Context-Dependence and the Development of Push-Pull Approaches for Integrated Management of Drosophila suzukii


{kind=link}
Abstract
:1. Introduction
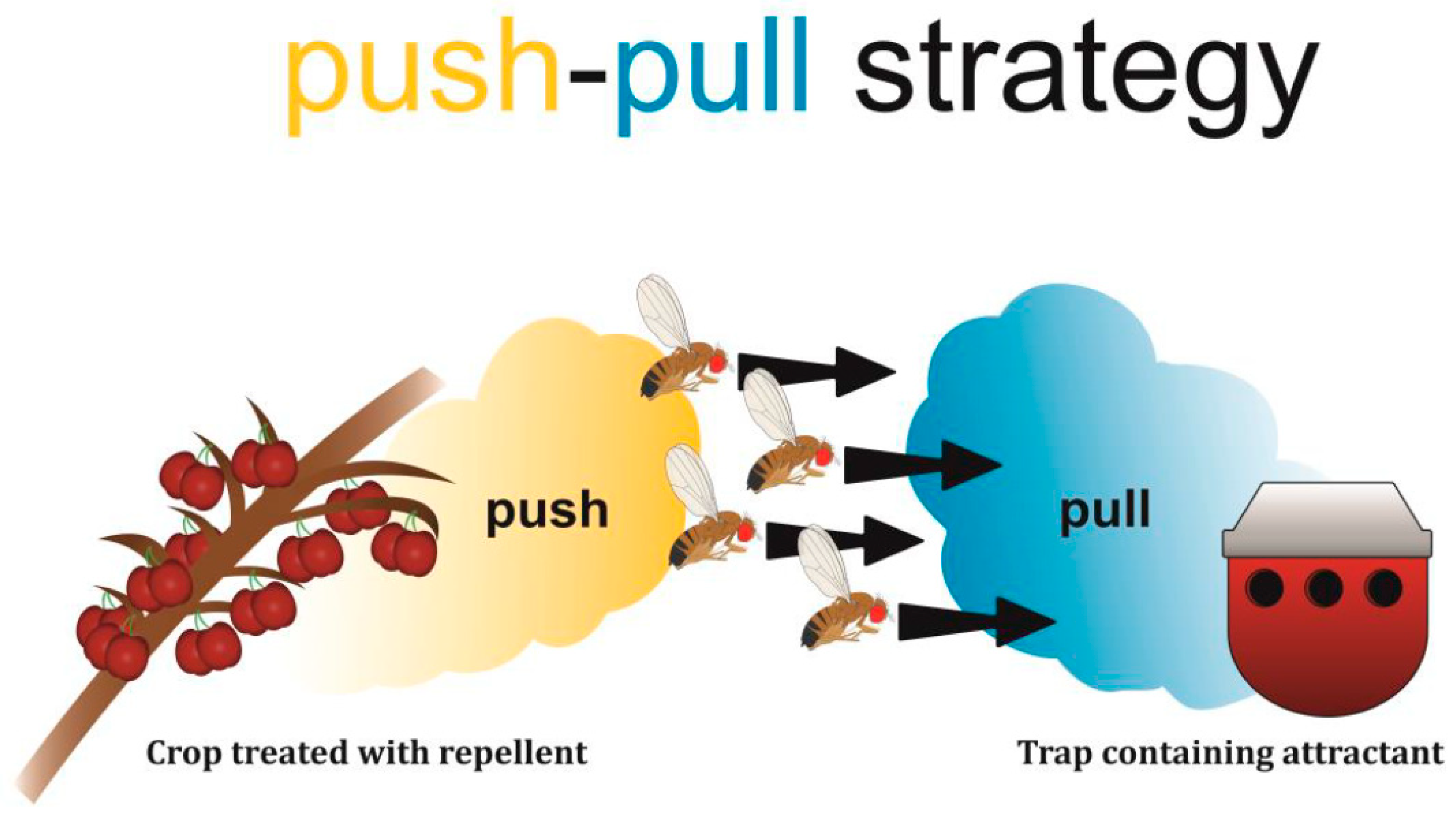
1.1. Push-Pull Approach as Part of Integrated Pest Management (IPM)
1.2. Fundamental Ecological Knowledge Required to Design Effective Push-Pull Systems
1.3. Invasive Pest Drosophila suzukii
2. Infochemicals
3. Context-Dependence
4. Drosophila suzukii—A New Invasive Pest
5. Chemical Ecology of Drosophila
5.1. Drosophila suzukii
5.2. Context Dependence in D. suzukii
6. What Is Needed for Development of Successful IPM of D. suzukii?
6.1. Push-Pull
6.2. Additional Density Reduction Methods
6.3. Additional Methods Under Investigation
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Prokopy, R.J.; Kogan, M. Integrated Pest Management. In Encyclopedia of Insects, 2nd ed.; Resh, V., Carde, R.T., Eds.; Academic Press: Oxford, UK, 2009; pp. 523–528. [Google Scholar]
- Radcliffe, E.B.; Hutchison, W.D.; Cancelado, R.E. Integrated Pest Management. Concepts, Tactics, Strategies and Case Studies; Cambridge University Press: Cambridge, UK, 2009; p. 529. [Google Scholar]
- Pimentel, D. Perspectives of integrated pest management. Crop Prot. 1982, 1, 5–26. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.O.; Pittchar, J.O.; Murage, A.W.; Birkett, M.A.; Bruce, T.J.A.; Pickett, J.A. Achieving food security for one million sub-Saharan African poor through push-pull innovation by 2020. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20120284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bale, J.S.; van Lenteren, J.C.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 761–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickett, J.A.; Woodcock, C.M.; Midega, C.A.O.; Khan, Z.R. Push-pull farming systems. Curr. Opin. Biotechnol. 2014, 26, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.R.; Cowles, R.S. Stimulo-deterrent diversion: A concept and its possible application to onion maggot control. J. Chem. Ecol. 1990, 16, 3197–3212. [Google Scholar] [CrossRef]
- Shelton, A.M.; Badenes-Perez, E. Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.A.; Bezerra-Silva, G.C.D.; Mastrangelo, T. The host marking pheromone application on the management of fruit flies—A review. Braz. Arch. Biol. Technol. 2012, 55, 835–842. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.M.; Suckling, D.M.; Byers, J.A.; Jang, E.B.; Wearing, C.H. Potential of “lure and kill” in long-term pest management and eradication of invasive species. J. Econ. Entomol. 2009, 102, 815–835. [Google Scholar] [CrossRef]
- Katsoyannos, B.I.; Boller, E.F. Second field application of oviposition-deterring pheromone of the European cherry fruit fly, Rhagoletis cerasi L. (Diptera: Tephritidae). J. Appl. Entomol. 1980, 89, 278–281. [Google Scholar] [CrossRef]
- Aluja, M.; Boller, E.F. Host-marking pheromone of Rhagoletis cerasi: Field deployment of synthetic pheromone as a novel cherry fly management strategy. Entomol. Exp. Appl. 1992, 65, 141–147. [Google Scholar] [CrossRef]
- Aluja, M.; Boller, E.F. Host marking pheromone of Rhagoletis cerasi: Foraging behavior in response to synthetic pheromonal isomers. J. Chem. Ecol. 1992, 18, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in Insect Olfaction: To Smell or Not to Smell? Annu. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Lemmen-Lechelt, J.K.; Wist, T.J.; Evenden, M.L. State-dependent plasticity in response to host-plant volatiles in a long-lived moth, Caloptilia fraxinella (Lepidoptera: Gracillariidae). J. Chem. Ecol. 2018, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.R.; Strickler, K.L. Finding and accepting host plants. In Chemical Ecology of Insects; Bell, W.J., Cardé, R.T., Eds.; Chapman & Hall: London, UK, 1984; pp. 127–157. [Google Scholar]
- Dicke, M.; Sabelis, M.W. Infochemical terminology: Based on cost-benefit analysis rather than origin of compounds? Funct. Ecol. 1988, 2, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Nordlund, D.A.; Lewis, W.J. Terminology of chemical releasing stimuli in intraspecific and interspecific interactions. J. Chem. Ecol. 1976, 2, 211–220. [Google Scholar] [CrossRef]
