphological characterization of the antennal sensilla of the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), with reference to their probable functions. Micron 2008, 39, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.; De, C.G.; Moraes, M.C.; Falcão, R.; Solino, L.F.; Laumann, R.A.; Silva, J.P.; Borges, M. Morphology, distribution and abundance of antennal sensilla in three stink bug species (Hemiptera: Pentatomidae). Micron 2010, 41, 289–300. [Google Scholar] [CrossRef]
- Romani, R.; Stacconi, M.V.; Riolo, P.; Isidoro, N. The sensory structures of the antennal flagellum in Hyalesthes obsoletus (Hemiptera: Fulgoromorpha: Cixiidae): A functional reduction? Arthropod Struct. Dev. 2009, 38, 473–483. [Google Scholar] [CrossRef]
- Wang, R.R.; Wan, X.Y.; Liang, A.P.; Bourgoin, T.A. SEM study of antennal and maxillary sensilla in Zema gressitti Fennah (Hemiptera: Fulgoromorpha: Tropiduchidae). Micron 2013, 44, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.N.; Pan, L.X.; Lu, Y.P.; Dietrich, C.H.; Dai, W. Reinvestigation of the antennal morphology of the white-backed planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Zool. Anz. 2016, 262, 20–28. [Google Scholar] [CrossRef]
- Ahmad, A.; Parveen, S.; Brozek, J.; Dey, D. Antennal sensilla of phytophagous and predatory pentatomids (Hemiptera: Pentatomidae): A comparative study of four genera. Zool. Anz. 2016, 261, 48–55. [Google Scholar] [CrossRef]
- Nowińska, A.; Brożek, J. Morphological study of the antennal sensilla in Gerromorpha (Insecta: Hemiptera: Heteroptera). Zoomorphology 2017, 136, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Liang, A.P.; Fletcher, M.J. Morphology of the antennal sensilla in four Australian spittlebug species (Hemiptera: Cercopidae) with implications for phylogeny. Aust. J. Entomol. 2002, 41, 39–44. [Google Scholar] [CrossRef]
- Boulard, M.; Boulard, J. Un organe énigmatique chez les femelles de Locris (Homoptera Cercopidae). Ann. Soc. Entomol. Fr. 1979, 15, 513–523. [Google Scholar]
- Ranieri, E.; Ruschioni, S.; Riolo, P.; Isidoro, N.; Romani, R. Fine structure of antennal sensilla of the spittlebug Philaenus spumarius, L. (Insecta: Hemiptera: Aphrophoridae) Chemoreceptors and thermo-/hygroreceptors. Arthropod Struct. Dev. 2016, 45, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Paladini, A.; Carvalho, G.S.; Valério, J.R. Ultrastructure and redescription of Notozulia entreriana (Berg) (Hemiptera: Cercopidae). Neotrop. Entomol. 2008, 37, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D. Insect antennae. Ann. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Bourgoin, T.; Deiss, V. Sensory plate organs of the antenna in the Meenoplidae-Kinnaridae group (Hemiptera: Fulgoromorpha). Int. J. Insect Morphol. Embryol. 1994, 23, 159–168. [Google Scholar] [CrossRef]
- Cheng, C.L.; Yang, C.T. Antennal sensory plaque organs of Fulgoridae (Homoptera: Fulgoroidea). Chin. J. Entomol. 1996, 16, 187–207. [Google Scholar]
- Marshall, A.T.; Lewis, C.T. Structural variation in the antennal sense organs of fulgoroid Homoptera (Insecta). Zool. J. Linn. Soc. 1971, 50, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Shih, H.T.; Yang, C.T. Antennal sensory plaque organs of the Cixiidae (Homoptera: Fulgoroidea). Chin. J. Entomol. 1997, 17, 33–39. [Google Scholar]
- Wang, R.R.; Wan, X.Y.; Liang, A.P.; Bourgoin, T. Ultrastructure of sensory equipment on the heads of Kallitaxila granulata (Stal) (Hemiptera: Fulgoromorpha: Tropiduchidae). Microsc. Res. Tech. 2012, 75, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Paladini, A.; Takiya, D.M.; Cavichioli, R.R.; Carvalho, G.S. Phylogeny and biogeography of Neotropical spittlebugs (Hemiptera: Cercopidae: Ischnorhininae): Revised tribal classification based on morphological data. Syst. Entomol. 2015, 40, 82–108. [Google Scholar] [CrossRef]
