Ants Sense, and Follow, Trail Pheromones of Ant Community Members

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.1.1. Lasius Niger
2.1.2. Camponotus Modoc
2.1.3. Myrmica Rubra
2.2. Gas Chromatographic-Electroantennographic Detection (GC-EAD) Analyses of Synthetic Ant Trail Pheromones
2.3. General Design of Trail-Following Bioassays
2.4. Statistics
3. Results
3.1. Gas Chromatographic-Electroantennographic Detection (GC-EAD) Analyses of Synthetic Ant Trail Pheromones
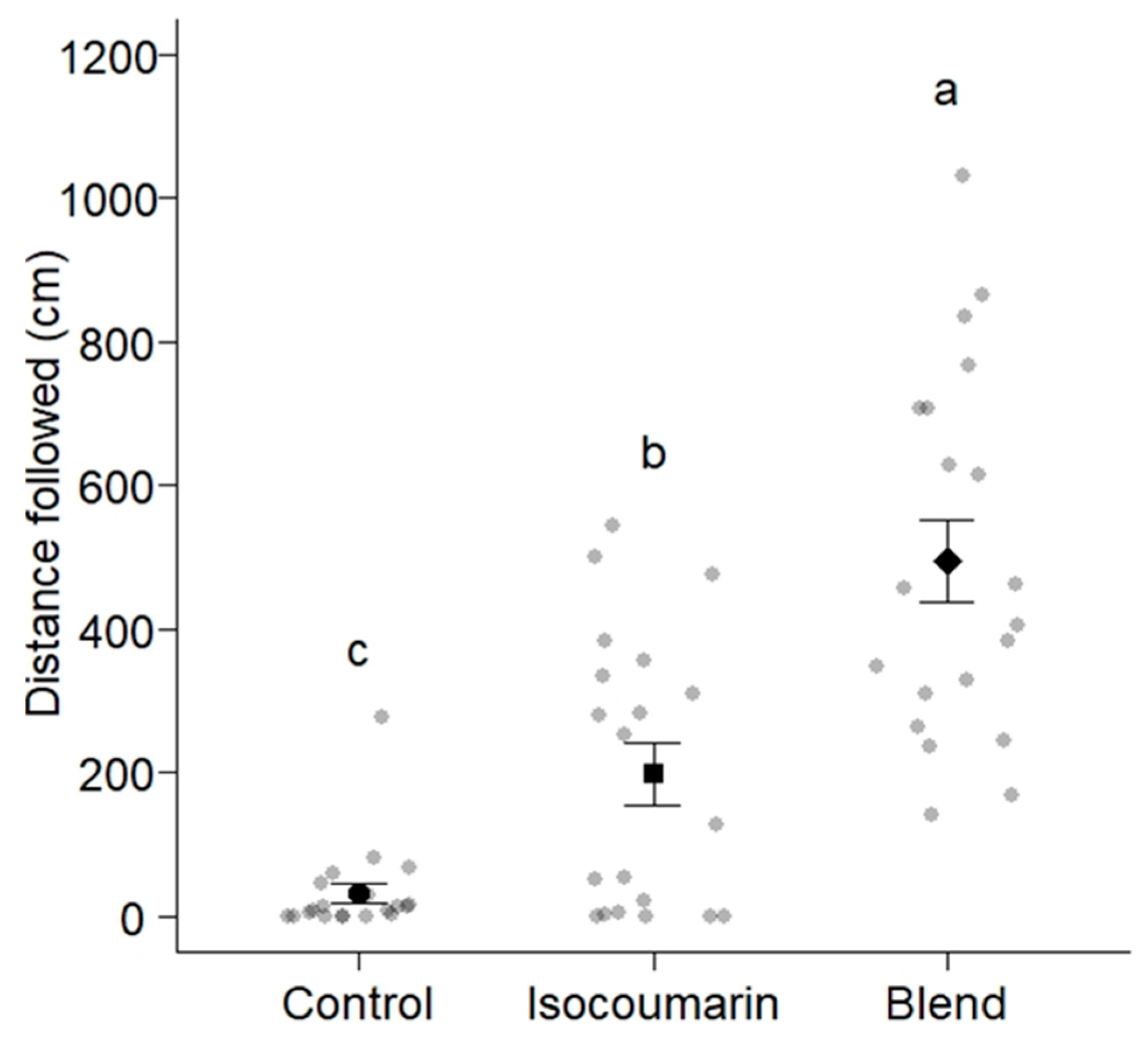
3.2. Trail-Following Bioassays
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Ants, 1st ed.; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Morgan, E.D. Trail pheromone of ants. Physiol. Entomol. 2009, 34, 1–17. [Google Scholar] [CrossRef]
- Detrain, C.; Deneubourg, J.L. Collective decision-making and foraging patterns in ants and honeybees. Adv. Insect Physiol. 2008, 35, 123–173. [Google Scholar] [CrossRef]
- Beckers, R.; Deneubourg, J.L.; Goss, S.; Pasteels, J.M. Collective decision making through food recruitment. Insectes Soc. 1990, 37, 258–267. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Alonso, L.E. Pheromone directed behaviour in ants. In Pheromone Communication in Social Insects; Ants, Wasps, Bees, and Termites, 1st ed.; Vander Meer, R.K., Breed, M.D., Espelie, K.E., Winston, M.L., Eds.; Westview Press: Boulder, CO, USA, 1998. [Google Scholar]
- Wilson, E.O. Trail sharing in ants. Psyche 1965, 72, 2–7. [Google Scholar] [CrossRef]
- Adams, E.S. Interaction between the ants Zacryptocerus maculatus and Azteca trigona: Interspecific parasitization of information. Biotropica 1990, 22, 200–206. [Google Scholar] [CrossRef]
- Gobin, B.; Peeters, C.; Billen, J.; Morgan, E.D. Interspecific trail following and commensalisms between the ponerine ant Gnamptogenys menadensis and the formicine ant Polyrhachis rufipes. J. Insect. Behav. 1998, 11, 361–369. [Google Scholar] [CrossRef]
- Menzel, F.; Pokorny, T.; Blüthgen, N.; Schmitt, T. Trail-sharing among tropical ants: Interspecific use of trail pheromones? Ecol. Entomol. 2010, 35, 495–503. [Google Scholar] [CrossRef]
- Klotz, J.H. Diel differences in foraging in two ant species (Hymenoptera: Formicidae). J. Kansas Entomol. Soc. 1984, 57, 111–118. [Google Scholar]
- Buckell, E.R. A list of the ants of British Columbia. J. Entomol. Soc. B. C. 1932, 29, 22–25. [Google Scholar]
- Higgins, R.J.; Lindgren, B.S. An evaluation of methods for sampling ants (Hymenoptera: Formicidae) in British Columbia, Canada. Can. Entomol. 2012, 144, 491–507. [Google Scholar] [CrossRef]
- Naumann, K.; Preston, W.B.; Ayre, G.L. An annotated checklist of the ants (Hymenoptera: Formicidae) of British Columbia. J. Entomol. Soc. B. C. 1999, 96, 29–68. [Google Scholar]
- Groden, E.; Drummond, F.A.; Garnas, J.; Franceour, A. Distribution of an invasive ant, Myrmica rubra (Hymenoptera: Formicidae), in Maine. J. Econ. Entomol. 2005, 98, 1774–1784. [Google Scholar] [CrossRef] [PubMed]