- Städler, E. Contact chemoreception. In Chemical Ecology of Insects; Bell, W.J., Cardé, R.T., Eds.; Chapman and Hall: London, UK, 1984; pp. 3–36. [Google Scholar]
- Elkinton, J.S.; Carde, R.T. Odor Dispersion. In Chemical Ecology of Insects, 1st ed.; Bell, W.J., Carde, R.T., Eds.; Chapman and Hall LtD: London, UK, 1984; pp. 37–70. [Google Scholar]
- Wajnberg, E.; Colazza, S. Chemical Ecology of Insect Parasitoids; John Wiley & Sons, Ltd.: Chichester, UK, 2013. [Google Scholar]
- Raguso, R.A.; Agrawal, A.A.; Douglas, A.E.; Jander, G.; Kessler, A.; Poveda, K.; Thaler, J.S. The raison d’etre of chemical ecology. Ecology 2015, 96, 617–630. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Sabelis, M.W. Costs and benefits of chemical information conveyance: Proximate and ultimate factors. In Insect Chemical Ecology: An Evolutionary Approach; Roitberg, B.D., Isman, M.B., Eds.; Chapman & Hall: New York, NY, USA, 1992; pp. 122–155. [Google Scholar]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Raguso, R.A. Wake Up and Smell the Roses: The Ecology and Evolution of Floral Scent. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 549–569. [Google Scholar] [CrossRef]
- Blum, M.S. Alarm Pheromonoes. In Behaviour, 1st ed.; Brooks, G.T., Ed.; Pergamon Press Ltd.: Oxford, UK, 1985; Volume 9, pp. 193–224. [Google Scholar]
- Tamaki, Y. Sex Pheromones. In Behaviour, 1st ed.; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press Ltd.: Oxford, UK, 1985; Volume 9, pp. 145–192. [Google Scholar]
- Prokopy, R.J. Visual and olfactory stimulus interaction in resoure finding by insects. In Mechanisms of Insect Olfaction, 1st ed.; Payne, T.L., Birch, M.C., Kennedy, C.E.J., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 81–90. [Google Scholar]
- Carde, R.T.; Baker, T.C. Sexual communication with pheromones. In Chemical Ecology of Insects, 1st ed.; Bell, W.J., Carde, R.T., Eds.; Chapman and Hall Ltd.: Oxford, UK, 1984; pp. 355–386. [Google Scholar]
- Bartelt, R.J.; Schaner, A.M.; Jackson, L.L. cis-Vaccenyl acetate as an aggregation pheromone in Drosophila melanogaster. J. Chem. Ecol. 1985, 11, 1747–1756. [Google Scholar] [CrossRef]
- Borden, J.H. Aggregation Pheromones. In Behaviour, 1st ed.; Brooks, G.T., Ed.; Pergamon Press Ltd.: Oxford, UK, 1985; Volume 9, pp. 257–285. [Google Scholar]
- Dumenil, C.; Woud, D.; Pinto, F.; Alkema, J.T.; Jansen, I.; Van der Geest, A.M.; Roessingh, S.; Billeter, J.C. Pheromonal Cues Deposited by Mated Females Convey Social Information about Egg-Laying Sites in Drosophila Melanogaster. J. Chem. Ecol. 2016, 42, 259–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardé, R.T.; Mafra-Neto, A. Insect Pheromone Research: New Directions; Cardé, R.T., Minks, A.K., Eds.; Chapman & Hall: New York, NY, USA, 1997; p. 684. [Google Scholar]
- Boppré, M. Insects pharmacophagously utilizing defensive plant chemicals (Pyrrolizidine Alkaloids). Naturwissenschaften 1986, 73, 17–26. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones. Curr. Biol. 2017, 27, R739–R743. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, B.; van Baalen, E.J.A.; Dicke, M.; Vet, L.E.M. Pheromone-mediated aggregation in nonsocial arthropods: An evolutionary ecological perspective. Annu. Rev. Entomol. 2005, 50, 321–346. [Google Scholar] [CrossRef]
- Nufio, C.R.; Papaj, D.R. Host marking behavior in phytophagous insects and parasitoids. Entomol. Exp. Appl. 2001, 99, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Roitberg, B.D.; Cairl, R.S.; Prokopy, R.J. Oviposition deterring pheromone influences dispersal distance in tephritid fruit flies. Entomol. Exp. Appl. 1984, 35, 217–220. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Roitberg, B.D. Joining and avoidance behavior in nonsocial insects. Annu. Rev. Entomol. 2001, 46, 631–665. [Google Scholar] [CrossRef]
- Wertheim, B. Evolutionary ecology of communication signals that induce aggregative behaviour. Oikos 2005, 109, 117–124. [Google Scholar] [CrossRef]