- Liang, A.P. Revision, phylogeny, and biogeography of the spittlebug tribe Capnodistini (Hemiptera: Cercopoidea: Aphrophoridae) from Southeast Asia. Ann. Entomol. Soc. Am. 2016, 108, 383–402. [Google Scholar] [CrossRef]
- Liang, A.P. Scanning electron microscopy of antennal sense organs in Prosapia bicincta (Say) (Hemiptera: Cercopidae). J. Entomol. Sci. 2001, 36, 335–341. [Google Scholar] [CrossRef]
- Paladini, A.; Carvalho, G.S.; Cavichioli, R.R. Taxonomic review of Sphenoclypeana and cladistic analysis of Ischnorhinini (Hemiptera: Cercopidae: Tomapidinae). Zootaxa 2010, 2502, 24–36. [Google Scholar] [CrossRef]
- Carvalho, G.S.; Sakakibara, A.M.; Webb, M.D. Two new species of the neotropical spittlebug genus Monecphora Amyot & Serville (Hemiptera: Cercopidae) with key and notes of species of the genus. Zootaxa 2016, 4078, 143. [Google Scholar]
- Shih, H.T.; Yang, J.T. Supplement on the species Aphrophora horizontalis Kato (Hemiptera: Cercopoidea: Aphrophoridae). Plant Prot. B 2005, 47, 77. [Google Scholar]
- Cryan, J.R.; Svenson, G.J. Family-level relationships of the spittlebugs and froghoppers (Hemiptera: Cicadomorpha: Cercopoidea). Syst. Entomol. 2010, 35, 393–415. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Ultrastructure and function of insect chemosensilla. Ann. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stall) and Nezara viridula L. (Hemiptera). Int. J. Insect Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Li, Q.L.; Wei, C. Antennal morphology of the cicada Meimuna mongolica (Distant) (Hemiptera: Cicadidae), with a deduction of its nymphal instars and discussion of the antennal morphological variations in relation to niche changes. Entomotaxonomia 2013, 2, 81–94. [Google Scholar]
- Catalá, S.S. Antennal sensilla of Triatominae (Hemiptera, Reduviidae): A comparative study of five genera. Int. J. Insect Morphol. Embryol. 1997, 26, 67–73. [Google Scholar] [CrossRef]
- Chinta, S.; Dickens, J.C.; Baker, G.T. Morphology and distribution of antennal sensilla of the tarnished plant bug, Lygus lineolaris (Palisot de beauvois) (Hemiptera: Miridae). Int. J. Insect Morphol. Embryol. 1997, 26, 21–26. [Google Scholar] [CrossRef]
- Gao, Z.Z.; Wu, W.J.; Liang, G.W. Observation of the antennal sensilla of Campylomma chinensis (Hemiptera: Miridae) by environmental scanning electron microscope. J. South China Agric. Univ. 2006, 27, 18–20. [Google Scholar]
- McIver, S.B. Structure of cuticle mechanoreceptors of arthropods. Ann. Rev. Entomol. 1975, 20, 381–397. [Google Scholar] [CrossRef]
- Sane, S.P.; Dieudonné, A.; Willism, M.A.; Daniel, T.L. Antennal mechanosensors mediate flight control in moths. Science 2007, 315, 863–866. [Google Scholar] [CrossRef]
- Slifer, E.H. The fine structure of arthropod chemoreceptors. Ann. Rev. Entomol. 1970, 15, 121–142. [Google Scholar] [CrossRef]
- Thurm, U. Ableitung der Rezeptor-potentiale und NervenimpulseeinzeIner Cuticula-Sensillen bei Insekten. Z. Naturforsch. 1962, 17, 285–286. [Google Scholar] [CrossRef]
- Keil, T.A. Morphology and Development of the Peripehral Olfactory Organs. In Insect Olfaction; Hansson, B.S., Ed.; Springer: Heidelberg, Germany, 1999; pp. 5–47. [Google Scholar]
- Altner, H.; Loftus, R. Ultrastructure and function of insect thermo- and hygroreceptors. Ann. Rev. Entomol. 1985, 30, 273–295. [Google Scholar] [CrossRef]
- Altner, H.; Schaller-Selzer, L.; Stetter, H.; Wohlrab, I. Poreless sensilla with inflexible sockets. A comparative study of a fundamental type of insect sensilla probably comprising thermo- and hygroreceptors. Cell Tissue Res. 1983, 234, 279–307. [Google Scholar] [CrossRef] [PubMed]
