Gamasina Mites (Acari: Mesostigmata) Associated with Animal Remains in the Mediterranean Region of Navarra (Northern Spain)


Abstract
:1. Introduction
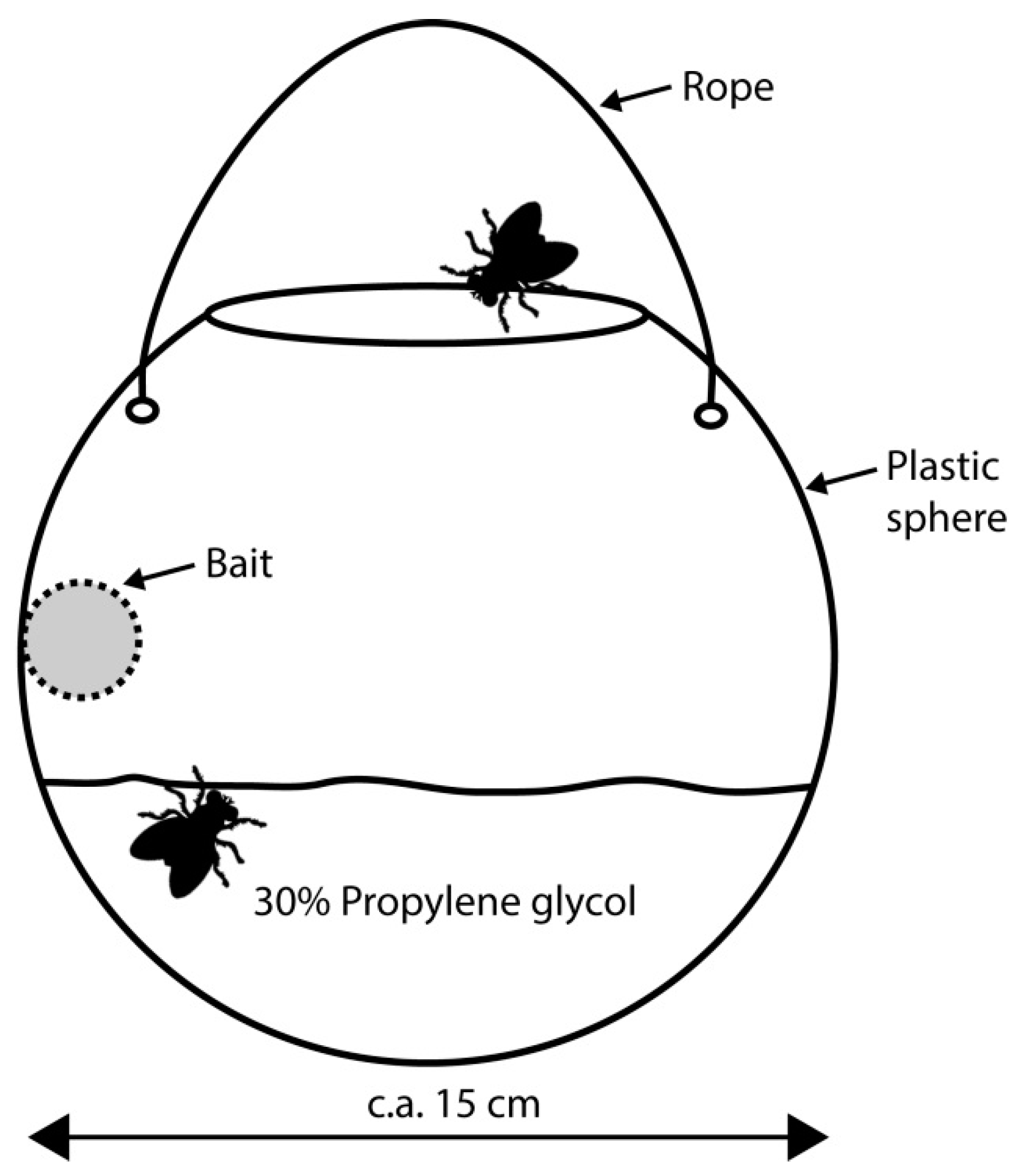
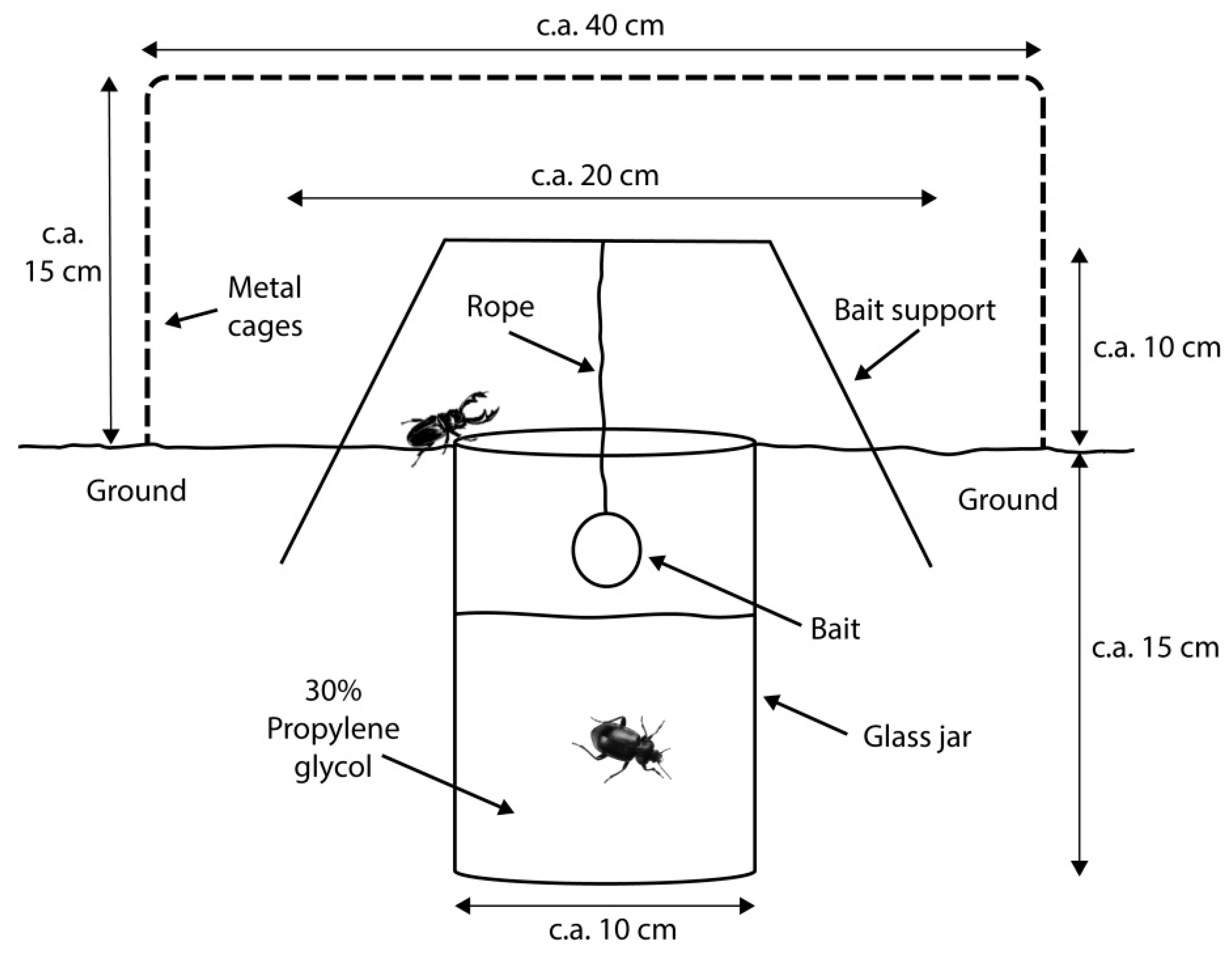
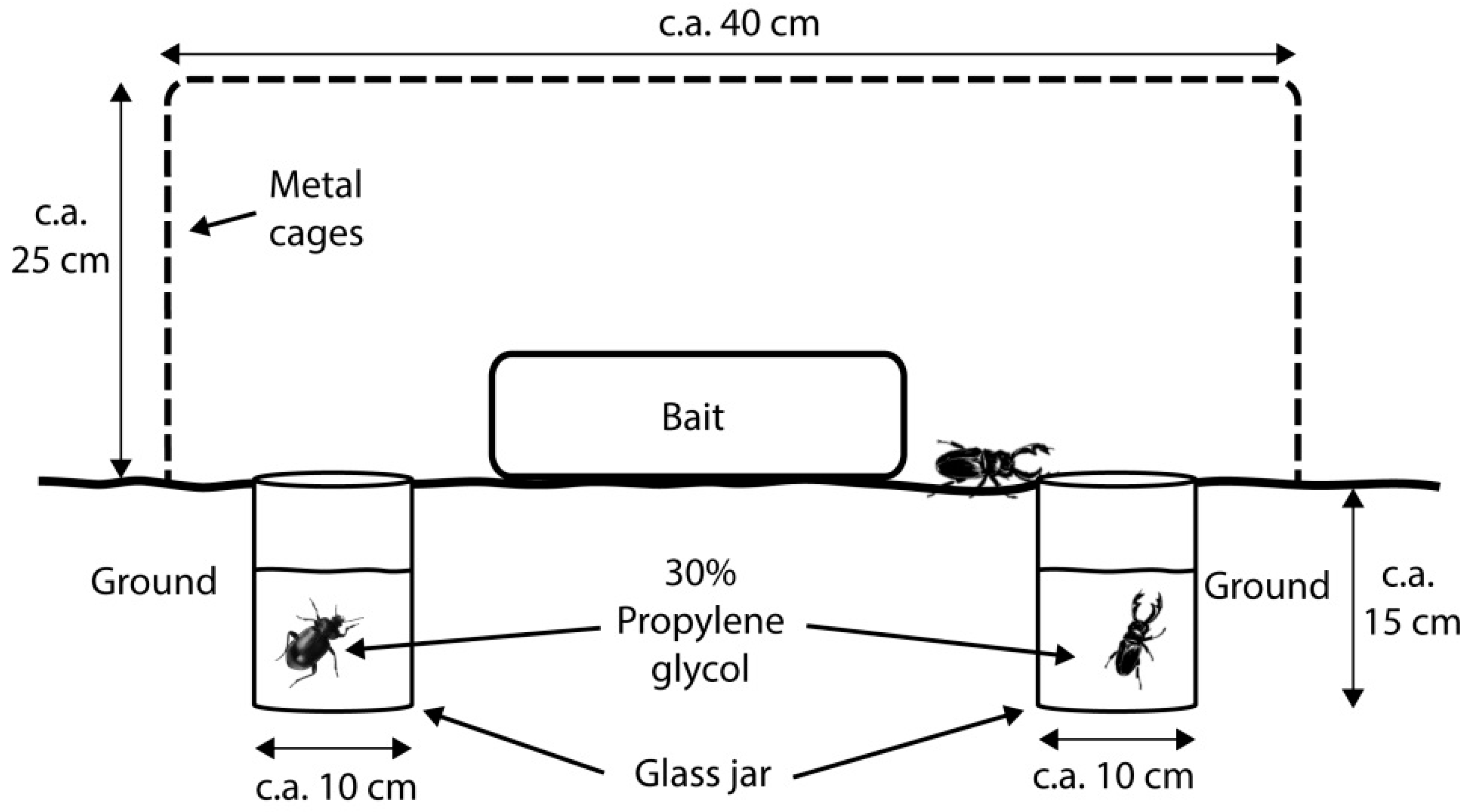
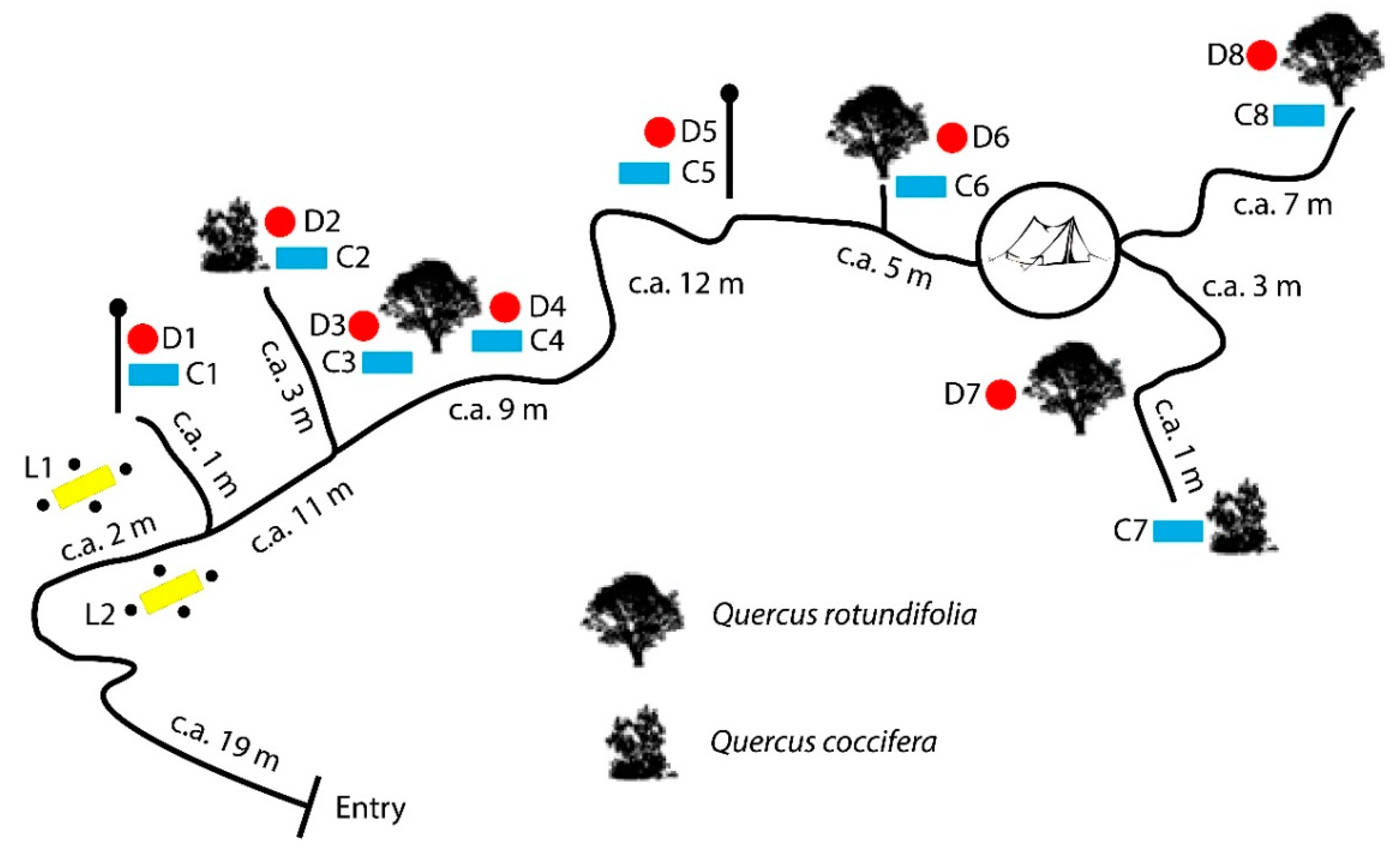
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bregetova, N.G.; Vainshtein, B.A.; Kadite, B.A.; Koroleva, E.V.; Petrovs, A.D.; Tikhomirov, S.I.; Shcherbat, G.I. A Key to the Soil-Inhabiting Mites. Mesostigmata; Gilyarov, M.S., Ed.; Nauka: Leningrad, Rusia, 1977. [Google Scholar]
- Hyatt, K.H.; Emberson, R.M. A review of the Macrochelidae (Acari: Mesostigmata) of the British Isles. Bull. Br. Mu. Nat. Hist. Zool. 1988, 54, 63–125. [Google Scholar]
- Saloña-Bordas, M.I.; Perotti, M.A. Acarología forense. Cienc. Forense 2015, 12, 91–112. [Google Scholar]
- Goff, M.L. Use of acari in establishing a postmortem interval in a homicide case on the island of Ohau, Hawaii. In Modern Acarology: Proceedings of the VIII International Congress of Acarology; Dusbábek, F., Bukva, V., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1991; pp. 439–442. ISBN1 9051030541. ISBN2 9789051030549. [Google Scholar]