- Nadel, R.L.; Wingfield, M.J.; Scholes, M.C.; Lawson, S.A.; Slippers, B. The potential for monitoring and control of insect pests in Southern Hemisphere forestry plantations using semiochemicals. Ann. For. Sci. 2012, 69, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Sarles, L.; Verhaeghe, A.; Francis, F.; Verheggen, F.J. Semiochemicals of Rhagoletis fruit flies: Potential for integrated pest management. Crop Prot. 2015, 78, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Stelinski, L.; Gut, L.; Thomson, D. Codling moth management and chemical ecology. Annu. Rev. Entomol. 2008, 53, 503–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, J.S. The concepts of olfactory ‘arrestment’ and ‘attraction’. Physiol. Entomol. 1978, 3, 91–98. [Google Scholar] [CrossRef]
- Smid, H.M.; Vet, L.E.M. The complexity of learning, memory and neural processes in an evolutionary ecological context. Curr. Opin. Insect Sci. 2016, 15, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, D.S.; Demandt, N.; Alkema, J.T.; Seehausen, O.; Groothuis, T.G.G.; Maan, M.E. Developmental effects of visual environment on species-assortative mating preferences in Lake Victoria cichlid fish. J. Evol. Biol. 2017, 30, 289–299. [Google Scholar] [CrossRef]
- Aller, T.; Caldwell, R.L. Investigation of the possible presence of an aggregation pheromone in the milkweed bugs, Oncopeltus fasciatus and Lygaeus kalmii. Physiol. Entomol. 1979, 4, 287–290. [Google Scholar] [CrossRef]
- Hassanali, A.; Njagi, P.G.N.; Bashir, M.O. Chemical ecology of locusts and related acridids. Annu. Rev. Entomol. 2005, 50, 223–245. [Google Scholar] [CrossRef]
- Webster, B.; Bruce, T.; Pickett, J.; Hardie, J. Volatiles functioning as host cues in a blend become nonhost cues when presented alone to the black bean aphid. Anim. Behav. 2010, 79, 451–457. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef]
- Ejima, A. Pleiotropic actions of the male pheromone cis-vaccenyl acetate in Drosophila melanogaster. J. Comp. Physiol. A 2015, 201, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Aluja, M.; Prokopy, R.J. Host odor and visual stimulus interaction during intratree host finding behavior of Rhagoletis pomonella flies. J. Chem. Ecol. 1993, 19, 2671–2696. [Google Scholar] [CrossRef]
- Castrovillo, P.J.; Carde, R.T. Environmental-regulation of female calling and male pheromone response periodicities in the codling moth (Laspeyresia pomonella). J. Insect Physiol. 1979, 25, 659–667. [Google Scholar] [CrossRef]
- Steed, B.E.; Wagner, M.R. Seasonal pheromone response by Ips pini in northern Arizona and western Montana, USA. Agric. For. Entomol. 2008, 10, 189–203. [Google Scholar] [CrossRef]
- De Meester, L. Evolutionary potential and local genetic differentiation in a phenotypically plastic trait of a cyclical parhenogen, Daphnia magna. Evolution 1996, 3, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Gorter, J.A.; Jagadeesh, S.; Gahr, C.; Boonekamp, J.J.; Levine, J.D.; Billeter, J.C. The nutritional and hedonic value of food modulate sexual receptivity in Drosophila melanogaster females. Sci. Rep. 2016, 6, 19441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorter, J.A. The Nature and Nurture of Female Receptivity: A Study in Drosophila melanogaster; University of Groningen: Groningen, The Netherlands, 2018; Chapter 4. [Google Scholar]
- Mery, F.; Varela, S.A.; Danchin, E.; Blanchet, S.; Parejo, D.; Coolen, I.; Wagner, R.H. Public versus personal information for mate copying in an invertebrate. Curr. Biol. 2009, 19, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Calabria, G.; Maca, J.; Bachli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- National Plant Protection Organization (Ed.) First Findings of Drosophila suzukii; Netherlands Food and Consumer Product Safety Authority: Wageningen, The Netherlands, 2012.
- Eggink, F. Suzuki-fruitvlieg rukt op in Nederland. In NOS.nl; Nos.nl: Hilversum, The Netherlands, 2014; Available online: https://nos.nl/artikel/2001130-suzuki-fruitvlieg-rukt-op-in-nederland.html (accessed on 15 December 2019).
- Nijland, R.; Helsen, H. Zoeken naar de zwakke plekken van suzuki. Wagening. World 2015, 2015, 22–27. [Google Scholar]
