Proteomic Analysis of the Venom from the Ruby Ant Myrmica rubra and the Isolation of a Novel Insecticidal Decapeptide

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ant Collection
2.2. Taxonomy
2.3. Venom Preparation
2.4. Disulfide Bond Reduction and Alkylation
2.5. RP-HPLC Analysis of Venom Samples and Subsequent Fractionation
2.6. Peptide Purification and Enrichment
2.7. Edman Degradation
2.8. Peptide Synthesis
2.9. RP-HPLC of Natural and Synthetic Peptide
2.10. Analysis of Crude Venom and Fractions by Mass Spectrometry
2.11. Antimicrobial Assay
2.12. Maintenance of Aphids and Feeding Assays with the M. rubra Peptide
2.13. Insecticides and Aphid Bioassays
2.14. Data Analysis
3. Results
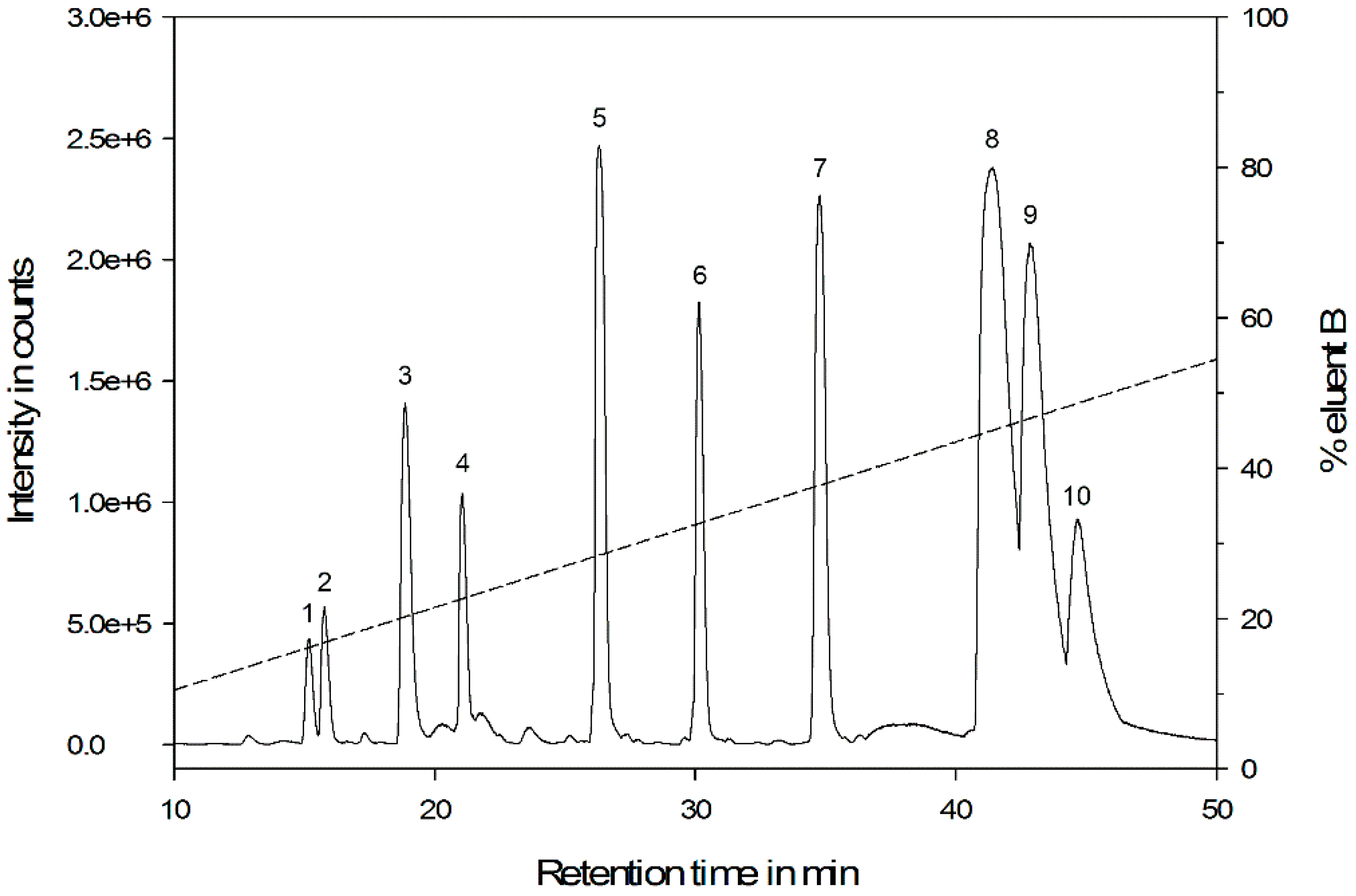
3.1. General Venom Characteristics
3.2. Reduction and Alkylation
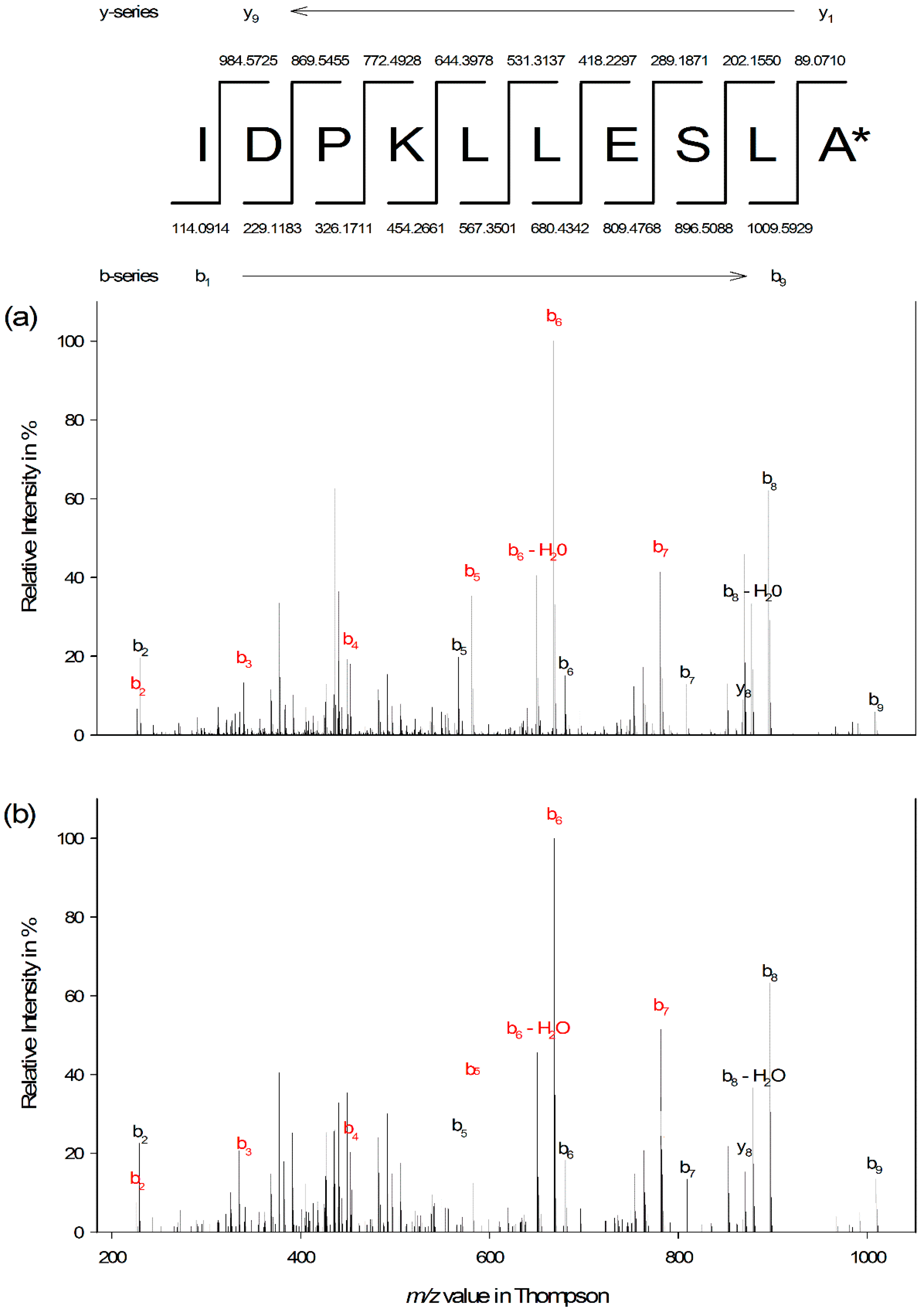
3.3. Edman Degradation and Peptide De Novo Sequencing
3.4. Effect of Peptide on Bacterial Strains
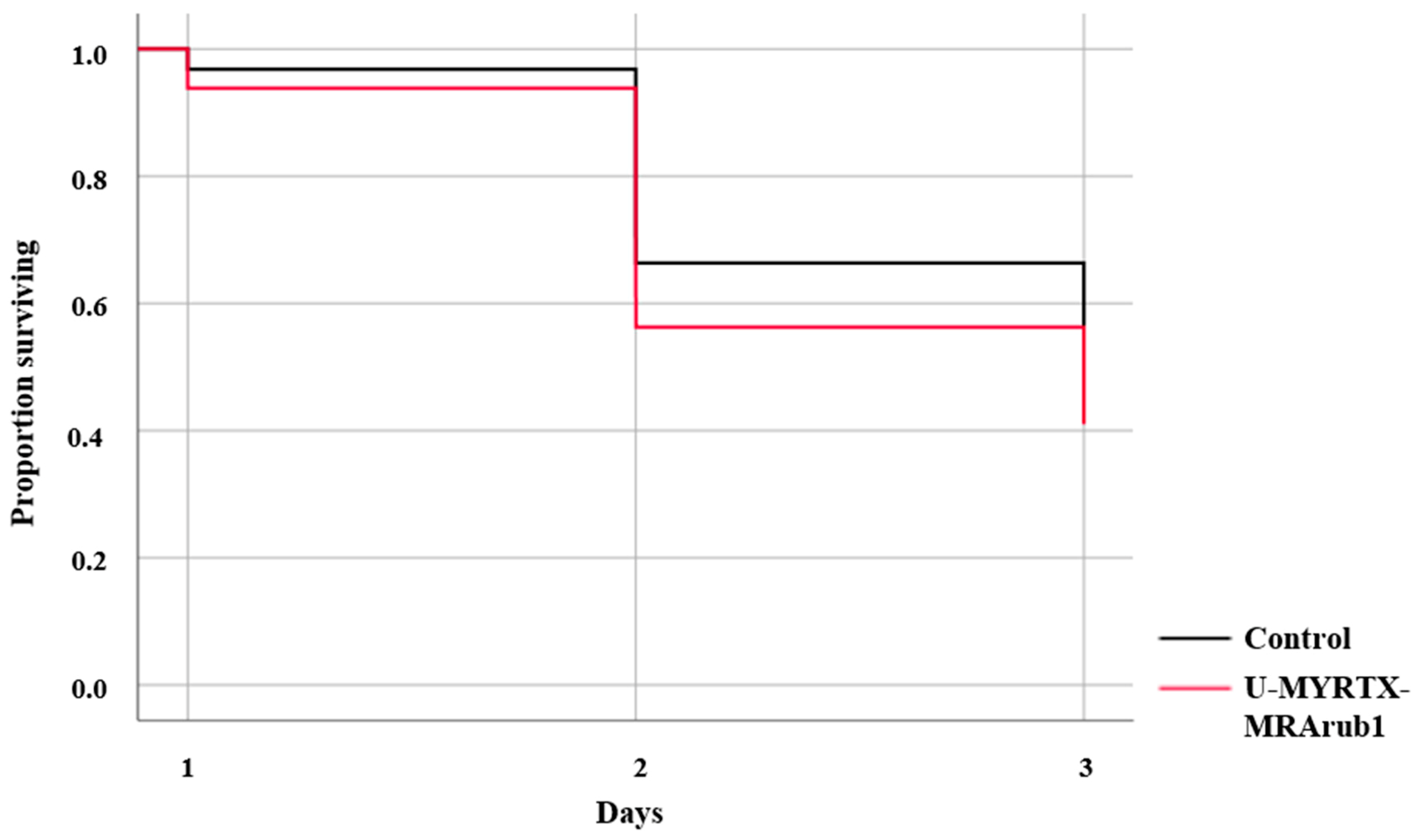
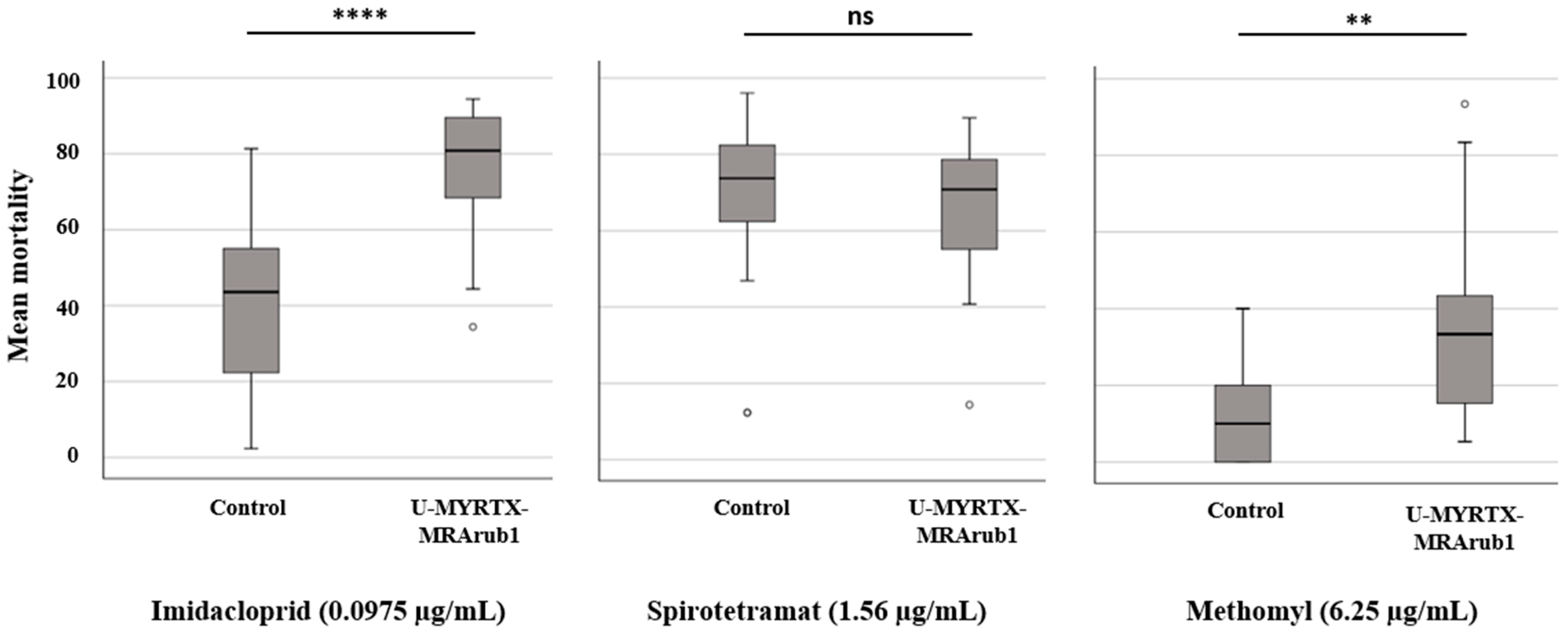
3.5. Effect of Peptide Treatment on A. pisum Susceptibility to Chemical Insecticides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- AntCat. An Online Catalog of the Ants of the World. Available online: http://antcat.org (accessed on 1 December 2018).
- AntWeb. Available online: https://www.antweb.org (accessed on 1 December 2018).