- Mašán, P. Mites (Acarina) associated with burying and carrion beetles (Coleoptera, Silphidae) and description of Poecilochirus mrciaki sp.n. (Mesostigmata, Gamasina). Biol. Bratisl. 1999, 54, 515–524. [Google Scholar]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; ISBN 9780896726208. [Google Scholar]
- Perotti, M.A.; Braig, H.R. Phoretic mites associated with animal and human decomposition. Exp. Appl. Acarol. 2009, 49, 85–124. [Google Scholar] [CrossRef] [PubMed]
- Camerik, A.M. Phoresy revisited. In Trends in Acarology; Sabelis, M., Bruin, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 333–336. ISBN 978-90-481-9837-5. [Google Scholar]
- Perotti, M.A.; Braig, H.R.; Goff, M.L. Phoretic Mites and Carcasses: Acari Transported by Organisms Associated with Animal and Human Decomposition. In Current Concepts in Forensic Entomology; Amendt, J., Campobasso, C.P., Grassberger, M., Goff, M.L., Eds.; Springer: New York, NY, USA, 2010; pp. 69–92. ISBN 9781402096846. [Google Scholar]
- Saloña-Bordas, M.I.; Perotti, M.A. First contribution of mites (Acari) to the forensic analysis of hanged corpses: A case study from Spain. Forensic Sci. Int. 2014, 244, e6–e11. [Google Scholar] [CrossRef]