- Hamby, K.A.; Bolda, M.P.; Sheehan, M.E.; Zalom, F.G. Seasonal monitoring for Drosophila suzukii (Diptera: Drosophilidae) in California commercial raspberries. Environ. Entomol. 2014, 43, 1008–1018. [Google Scholar] [CrossRef]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef]
- Bal, H.K.; Adams, C.; Grieshop, M. Evaluation of off-season potential breeding sources for spotted wing Drosophila (Drosophila suzukii Matsumura) in Michigan. J. Econ. Entomol. 2017, 110, 2466–2470. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Thistlewood, H.M.A.; Gill, P.; Beers, E.H.; Shearer, P.W.; Walsh, D.B.; Rozema, B.M.; Acheampong, S.; Castagnoli, S.; Yee, W.L.; Smytheman, P.; et al. Spatial analysis of seasonal dynamics and overwintering of Drosophila suzukii (Diptera: Drosophilidae) in the Okanagan-Columbia basin, 2010–2014. Environ. Entomol. 2018, 47, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreazza, F.; Bernardi, D.; Dos Santos, R.S.S.; Garcia, F.R.M.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila suzukii in southern neotropical region: Current status and future perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, L.A.; Mendes, M.F.; Kruger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R.M. Global potential distribution of Drosophila suzukii (Diptera, Drosophilidae). PLoS ONE 2017, 12, e0174318. [Google Scholar] [CrossRef] [Green Version]
- Cha, D.H.; Adams, T.; Rogg, H.; Landolt, P.J. Identification and field evaluation of fermentation volatiles from wine and vinegar that mediate attraction of spotted wing Drosophila, Drosophila suzukii. J. Chem. Ecol. 2012, 38, 1419–1431. [Google Scholar] [CrossRef]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing Drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef]
- Bolda, M.P.; Goodhue, R.E.; Zalom, F.G. Spotted wing Drosophila: Potential economic impact of a newly established pest. Agric. Econ. Res. Update 2010, 13, 5–8. [Google Scholar]
- Wang, X.G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Arno, J.; Sola, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Panel, A.D.C.; Zeeman, L.; van der Sluis, B.J.; van Elk, P.; Pannebakker, B.A.; Wertheim, B.; Helsen, H.H.M. Overwintered Drosophila suzukii are the main source for infestations of the first fruit crops of the season. Insects 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnsworth, D.; Hamby, K.A.; Bolda, M.; Goodhue, R.E.; Williams, J.C.; Zalom, F.G. Economic analysis of revenue losses and control costs associated with the spotted wing drosophila, Drosophila suzukii (Matsumura), in the California raspberry industry. Pest Manag. Sci. 2017, 73, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gress, B.E.; Zalom, F.G. Identification and risk assessment of spinosad resistance in a California population of Drosophila suzukii. Pest Manag. Sci. 2019, 75, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Schetelig, M.F.; Lee, K.Z.; Otto, S.; Talmann, L.; Stökl, J.; Degenkolb, T.; Vilcinskas, A.; Halitschke, R. Environmentally sustainable pest control options for Drosophila suzukii. J. Appl. Entomol. 2018, 142, 3–17. [Google Scholar] [CrossRef]
- Beers, E.H.; Van Steenwyk, R.A.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef]
- Lebreton, S.; Borrero-Echeverry, F.; Gonzalez, F.; Solum, M.; Wallin, E.A.; Hedenstrom, E.; Hansson, B.S.; Gustavsson, A.L.; Bengtsson, M.; Birgersson, G.; et al. A Drosophila female pheromone elicits species-specific long-range attraction via an olfactory channel with dual specificity for sex and food. BMC Biol. 2017, 15, 88. [Google Scholar] [CrossRef] [Green Version]
- Becher, P.G.; Flick, G.; Rozpedowska, E.; Schmidt, A.; Hagman, A.; Lebreton, S.; Larsson, M.C.; Hansson, B.S.; Piskur, J.; Witzgall, P.; et al. Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol. 2012, 26, 822–828. [Google Scholar] [CrossRef]
- Ebbs, M.L.; Amrein, H. Taste and pheromone perception in the fruit fly Drosophila melanogaster. Pflüg. Arch. Eur. J. Physiol. 2007, 454, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Diaz, C.; Martin, F.; Garcia-Fernandez, J.M.; Alcorta, E. The two main olfactory receptor families in Drosophila, ORs and IRs: A comparative approach. Front. Cell. Neurosci. 2018, 12, 253. [Google Scholar] [CrossRef]