- Touchard, A.; Aili, R.S.; Fox, G.E.; Escoubas, P.; Orivel, J.; Nicholson, M.G.; Dejean, A. The biochemical toxin arsenal from ant venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A.B. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Téné, N.; Song, P.C.T.; Lefranc, B.; Leprince, J.; Treilhou, M.; Bonnafé, E. Deciphering the Molecular Diversity of an Ant Venom Peptidome through a Venomics Approach. J. Proteome Res. 2018, 17, 3503–3516. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis Part 1: Causes and threats. P&T Comm. 2015, 40, 277–283. [Google Scholar]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef]
- Keymanesh, K.; Soltani, S.; Sardari, S. Application of antimicrobial peptides in agriculture and food industry. World J. Microbiol. Biotechnol. 2009, 25, 933–944. [Google Scholar] [CrossRef]
- Orivel, J.; Redeker, V.; Le Caer, J.P.; Krier, F.; Revol-Junelles, A.M.; Longeon, A.; Chaffotte, A.; Dejean, A.; Rossier, J. Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii. J. Biol. Chem. 2001, 276, 17823–17829. [Google Scholar] [CrossRef]
- Schmidt, C.A.; Shattuck, S.O. The Higher Classification of the Ant Subfamily Ponerinae (Hymenoptera: Formicidae), with a Review of Ponerine Ecology and Behavior. Zootaxa 2014, 3817. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Copello, J.A.; Evans, M.S.; Suarez, A.V. A biochemical characterization of the major peptides from the Venom of the giant Neotropical hunting ant Dinoponera australis. Toxicon 2010, 55, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Rifflet, A.; Gavalda, S.; Tene, N.; Orivel, J.; Leprince, J.; Guilhaudis, L.; Genin, E.; Vetillard, A.; Treilhou, M. Identification and characterization of a novel antimicrobial peptide from the venom of the ant Tetramorium bicarinatum. Peptides 2012, 38, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.B.; Torres, A.F.C.; Mello, C.P.; de Menezes, R.R.P.P.B.; Sampaio, T.L.; Canuto, J.A.; da Silva, J.J.A.; Freire, V.N.; Quinet, Y.P.; Havt, A.; et al. Antimicrobial effect of Dinoponera quadriceps (Hymenoptera: Formicidae) venom against Staphylococcus aureus strains. J. Appl. Microbiol. 2014, 117, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Schluns, H.; Crozier, R.H. Molecular and chemical immune defenses in ants (Hymenoptera: Formicidae). Myrmecol. News 2009, 12, 237–249. [Google Scholar]
- Pluzhnikov, K.A.; Kozlov, S.A.; Vassilevski, A.A.; Vorontsova, O.V.; Feofanov, A.V.; Grishin, E.V. Linear antimicrobial peptides from Ectatomma quadridens ant venom. Biochimie 2014, 107 (Pt B), 211–215. [Google Scholar] [CrossRef]
- Aili, S.R.; Touchard, A.; Petitclerc, F.; Dejean, A.; Orivel, J.; Padula, M.P.; Escoubas, P.; Nicholson, G.M. Combined peptidomic and proteomic analysis of electrically stimulated and manually dissected venom from the South American bullet ant Paraponera clavata. J. Proteome Res. 2017, 16, 1339–1351. [Google Scholar] [CrossRef]
- Szolajska, E.; Poznanski, J.; Ferber, M.L.; Michalik, J.; Gout, E.; Fender, P.; Bailly, I.; Dublet, B.; Chroboczek, J. Poneratoxin, a neurotoxin from ant venom. Eur. J. Biochem. 2004, 271, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Piek, T.; Duval, A.; Hue, B.; Karst, H.; Lapied, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Schmidt, J.O. Poneratoxin, a novel peptide neurotoxin from the venom of the ant, Paraponera clavata. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 99, 487–495. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Terumi, N.; Schmidt, J.O. Pharmacological characterization and chemical fractionation of the venom of the ponerine ant, Paraponera clavata (F.). Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 99, 481–486. [Google Scholar] [CrossRef]
- Touchard, A.; Brust, A.; Cardoso, F.C.; Chin, Y.K.; Herzig, V.; Jin, A.H.; Dejean, A.; Alewood, P.F.; King, G.F.; Orivel, J.; et al. Isolation and characterization of a structurally unique beta-hairpin venom peptide from the predatory ant Anochetus emarginatus. Biochim. Biophys. Acta 2016, 1860, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J.-M. Diversity of folds in animal toxins acting on ion channels. Biochem. J. 2004, 378, 717. [Google Scholar] [CrossRef] [Green Version]
- Zeitler, B.; Herrera Diaz, A.; Dangel, A.; Thellmann, M.; Meyer, H.; Sattler, M.; Lindermayr, C. De-novo design of antimicrobial peptides for plant protection. PLoS ONE 2013, 8, e71687. [Google Scholar] [CrossRef]
- Montesinos, E. Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett. 2007, 270, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Will, T.; Vilcinskas, A. Aphid-proof plants: Biotechnology-based approaches for aphid control. Adv. Biochem. Eng./Biotechnol. 2013, 136, 179–203. [Google Scholar] [CrossRef]