- Altner, H.; Tichy, H.; Altner, I. Lamellated outer dendritic segments of a sensory cell within a poreless thermo- and hygroreceptive sensillum of the insect Carausius morosus. Cell Tissue Res. 1978, 191, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Altner, H.; Routil, C.; Loftus, R. The structure of bimodal chemo-, thermo-, and hygroreceptive sensilla on the antenna of Locusta migratoria. Cell Tissue Res. 1981, 215, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Altner, H.; Prillinger, L. Ultrastructure of invertebrate chemo-, thermo-, andhygroreceptors and its functional significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar]
- Keil, T.A.; Steinbrecht, R.A. Mechanosensitive and olfactory sensilla of insects. In Insect Ultrastructure; King, R.C., Akai, H., Eds.; Plenum: New York, NY, USA, 1984; Volume 2, pp. 477–516. [Google Scholar]
- Steinbrecht, R.A. Olfactory receptors. In Atlas of Arthropod Sensory Receptors—Dynamic Morphology in Relation to Function; Eguchi, E., Tominaga, Y., Eds.; Springer: Tokyo, Japan, 1999; pp. 155–176. [Google Scholar]
- Fu, B.X.; Bellis, G.A.; Hong, J.; Wang, J.R.; Wu, Q.; Tang, Q.Y.; Cheng, J.A.; Zhu, Z.R. Morphology, distribution, and abundance of antennal sensilla of male and female macropterous and brachypterous small brown planthopper, Laodelphax striatellus (Fallén) (Hemiptera: Delphacidae). Microsc. Res. Tech. 2012, 75, 1492–1512. [Google Scholar] [CrossRef]
- Fu, B.X.; Zhu, Z.R.; Rong, N.H.; Hong, J.; Zhang, C.X.; Cheng, J.A. Phase-related developmental characteristics of antennal sensilla of nymphal Laodelphax striatellus (Hemiptera: Delphacidae), a serious virus-transmitting insect vector of graminaceous crops. Ann. Entomol. Soc. Am. 2013, 106, 626–636. [Google Scholar] [CrossRef]
- Bromley, A.K.; Dunn, J.A.; Anderson, M. Ultrastructure of the antennal sensilla of aphids. II. Trichoid, chordotonal and campaniform sensilla. Cell Tissue Res. 1980, 205, 493–511. [Google Scholar] [CrossRef]
- Keil, T.A. Functional morphology of insect mechanoreceptors. Microsc. Res. Tech. 1997, 39, 506–531. [Google Scholar] [CrossRef]
- Hallberg, E.; Hansson, B.; Steinbrecht, R.A. Morphological characteristic of antennal sensilla in the European cornborer Ostrinia nubilalis (Lepidoptera: Pyralidae). Tissue Cell 1994, 26, 489–502. [Google Scholar] [CrossRef]
- Pophof, B. Olfactory responses from sensilla coeloconica of the silkmoth Bombyx mori. Physiol. Entomol. 1997, 22, 239–248. [Google Scholar] [CrossRef]
- Pophof, B.; Stange, G.; Abrell, L. Volatile organic compounds as signals in a plantherbivore system: Electrophysiological responses in olfactory sensilla of the moth Cactoblastis cactorum. Chem. Senses 2005, 30, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Hunger, T.; Steinbrecht, R.A. Functional morphology of a double-walled multiporous olfactory sensillum: The sensillum coeloconicum of Bombyx mori (Insecta, Lepidoptera). Tissue Cell 1998, 30, 14–29. [Google Scholar] [CrossRef]
- Diehl, P.A.; Vlimant, M.; Guerenstein, P.; Guerin, P.M. Ultrastructure and receptor cell responses of the antennal grooved peg sensilla of Triatoma infestans (Hemiptera: Miridae). J. Insect Physiol. 2003, 41, 857–867. [Google Scholar] [CrossRef]
- Stacconi, M.V.R.; Romani, R. Antennal sensory structures in Scaphoideus titanus Ball (Hemiptera: Cicadellidae). Microsc. Res. Tech. 2012, 75, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T. Sexual communications in planthoppers. In The Rice Brown Planthopper; Kono, T., Ed.; FFTC/ASPAC: Taipei, Taiwan, 1977; pp. 84–94. [Google Scholar]
- Ichikawa, T.; Sakuma, M.; Ishii, S. Substrate vibrations: Mating signal of three species of planthoppers which attack the rice plant (Homoptera: Delphacidae). Appl. Entomol. Zool. 1975, 10, 162–171. [Google Scholar] [CrossRef]