- Datta, S.R.; Vasconcelos, M.L.; Ruta, V.; Luo, S.; Wong, A.; Demir, E.; Flores, J.; Balonze, K.; Dickson, B.J.; Axel, R. The Drosophila pheromone cVA activates a sexually dimorphic neural circuit. Nature 2008, 452, 473–477. [Google Scholar] [CrossRef]
- Heimbeck, G.; Bugnon, V.; Gendre, N.; Keller, A.; Stocker, R.F. A central neural circuit for experience-independent olfactory and courtship behavior in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 15336–15341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stensmyr, M.C.; Dweck, H.K.M.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebreton, S.; Becher, P.G.; Hansson, B.S.; Witzgall, P. Attraction of Drosophila melanogaster males to food-related and fly odours. J. Insect Physiol. 2012, 58, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, B.; Allemand, R.; Vet, L.E.M.; Dicke, M. Effects of aggregation pheromone on individual behaviour and food web interactions: A field study on Drosophila. Ecol. Entomol. 2006, 31, 216–226. [Google Scholar] [CrossRef]
- Butterworth, F.M. Lipids of Drosophila: A newly detected lipid in the male. Science 1969, 163, 1356–1357. [Google Scholar] [CrossRef]
- Jallon, J.-M. A few chemical words exchanged by Drosophila during courtship and mating. Behav. Genet. 1984, 14, 441–478. [Google Scholar] [CrossRef]
- Billeter, J.C.; Levine, J.D. Who is he and what is he to you? Recognition in Drosophila melanogaster. Curr. Opin. Neurobiol. 2013, 23, 17–23. [Google Scholar] [CrossRef]
- Wertheim, B.; Dicke, M.; Vet, L.E.M. Behavioural plasticity in support of a benefit for aggregation pheromone use in Drosophila melanogaster. Entomol. Exp. Appl. 2002, 103, 61–71. [Google Scholar] [CrossRef]
- Spieth, H.T. Courtship behavior in Drosophila. Annu. Rev. Entomol. 1974, 19, 385–405. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Wertheim, B. The mode of evolution of aggregation pheromones in Drosophila species. J. Evol. Biol. 2005, 18, 1253–1263. [Google Scholar] [CrossRef]
- Wertheim, B.; Marchais, J.; Vet, L.E.M.; Dicke, M. Allee effect in larval resource exploitation in Drosophila: An interaction among density of adults, larvae, and micro- organisms. Ecol. Entomol. 2002, 27, 608–617. [Google Scholar] [CrossRef]
- Rohlfs, M.; Obmann, B.; Petersen, R. Competition with filamentous fungi and its implication for a gregarious lifestyle in insects living on ephemeral resources. Ecol. Entomol. 2005, 30, 556–563. [Google Scholar] [CrossRef]
- Trienens, M.; Keller, N.P.; Rohlfs, M. Fruit, flies and filamentous fungi—Experimental analysis of animal-microbe competition using Drosophila melanogaster and Aspergillus mould as a model system. Oikos 2010, 119, 1765–1775. [Google Scholar] [CrossRef]
- Etienne, R.; Wertheim, B.; Hemerik, L.; Schneider, P.; Powell, J. The interaction between dispersal, the Allee effect andscramble competition affects population dynamics. Ecol. Model. 2002, 148, 153–168. [Google Scholar] [CrossRef]
- Everaerts, C.; Farine, J.P.; Cobb, M.; Ferveur, J.F. Drosophila cuticular hydrocarbons revisited: Mating status alters cuticular profiles. PLoS ONE 2010, 5, e9607. [Google Scholar] [CrossRef]
- Kent, C.; Azanchi, R.; Smith, B.; Formosa, A.; Levine, J.D. Social context influences chemical communication in D. melanogaster males. Curr. Biol. 2008, 18, 1384–1389. [Google Scholar] [CrossRef] [Green Version]
- Krupp, J.J.; Kent, C.; Billeter, J.C.; Azanchi, R.; So, A.K.C.; Schonfeld, J.A.; Smith, B.P.; Lucas, C.; Levine, J.D. Social experience modifies pheromone expression and mating behavior in male Drosophila melanogaster. Curr. Biol. 2008, 18, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Cobb, M.; Connolly, K.; Burnet, B. Courtship behavior in the melanogaster species subgroup of Drosophila. Behaviour 1985, 95, 203–231. [Google Scholar]
- Billeter, J.C.; Atallah, J.; Krupp, J.J.; Millar, J.G.; Levine, J.D. Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 2009, 461, 987. [Google Scholar] [CrossRef]