- Saloña-Bordas, M.I.; Bahillo de la Puebla, P.; Díaz Martín, B.; Sumner, J.; Perotti, M.A. Ixodes ricinus (Ixodidae), an occasional phoront on necrophagous and coprophagous beetles in Europe. Exp. Appl. Acarol. 2015, 65, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Farish, D.J.; Axtell, R.C. Phoresy redefined and examined in Macrocheles muscaedomesticae (Acarina: Marcochelidae). Acarologia 1971, 13, 16–29. [Google Scholar]
- Szymkowiak, P.; Górski, G.; Bajerlein, D. Passive dispersal in arachnids. Biol. Lett. 2007, 44, 75–101. [Google Scholar]
- Nehring, V.; Müller, J.K. Social environment affects the life history tactic of a phoretic mite. J. Evol. Biol. 2009, 22, 1616–1623. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, E.E.; Walter, D.E. Antennoseius (Vitzthumia) janus n.sp. (Acari: Ascidae), a mesostigmatic mite exhibiting adult female dimorphism. Can. J. Zool. 1989, 67, 1291–1310. [Google Scholar] [CrossRef]
- Athias-Binche, F.; Schwarz, H.H.; Meierhofer, I. Phoretic association of Neoseius novus (Ouds., 1902) (Acari: Uropodina) with Nicrophorus spp. (Coleoptera: Silphidae): A case of sympatric speciation? Int. J. Acarol. 1993, 19, 75–86. [Google Scholar] [CrossRef]