- Obata, T.; Kim, M.; Koh, H.; Fukami, H. Planthopper attractant(s) in the rice plant. Jpn. J. Appl. Ent. Zool. 1981, 25, 47–51, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Chen, X.S.; Liang, A.P. Identification of a self-regulatory pheromone system that controls nymph aggregation behavior of rice spittlebug Callitettix versicolor. Front. Zool. 2015, 12, 1–12. [Google Scholar] [CrossRef]
- Cocroft, R.B. Offspring-parent communication in a subsocial treehopper (Hemiptera: Membracidae: Umbonia crassicornis). Behaviour 1998, 136, 1–21. [Google Scholar] [CrossRef]
- Hunt, R.E.; Fox, J.P.; Haynes, K.F. Behavioral response of Graminella nigrifrons (Homoptera: Cicadellidae) to experimentally manipulated vibrational signals. J. Insect Behav. 1992, 5, 1–13. [Google Scholar] [CrossRef]
- Hunt, R.E. Vibrational signals associated with mating behaviorin the treehopper. Enchenopa binotata Say (Hemiptera: Homoptera: Membracidae). J. N. Y. Entomol. Soc. 1994, 102, 266–270. [Google Scholar]
- Lopez Machado, F.E.; Peck, D.C.; Montoya Lerma, J. The importance of vibrational communication in the reproductive behavior of grassland spittlebugs (Homoptera: Cercopidae). Rev. Colomb. Entomol. 2001, 27, 9–15. [Google Scholar]
- Mazzoni, V.; Lucchi, A.; Ioriatti, C.; Virant-Doberlet, M.; Anfora, G. Mating behavior of Hyalesthes obsoletus (Hemiptera: Cixiidae). Ann. Entomol. Soc. Am. 2010, 103, 813–822. [Google Scholar] [CrossRef]
- Tishechkin, D.Y. Acoustic signalling in Paralimnini leafhoppers (Homoptera: Cicadellidae: Deltacephalinae). Zool. Zhurnal 1992, 71, 58–65. [Google Scholar]
- Virant-Doberlet, M.; Zezlina, I. Vibrational communication of Metcalfa pruinosa (Hemiptera: Fulgoroidea: Flatidae). Ann. Entomol. Soc. Am. 2007, 100, 73–82. [Google Scholar] [CrossRef]
- Zhantiev, R.D.; Tishechkin, D.Y. Acoustic signalling of cicadellids of the genus Macropsis Lew. (Homoptera, Cicadellidae) of the European part of the USSR. Entomol. Oboz. 1989, 68, 469–477. [Google Scholar]
- Mazzoni, V.; Ioriatti, C.; Trona, F.; Lucchi, A.; De Cristofaro, A.; Anfora, G. Study on the role of olfaction in host plant detection of Scaphoideus titanus (Hemiptera: Cicadellidae) nymphs. J. Econ. Entomol. 2009, 102, 974–980. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensilla Type | Distribution | Length (μm) | External Diameter (μm) | N | Amount |
|---|---|---|---|---|---|
| Sensilla trichodea | Scape and pedicel | 56.33 ± 3.85 | 2.71 ± 0.18 | 7 | 24–30 |
| Sensilla basiconica 1 | Scape | 11.96 ± 1.12 | 2.32 ± 0.07 | 7 | 6–7 |
| Sensilla basiconica 2 | Flagellum | 57.08 ± 2.84 (♀) 72.10 ± 2.33 (♂) | 20.37 ± 0.99 18.49 ± 0.63 | 5 | 1 |
| Sensilla campaniformia 1 | Scape and pedicel | NA | 4.67 ± 0.56 | 5 | 1 |
| Sensilla campaniformia 2 | Tip of pedicel | NA | 5.91 ± 0.75 | 5 | 1 |
| Sensilla campaniformia 3 | Tip of pedicel | NA | 12.55 ± 1.01 | 5 | 1 |
| Sensilla coeloconica 1 | Pedicel | NA | 2.17 ± 0.36 | 8 | 13 |
| Sensilla coeloconica 2 | Flagellum | NA | 4.37 ± 0.58 | 8(♀) 8(♂) | 3–4 4–6 |
| Sensilla coeloconica 3 | Flagellum | NA | 8.03 ± 1.02 | 8(♀) 8(♂) | 16–18 100–106 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Wu, N.; Brożek, J.; Dai, W. Antennal Morphology and Sexual Dimorphism of Antennal Sensilla in Callitettix versicolor (Fabricius) (Hemiptera: Cercopidae). Insects 2019, 10, 56. https://doi.org/10.3390/insects10020056
Zhu Q, Wu N, Brożek J, Dai W. Antennal Morphology and Sexual Dimorphism of Antennal Sensilla in Callitettix versicolor (Fabricius) (Hemiptera: Cercopidae). Insects. 2019; 10(2):56. https://doi.org/10.3390/insects10020056
Chicago/Turabian StyleZhu, Qing, Nan Wu, Jolanta Brożek, and Wu Dai. 2019. "Antennal Morphology and Sexual Dimorphism of Antennal Sensilla in Callitettix versicolor (Fabricius) (Hemiptera: Cercopidae)" Insects 10, no. 2: 56. https://doi.org/10.3390/insects10020056