- Luna-Ramirez, K.; Skaljac, M.; Grotmann, J.; Kirfel, P.; Vilcinskas, A. Orally delivered scorpion antimicrobial peptides exhibit activity against Pea Aphid (Acyrthosiphon pisum) and its bacterial symbionts. Toxins 2017, 9, 261. [Google Scholar] [CrossRef]
- Pal, N.; Yamamoto, T.; King, G.F.; Waine, C.; Bonning, B. Aphicidal efficacy of scorpion- and spider-derived neurotoxins. Toxicon 2013, 70, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Zhang, Y.-J.; Zhai, X.-M.; Zhao, J.-J.; Peng, D.-L.; Wu, G. Expression of a scorpion toxin gene BmKit enhances the virulence of Lecanicillium lecanii against aphids. J. Pest Sci. 2015, 88, 637–644. [Google Scholar] [CrossRef]
- Yang, S.; Fitches, E.; Pyati, P.; Gatehouse, J.A. Effect of insecticidal fusion proteins containing spider toxins targeting sodium and calcium ion channels on pyrethroid-resistant strains of peach-potato aphid (Myzus persicae). Pest Manag. Sci. 2015, 71, 951–956. [Google Scholar] [CrossRef]
- Bonning, B.C.; Pal, N.; Liu, S.; Wang, Z.; Sivakumar, S.; Dixon, P.M.; King, G.F.; Miller, W.A. Toxin delivery by the coat protein of an aphid-vectored plant virus provides plant resistance to aphids. Nat. Biotechnol. 2013, 32, 102. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Barzman, M.; Barberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015. [Google Scholar] [CrossRef]
- Klatt, B.K.; Rundlöf, M.; Smith, H.G. Maintaining the restriction on neonicotinoids in the European Union—Benefits and risks to bees and pollination services. Front. Ecol. Evol. 2016, 4, 4. [Google Scholar] [CrossRef]
- Wetterer, J.K.; Radchenko, A.G. Worldwide spread of the ruby ant, Myrmica rubra (Hymenoptera: Formicidae). Myrmecol. News 2011, 14, 87–96. [Google Scholar]
- Arevalo, H.A.; Groden, E. European Fire Ant, Red Ant (Suggested Common Names), Myrmica rubra Linnaeus (Insecta: Hymenoptera: Formicidae: Myrmicinae); University of Florida: Gainesville, FL, USA, 2007. [Google Scholar]
- McPhee, K.; Garnas, J.; Drummond, F.; Groden, E. Homopterans and an Invasive Red Ant, Myrmica rubra (L.), in Maine. Environ. Entomol. 2012, 41, 59–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostás, M.; Blassmann, K. Insects had it first: Surfactants as a defence against predators. Proc. R. Soc. B Biol. Sci. 2008, 276, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Verble-Pearson, R.; Pearson, S. European Fire Ant Presence Decreases Native Arboreal Insect Abundance in Acadia National Park, Maine, USA. Nat. Areas J. 2016, 36, 162–166. [Google Scholar] [CrossRef]
- Chen, W.; Adams, E.S. The Distribution and Habitat Affinities of the Invasive Ant Myrmica rubra (Hymenoptera: Formicidae) in Southern New England. Environ. Entomol. 2018, 47, 527–534. [Google Scholar] [CrossRef]
- Naumann, K.; Higgins, R.J. The European fire ant (Hymenoptera: Formicidae) as an invasive species: Impact on local ant species and other epigaeic arthropods. Can. Entomol. 2014, 147, 592–601. [Google Scholar] [CrossRef]
- Evershed, R.P.; Morgan, E.D.; Cammaerts, M.C. 3-ethyl-2,5-dimethylpyrazine, the trail pheromone from the venom gland of eight species of Myrmica ants. Insect Biochem. 1982, 12, 383–391. [Google Scholar] [CrossRef]
- Seifert, B. Die Ameisen Mittel- und Nordeuropas; lutra: Goerlitz, Germany, 2007; ISBN 978-3-936412-03-1. [Google Scholar]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Schmidtberg, H.; Skaljac, M.; Vilcinskas, A. Heat shock protein 83 plays pleiotropic roles in embryogenesis, longevity, and fecundity of the pea aphid Acyrthosiphon pisum. Dev. Genes Evol. 2017, 227, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Vilcinskas, A. The structural sheath protein of aphids is required for phloem feeding. Insect Biochem. Mol. Biol. 2015, 57, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, A.; Van Damme, E.J.M.; Smagghe, G. Evaluation of the susceptibility of the pea aphid, Acyrthosiphon pisum, to a selection of novel biorational insecticides using an artificial diet. J. Insect Sci. 2009, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Febvay, G.; Delobel, B.; Rahbé, Y. Influence of the amino acid balance on the improvement of an artificial diet for a biotype of Acyrthosiphon pisum (Homoptera: Aphididae). Can. J. Zool. 1988, 6611, 2449–2453. [Google Scholar] [CrossRef]
- IRAC—Insecticide Resistance Action Committee. Sucking Pests MoA Poster. Available online: http://www.irac-online.org/documents/sucking-pests-moa-poster/ (accessed on 1 December 2018).