- Van Der Goes Van Naters, W. Drosophila Pheromones: From Reception to Perception. In Neurobiology of Chemical Communication; Mucignat-Caretta, C., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014. [Google Scholar]
- Karageorgi, M.; Bracker, L.B.; Lebreton, S.; Minervino, C.; Cavey, M.; Siju, K.P.; Grunwald Kadow, I.C.; Gompel, N.; Prud’homme, B. Evolution of multiple sensory systems drives novel egg-laying behavior in the fruit pest Drosophila suzukii. Curr. Biol. 2017, 27, 847–853. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, K.R.; Abraham, J.; Angeli, S.; Syed, Z.; Rodriguez-Saona, C. Advances in the chemical ecology of the spotted wing Drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 2018, 44, 922–939. [Google Scholar] [CrossRef] [PubMed]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, reproductive output and population growth of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae) on artificial diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prevost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the Asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, J.R.; Unelius, C.R.; Lee, J.C.; Suckling, D.M.; Qian, M.C.; Bruck, D.J. Attractiveness of fermentation and related products to spotted wing Drosophila (Diptera: Drosophilidae). Environ. Entomol. 2014, 43, 439–447. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Hesler, S.P.; Cha, D.H.; Loeb, G.M. Behavioral response of spotted-wing drosophila, Drosophila suzukii Matsumura, to aversive odors and a potential oviposition deterrent in the field. Pest Manag. Sci. 2016, 72, 701–706. [Google Scholar] [CrossRef]
- Krause Pham, C.; Ray, A. Conservation of olfactory avoidance in Drosophila species and identification of repellents for Drosophila suzukii. Sci. Rep. 2015, 5, 11527. [Google Scholar] [CrossRef]
- Revadi, S.; Lebreton, S.; Witzgall, P.; Anfora, G.; Dekker, T.; Becher, P.G. Sexual Behavior of Drosophila suzukii. Insects 2015, 6, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Anagnostou, C.; Dorsch, M.; Rohlfs, M. Influence of dietary yeasts on Drosophila melanogaster life-history traits. Entomol. Exp. Appl. 2010, 136, 1–11. [Google Scholar] [CrossRef]
- Carvalho, M.; Schwudke, D.; Sampaio, J.L.; Palm, W.; Riezman, I.; Dey, G.; Gupta, G.D.; Mayor, S.; Riezman, H.; Shevchenko, A.; et al. Survival strategies of a sterol auxotroph. Development 2010, 137, 3675–3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond-Barbosa, D.; Spradling, A.C. Stem cells and their progeny respond to nutritional changes during Drosophila oogenesis. Dev. Biol. 2001, 231, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, J.A.; Eisen, J.A.; Kopp, A. Yeast communities of diverse Drosophila species: Comparison of two symbiont groups in the same hosts. Appl. Environ. Microbiol. 2012, 78, 7327–7336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurung, K.; Wertheim, B.; Falcao Salles, J. The microbiome of pest insects: It is not just bacteria. Entomol. Exp. Appl. 2019, 167, 156–170. [Google Scholar] [CrossRef] [Green Version]
- Cha, D.H.; Adams, T.; Werle, C.T.; Sampson, B.J.; Adamczyk, J.J., Jr.; Rogg, H.; Landolt, P.J. A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest Manag. Sci. 2014, 70, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Cha, D.H.; Hesler, S.P.; Park, S.; Adams, T.B.; Zack, R.S.; Rogg, H.; Loeb, G.M.; Landolt, P.J. Simpler is better: Fewer non-target insects trapped with a four-component chemical lure vs. a chemically more complex food-type bait for Drosophila suzukii. Entomol. Exp. Appl. 2015, 154, 251–260. [Google Scholar] [CrossRef]
- Cha, D.H.; Landolt, P.J.; Adams, T.B. Effect of chemical ratios of a microbial-based feeding attractant on trap catch of Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2017, 46, 907–915. [Google Scholar] [CrossRef]
- Dekker, T.; Revadi, S.; Mansourian, S.; Ramasamy, S.; Lebreton, S.; Becher, P.G.; Angeli, S.; Rota-Stabelli, O.; Anfora, G. Loss of Drosophila pheromone reverses its role in sexual communication in Drosophila suzukii. Proc. R. Soc. Biol. 2015, 282, 20143018. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, H.; Takahashi, K.H.; Kimura, M.T. Spatial distributions and clutch sizes of Drosophila species ovipositing on cherry fruits of different stages. Popul. Ecol. 2006, 48, 233–237. [Google Scholar] [CrossRef]