- Beaulieu, F.; Déchêne, A.D.; Walter, D.E. Phase morphs and phoresy: New species of Antennoseius (Vitzthumia) mites (Acari: Mesostigmata: Ascidae) associated with pyrophilous carabids (Carabidae: Sericoda spp.) in Alberta, Canada. Zootaxa 2008, 1961, 37–57. [Google Scholar]
- Lindquist, E.E.; Moraza, M.L. A new genus of flower-dwelling melicharid mites (Acari: Mesostigmata: Ascoidea) phoretic on bats and insects in Costa Rica and Brazil. Zootaxa 2008, 1685, 1–37. [Google Scholar]
- Prichard, J.G.; Kossoris, P.D.; Leibovitch, R.A.; Robertson, L.D.; Lovell, F.W. Implications of Trombiculid Mite Bites: Report of a Case and Submission of Evidence in a Murder Trial. J. Forensic Sci. 1986, 31, 301–306. [Google Scholar] [CrossRef]
- Solarz, K. Indoor mites and forensic acarology. Exp. Appl. Acarol. 2009, 49, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Turner, B. Forensic entomology: A template for forensic acarology? Exp. Appl. Acarol. 2009, 49, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Desch, C.E. Human hair follicle mites and forensic acarology. Exp. Appl. Acarol. 2009, 49, 143–146. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Ramos-Pastrana, Y.; Wolff, M. Postmortem interval estimation based on Chrysomya albiceps (Diptera, Calliphoridae) in a forensic case in the Andean Amazon, Caquetá, Colombia. Acta Amaz. 2017, 47, 369–374. [Google Scholar] [CrossRef]