- Skaljac, M.; Kirfel, P.; Grotmann, J.; Vilcinskas, A. Fitness costs of infection with Serratia symbotica are associated with greater susceptibility to insecticides in the pea aphid Acyrthosiphon pisum. Pest Manag. Sci. 2018, 74, 1829–1836. [Google Scholar] [CrossRef]
- IRAC—Insecticide Resistance Action Committee. Susceptibility Test Method 019. Available online: https://www.irac-online.org/methods/aphids-adultnymphs/ (accessed on 1 December 2018).
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Senko, M.W.; Beu, S.C.; McLafferty, F.W. Determination of monoisotopic masses and ion populations for large biomolecules from resolved isotopic distributions. J. Am. Soc. Mass. Spectrom. 1995, 6, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Heep, J.; Bioresources Research Group, Fraunhofer Institute for Molecular Biology and Applied Ecology, Gießen, Germany. Unpublished data. 2018.
- Cologna, C.T.; Cardoso, J.d.S.; Jourdan, E.; Degueldre, M.; Upert, G.; Gilles, N.; Uetanabaro, A.P.T.; Costa Neto, E.M.; Thonart, P.; de Pauw, E.; et al. Peptidomic comparison and characterization of the major components of the venom of the giant ant Dinoponera quadriceps collected in four different areas of Brazil. J. Proteom. 2013, 94, 413–422. [Google Scholar] [CrossRef]
- Touchard, A.; Koh, J.M.; Aili, S.R.; Dejean, A.; Nicholson, G.M.; Orivel, J.; Escoubas, P. The complexity and structural diversity of ant venom peptidomes is revealed by mass spectrometry profiling. Rapid Commun. Mass Spectrom. 2015, 29, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Rivier, J.; Clark, C.; Ramilo, C.A.; Corpuz, G.P.; Abogadie, F.C.; Mena, E.E.; Woodward, S.R.; Hillyard, D.R.; Cruz, L.J. Diversity of conus neuropeptides. Science 1990, 249, 257. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Čeřovský, V.; Hovorka, O.; Cvačka, J.; Voburka, Z.; Bednárová, L.; Borovičková, L.; Slaninová, J.; Fučík, V. Melectin: A novel antimicrobial peptide from the venom of the cleptoparasitic bee Melecta albifrons. ChemBioChem 2008, 9, 2815–2821. [Google Scholar] [CrossRef] [PubMed]
- Tincu, J.A.; Taylor, S.W. Antimicrobial peptides from marine invertebrates. Antimicrob. Agents Chemother. 2004, 48, 3645. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.C.C.; Zanotta, L.C.; Kyaw, C.M.; Schwartz, E.N.F.; Schwartz, C.A.; Sebben, A.; Sousa, M.V.; Fontes, W.; Castro, M.S. Ocellatins: New antimicrobial peptides from the skin secretion of the South American frog Leptodactylus ocellatus (Anura: Leptodactylidae). Protein J. 2004, 23, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Cao, L.; He, Y.; Hu, J.; Di, Z.; Wu, Y.; Li, W.; Cao, Z. Ctriporin, a new anti-methicillin-resistant Staphylococcus aureus Peptide from the Venom of the Scorpion Chaerilus tricostatus. Antimicrob. Agents Chemother. 2011, 55, 5220. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Novotny, M.J.; Morris, W.L.; Tang, Y.Q.; Smith, W.; Cullor, J.S. Indolicidin, a novel bactericidal tridecapeptide amide from neutrophils. J. Biol. Chem. 1992, 267, 4292–4295. [Google Scholar]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Luna-Ramirez, K.; Tonk, M.; Rahnamaeian, M.; Vilcinskas, A. Bioactivity of Natural and Engineered Antimicrobial Peptides from Venom of the Scorpions Urodacus yaschenkoi and U. manicatus. Toxins 2017, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Y.; Lee, W.