- Matavelli, C.; Carvalho, M.J.A.; Martins, N.E.; Mirth, C.K. Differences in larval nutritional requirements and female oviposition preference reflect the order of fruit colonization of Zaprionus indianus and Drosophila simulans. J. Insect Physiol. 2015, 82, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Silva-Soares, N.F.; Nogueira-Alves, A.; Beldade, P.; Mirth, C.K. Adaptation to new nutritional environments: Larval performance, foraging decisions, and adult oviposition choices in Drosophila suzukii. BMC Ecol. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.T.; Koivunen, E.E.; Swett, C.L.; Hamby, K.A. Associations between Drosophila suzukii (Diptera: Drosophilidae) and fungi in raspberries. Environ. Entomol. 2019, 48, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Belien, T.; Bylemans, D. Olfactory preference of Drosophila suzukii shifts between fruit and fermentation cues over the season: Effects of physiological status. Insects 2019, 10, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.S.; Wallingford, A.K.; Loeb, G.M.; Lee, J.C. Physiological status of Drosophila suzukii (Diptera: Drosophilidae) affects their response to attractive odours. J. Appl. Entomol. 2018, 142, 473–482. [Google Scholar] [CrossRef]
- Swoboda-Bhattarai, K.A.; McPhie, D.R.; Burrack, H.J. Reproductive status of Drosophila suzukii (Diptera: Drosophilidae) females influences ttraction to fermentation-based baits and ripe fruits. J. Econ. Entomol. 2017, 110, 1648–1652. [Google Scholar] [CrossRef]
- Hamby, K.A.; Bellamy, D.E.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; York, R.M.; Biondi, A. Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Grassi, A.; Gottardello, A.; Dalton, D.T.; Tait, G.; Rendon, D.; Ioriatti, C.; Gibeaut, D.; Stacconi, M.V.R.; Walton, V.M. Seasonal reproductive biology of Drosophila suzukii (Diptera: Drosophilidae) in temperate climates. Environ. Entomol. 2018, 47, 166–174. [Google Scholar] [CrossRef]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, D.M.; Leach, H.L.; Xu, P.; Dong, K.; Isaacs, R.; Gut, L.J. Comparative antennal and behavioral responses of summer and winter morph Drosophila suzukii (Diptera: Drosophilidae) to ecologically relevant volatiles. Environ. Entomol. 2018, 47, 700–706. [Google Scholar] [CrossRef]
- Menger, D.J.; Omusula, P.; Holdinga, M.; Homan, T.; Carreira, A.S.; Vandendaele, P.; Derycke, J.L.; Mweresa, C.K.; Mukabana, W.R.; van Loon, J.J.A.; et al. Field evaluation of a push-pull system to reduce malaria transmission. PLoS ONE 2015, 10, e0123415. [Google Scholar] [CrossRef] [PubMed]
- Karuppuchamy, P.; Venugopal, S. Integrated Pest Management. In Ecofriendly Pest Management for Food Security; Academic Press Elsevier: Amsterdam, The Netherlands, 2016; pp. 651–684. [Google Scholar] [CrossRef]
- Baker, T.C. Chemical Control of Behavior. In Behaviour, 1st ed.; Brooks, G.T., Ed.; Pergamon Press Ltd.: Oxford, UK, 1985; Volume 9, pp. 621–672. [Google Scholar]
- Kruitwagen, A.; Beukeboom, L.W.; Wertheim, B. Optimization of native biocontrol agents, with parasitoids of the invasive pest Drosophila suzukii as an example. Evol. Appl. 2018, 11, 1473–1497. [Google Scholar] [CrossRef] [PubMed]
- Daane, K.M.; Wang, X.G.; Biondi, A.; Miller, B.; Miller, J.C.; Riedl, H.; Shearer, P.W.; Guerrieri, E.; Giorgini, M.; Buffington, M.; et al. First exploration of parasitoids of Drosophila suzukii in South Korea as potential classical biological agents. J. Pest Sci. 2016, 89, 823–835. [Google Scholar] [CrossRef]
- Girod, P.; Lierhmann, O.; Urvois, T.; Turlings, T.C.J.; Kenis, M.; Haye, T. Host specificity of Asian parasitoids for potential classical biological control of Drosophila suzukii. J. Pest Sci. 2018, 91, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girod, P.; Rossignaud, L.; Haye, T.; Turlings, T.C.J.; Kenis, M. Development of Asian parasitoids in larvae of Drosophila suzukii feeding on blueberry and artificial diet. J. Appl. Entomol. 2018, 142, 483–494. [Google Scholar] [CrossRef] [Green Version]