- Naumann, K.; Higgins, R.J. The European fire ant (Hymenoptera: Formicidae) as an invasive species: Impact on local ant species and other epigaeic arthropods. Can. Entomol. 2015, 147, 592–601. [Google Scholar] [CrossRef]
- Renyard, A.; Alamsetti, S.; Gries, R.; Munoz, A.; Gries, G. Identification of the trail pheromone of the carpenter ant, Camponotus modoc. J. Chem. Ecol. 2019. accepted for publication. [Google Scholar]
- Bestmann, H.J.; Kern, F.; Schäfer, D.; Witschel, M.C. 3,4-Dihydroisocoumarins, a new class of ant trail pheromones. Angew. Chem. Int. Ed. 1992, 31, 795–796. [Google Scholar] [CrossRef]
- Evershed, R.P.; Morgan, E.D.; Cammaerts, M.C. 3-Ethyl-2,5-dimethylpyrazine, the trail pheromone from the venom gland of eight species of Myrmica ants. Insect Biochem. 1982, 12, 383–391. [Google Scholar] [CrossRef]
- Attygalle, A.B.; Morgan, E.D. Identification of trail pheromone of the ant Tetramorium caespitum L. (Hymenoptera: Myrmicinae). Naturwissenschaften 1983, 70, 364–365. [Google Scholar] [CrossRef]
- Hölldobler, B.; Oldham, N.J.; Morgan, E.D.; König, W.A. Recruitment pheromones in the ants Aphaenogaster albisetosus and A. cockerelli (Hymenoptera: Formicidae). J. Insect Physiol. 1995, 41, 739–744. [Google Scholar] [CrossRef]
- Van Vorhis Key, S.E.; Baker, T.C. Trail-following responses of the Argentine ant, Iridomyrmex humilis (Mayr), to a synthetic trail pheromone component and analogs. J. Chem. Ecol. 1982, 8, 3–14. [Google Scholar] [CrossRef]
- Mackay, W.P.; Mackay, E. The Ants of New Mexico (Hymenoptera: Formicidae); Edwin Mellend Press: Lewiston, NY, USA, 2002. [Google Scholar]
- Wilson, E.O. A monographic revision of the ant genus Lasius. Bull. Mus. Comp. Zool. 1955, 113, 1–201. [Google Scholar]
- Wing, M.W. Taxonomic Revision of the Nearctic Genus Acanthomyops (Hymenoptera: Formicidae); Cornell University, New York State College of Agriculture, Agricultural Experimentation Station: Ithica, NY, USA, 1968. [Google Scholar]
- Arn, H.; Städler, E.; Rauscher, S. The electroantennographic detector—A selective and sensitive tool in the gas chromatographic analysis of insect pheromones. Z. Naturforsch. 1975, 30c, 11–12. [Google Scholar] [CrossRef]
- Gries, R.; Khaskin, G.; Gries, G.; Bennett, R.G.; King, G.G.S.; Morewood, P.; Slessor, K.N.; Morewood, W.D. (Z,Z)-4,7-Tridecadien-(S)-2-yl acetate: Sex pheromone of Douglas-fir cone gall midge, Contarinia oregonensis. J. Chem. Ecol. 2002, 28, 2283–2297. [Google Scholar] [CrossRef] [PubMed]
- Staddon, B.W.; Everton, I.J. Haemolymph of the milkweed bug Oncopeltus fasciatus (Heteroptera: Lygaeidae): Inorganic constituents and amino acids. Comp. Biochem. Physiol. A 1980, 65, 371–374. [Google Scholar] [CrossRef]
- Hothorn, T.; Betz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Lemon, J. Plotrix: A package in the red light district of R. R-News 2006, 6, 8–12. [Google Scholar]
- Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef]
- Hoefele, D.; Chalissery, J.M.; Gries, R.; Gries, G. Effects of trail pheromone purity, dose and type of placement on recruiting European fire ants, Myrmica rubra, to food baits. J. Entomol. Soc. B. C. 2019. under review. [Google Scholar]
- Higgins, R.J.; (Thompson Rivers University, Kamloops, BC, Canada); Hoefele, D.; (Simon Fraser University, Burnaby, BC, Canada). Personal communication, 2017.
- Fedoseeva, E.B. A technological approach to the description of group foraging in the ant Myrmica rubra. Entomol. Rev. 2015, 95, 984–999. [Google Scholar] [CrossRef]
- Carroll, C.R.; Janzen, D.H. Ecology of foraging by ants. Annu. Rev. Ecol. Syst. 1973, 4, 231–257. [Google Scholar] [CrossRef]
- Welzel, K.F.; Choe, D.H. Development of a pheromone-assisted baiting technique for Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2016, 109, 1303–1309. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Species | Name of Pheromone (Amount Tested; Ant Equivalents (AEs)) | Pheromone Structures (Synthetic Sources a–e) |
|---|---|---|
| Ca. modoc | (2S,4R,5S)-2,4-Dimethyl-5-hexanolide (7.5 ng; 2 AEs) (“hexanolide”) [16] |  (a) (a) |
| La. niger | 3,4-Dihydro-8-hydroxy-3-7-trimethylisocoumarin (0.5 ng; 1 AE) (“isocoumarin”) [17] |  (a) (a) |
| M. rubra | 3-Ethyl-2,5-dimethylpyrazine (5 ng; 1 AE) [18] |  (b) (b) |
| T. caespitum | 2,5-Dimethylpyrazine (1 ng; 1 AE) [19] |  (c) (c) |
| N. cockerelli | 4-Methyl-3-heptanone (10 ng;1 AE) [20] |  (d) (d) |
| Li. humilis | (Z)-9-Hexadecenal (10 ng; 1 AE) [21] | CH3-(CH2)4-CH=CH-(CH2)7-CHO (e) |
| Distribution | Species | Trail Pheromone | Produced by/Antennal Response | ||
|---|---|---|---|---|---|
| La. niger | Ca. modoc | M. rubra | |||
| Sympatric | La. niger | Isocoumarin * | yes/yes | yes/yes | no/yes |
| Ca. modoc | Hexanolide ** | no/yes | yes/yes | no/yes | |
| M. rubra | 3-Ethyl-2,5-dimethylpyrazine | no/yes | no/yes | yes/yes | |
| Allopatric | T. caespitum | 2,5-Dimethylpyrazine | no/yes | no/yes | no/yes |
| N. albisetosus | 4-Methyl-3-heptanone | no/yes | no/no | no/no | |
| Li. humilis | (Z)-9-Hexadecenal | no/yes | no/no | no/no | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chalissery, J.M.; Renyard, A.; Gries, R.; Hoefele, D.; Alamsetti, S.K.; Gries, G. Ants Sense, and Follow, Trail Pheromones of Ant Community Members. Insects 2019, 10, 383. https://doi.org/10.3390/insects10110383
Chalissery JM, Renyard A, Gries R, Hoefele D, Alamsetti SK, Gries G. Ants Sense, and Follow, Trail Pheromones of Ant Community Members. Insects. 2019; 10(11):383. https://doi.org/10.3390/insects10110383
Chicago/Turabian StyleChalissery, Jaime M., Asim Renyard, Regine Gries, Danielle Hoefele, Santosh Kumar Alamsetti, and Gerhard Gries. 2019. "Ants Sense, and Follow, Trail Pheromones of Ant Community Members" Insects 10, no. 11: 383. https://doi.org/10.3390/insects10110383