- Saloña, M.I.; Moraza, M.L.; Carles-Tolrá, M.; Iraola, V.; Bahillo, P.; Yélamos, T.; Outerelo, R.; Alcaraz, R. Searching the soil: Forensic importance of edaphic fauna after the removal of a corpse. J. Forensic Sci. 2010, 55, 1652–1655. [Google Scholar] [CrossRef]
- Sumodan, P.K. Insect detectives. Resonance 2002, 7, 51–58. [Google Scholar] [CrossRef]
- Catts, E.P.; Goff, M.L. Forensic Entomology in Criminal Investigations. Annu. Rev. Entomol. 1992, 37, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Athias-Henriot, C. Mésostigmates (Uro.Excl.) édaphiques méditerráneens (Acaromorpha, Anactinotrichida). Acarologia 1961, 3, 381–509. [Google Scholar]
- Krantz, G.W. A manual of Acarology; Texas Tech University Press: Corvallis, OR, USA, 1978; ISBN 0882460641. [Google Scholar]
- Evans, G.O. Adults of the genus Crassicheles Karg (Acari: Mesostigmata). Int. J. Acarol. 1980, 6, 33–38. [Google Scholar] [CrossRef]
- Hyatt, K.H. Mites of the subfamily Parasitinae (Mesostigmata: Parasitidae) in the British Isles. Bull. Br. Mus. (Nat. Hist. Zool.) 1980, 38, 237–378. [Google Scholar]
- Karg, W. The Family Acari (Acarina), Milben, Unterordnung Parasitiformes (Anactinochaeta), Uropodina Kramer, Schildkroȯtenmilben; VEB Gustav Fischer Verlag: Jena, Germany, 1989; ISBN 3334003086. [Google Scholar]