-H.; Zhang, Y. Antimicrobial peptides from the venom gland of the social wasp Vespa tropica. Toxicon 2013, 74, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European Red Frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [PubMed]
- Zelezetsky, I.; Tossi, A. Alpha-helical antimicrobial peptides—Using a sequence template to guide structure–activity relationship studies. Biochim. Biophys. Acta 2006, 1758, 1436–1449. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Rinaldi, A.C.; Giulio, A.D.; Mignogna, G.; Bozzi, A.; Barra, D.; Simmaco, M. Structure–function relationships of temporins, small antimicrobialpeptides from amphibian skin. Eur. J. Biochem. 2000, 267, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Yamazaki, K.; Kawakami, S.; Miyoshi, D.; Ooi, T.; Hashimoto, S.; Taguchi, S. In vivo target exploration of apidaecin based on Acquired Resistance induced by Gene Overexpression (ARGO assay). Sci. Rep. 2017, 7, 12136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, J.F.; Muñoz, A.; Pérez-Payá, E.; Misra, S.; López-García, B. Identification and Rational Design of Novel Antimicrobial Peptides for Plant Protection. Annu. Rev. Phytopathol. 2008, 46, 273–301. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | MWcrude (Da) | MWred. (Da) | MWalk. (Da) | RA (%) | aa Length a | S-S |
|---|---|---|---|---|---|---|---|
| 1 | 15.17 | 544.2524 | 544.2518 | 544.2525 | 18 | 5 | 0 |
| 2 | 15.77 | 1100.6561 | 1100.6547 | 1100.6544 | 23 | 10 | 0 |
| 3 | 18.84 | 1463.8353 | 1465.8501 | 1579.8809 | 57 | 13 | 1 |
| 4 | 21.05 | 1401.7441 | 1401.7418 | 1401.7336 | 42 | 13 | 0 |
| 5 | 26.30 | 1096.6506 | 1096.6470 | 1096.6487 | 100 | 10 | 0 |
| 6 | 30.15 | 1112.6765 | 1112.6713 | 1112.6750 | 74 | 10 | 0 |
| 7 | 34.78 | 1590.8308 | 1592.8418 | 1706.8684 | 92 | 14 | 1 |
| 8 | 41.41 | 2477.4727 | 2477.4578 | 2477.4604 | 41 | 23 | 0 |
| 9 | 42.83 | 2837.5846 | 2837.5700 | 2837.5726 | 84 | 26 | 0 |
| 10 | 44.66 | 2525.4244 | 2525.4101 | 2525.4146 | 38 | 23 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heep, J.; Klaus, A.; Kessel, T.; Seip, M.; Vilcinskas, A.; Skaljac, M. Proteomic Analysis of the Venom from the Ruby Ant Myrmica rubra and the Isolation of a Novel Insecticidal Decapeptide. Insects 2019, 10, 42. https://doi.org/10.3390/insects10020042
Heep J, Klaus A, Kessel T, Seip M, Vilcinskas A, Skaljac M. Proteomic Analysis of the Venom from the Ruby Ant Myrmica rubra and the Isolation of a Novel Insecticidal Decapeptide. Insects. 2019; 10(2):42. https://doi.org/10.3390/insects10020042
Chicago/Turabian StyleHeep, John, Alica Klaus, Tobias Kessel, Maximilian Seip, Andreas Vilcinskas, and Marisa Skaljac. 2019. "Proteomic Analysis of the Venom from the Ruby Ant Myrmica rubra and the Isolation of a Novel Insecticidal Decapeptide" Insects 10, no. 2: 42. https://doi.org/10.3390/insects10020042