- EPPO. PM 6/2 (3) Import and release of non-indigenous biological control agents. EPPO Bull. 2014, 44, 320–329. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bale, J.; Bigler, E.; Hokkanen, H.M.T.; Loomans, A.M. Assessing risks of releasing exotic biological control agents of arthropod pests. Annu. Rev. Entomol. 2006, 51, 609–634. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.; Hinz, H.; Mulema, J.; Weyl, P.; Ryan, M.J. Biological control and the Nagoya Protocol on access and benefit sharing—A case of effective due diligence. Biocontrol Sci. Technol. 2018, 28, 914–926. [Google Scholar] [CrossRef]
- Lommen, S.T.E.; de Jong, P.W.; Pannebakker, B.A. It is time to bridge the gap between exploring and exploiting: Prospects for utilizing intraspecific genetic variation to optimize arthropods for augmentative pest control—A review. Entomol. Exp. Appl. 2017, 162, 108–123. [Google Scholar] [CrossRef]
- Lee, K.Z.; Vilcinskas, A. Analysis of virus susceptibility in the invasive insect pest Drosophila suzukii. J. Invertebr. Pathol. 2017, 148, 138–141. [Google Scholar] [CrossRef]
- Hübner, A.; Englert, C.; Herz, A. Effect of entomopathogenic nematodes on different developmental stages of Drosophila suzukii in and outside fruits. BioControl 2017, 62, 669–680. [Google Scholar] [CrossRef]
- Bybee-Finley, K.A.; Ryan, M.R. Advancing intercropping research and practices in industrialized agricultural landscapes. Agriculture 2018, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Area-Wide Integrated Pest Management and the Sterile Insect Technique. In Sterile Insect Technique; Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Springer: Dordrecht, The Netherlands, 2005; pp. 39–68. [Google Scholar] [CrossRef]
- Krafsur, E.S. Sterile insect technique for suppressing and eradicating insect populations: 55 years and counting. J. Agric. Entomol. 1998, 15, 303–317. [Google Scholar]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Caceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepenbrock, L.M.; Burrack, H.J. Efficacy of bait sprays for the control of Drosophila suzukii (Diptera: Drosophilidae) in Blueberries, 2014. Arthropod Manag. Tests 2015, 40. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Baronio, C.A.; Pasinato, J.; Nava, D.E.; Botton, M. Toxicities and effects of insecticidal toxic baits to control Drosophila suzukii and Zaprionus indianus (Diptera: Drosophilidae). Pest Manag. Sci. 2017, 73, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swoboda-Bhattarai, K.A.; Burrack, H.J. Influence of edible fruit coatings onDrosophila suzukii(Matsumura) (Diptera: Drosophilidae) oviposition and development. Int. J. Pest Manag. 2014, 60, 279–286. [Google Scholar] [CrossRef]
- Kuesel, R.; Hicks, D.S.; Archer, K.; Sciligo, A.; Bessin, R.; Gonthier, D. Effects of fine-mesh exclusion netting on pests of blackberry. Insects 2019, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Leadbeater, E.; Chittka, L. Social learning in insects—From miniature brains to consensus building. Curr. Biol. 2007, 17, R703–R713. [Google Scholar] [CrossRef] [Green Version]
- Danchin, E.; Giraldeau, L.-A.; Valone, T.J.; Wagner, R.H. Public information: From nosy neighbors to cultural evolution. Science 2004, 305, 487–491. [Google Scholar] [CrossRef] [Green Version]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkema, J.T.; Dicke, M.; Wertheim, B. Context-Dependence and the Development of Push-Pull Approaches for Integrated Management of Drosophila suzukii. Insects 2019, 10, 454. https://doi.org/10.3390/insects10120454
Alkema JT, Dicke M, Wertheim B. Context-Dependence and the Development of Push-Pull Approaches for Integrated Management of Drosophila suzukii. Insects. 2019; 10(12):454. https://doi.org/10.3390/insects10120454
Chicago/Turabian StyleAlkema, Jeroen T., Marcel Dicke, and Bregje Wertheim. 2019. "Context-Dependence and the Development of Push-Pull Approaches for Integrated Management of Drosophila suzukii" Insects 10, no. 12: 454. https://doi.org/10.3390/insects10120454