- Karg, W. Acari (Acarina), Milben Parasitiformes (Anactinochaeta), Cohors Gamasina Leach; Gustav Fischer Verlag Jena: Raubmilben, Germany, 1993; ISBN 3334604454. [Google Scholar]
- Mašán, P.; Halliday, B. Review of the European genera of Eviphididae (Acari: Mesostigmata) and the species occurring in Slovakia. Zootaxa 2010, 2585, 1–122. [Google Scholar]
- Hallan, J. Synopsis of the described Arachnida of the world. Available online: http://bug.tamu.edu/research/collection/hallan/Acari/Family/Mesostigmata1.htm (accessed on 15 October 2018).
- Tüzün, A.; Murat, H.G.; Bilgili, G. Forensic Acarology. Türk Bilim. Derlemeler Derg. 2015, 8, 54–56. [Google Scholar]
- Dhooria, M.S. (Ed.) Forensic Acarology. In Fundamentals of Applied Acarology; Springer: Singapore, 2016; pp. 441–448. [Google Scholar]
- Perotti, M.A. Mégnin re-analysed: The case of the newborn baby girl, Paris, 1878. Exp. Appl. Acarol. 2009, 49, 37–44. [Google Scholar] [CrossRef]
- Krantz, G.W. Reflections on the biology, morphology and ecology of the Macrochelidae. Rev. Lit. Arts Am. 1998, 22, 125–137. [Google Scholar]
- Bahrami, F.; Arbabi, M.; Shoushtari, R.V.; Kazemi, S. Mesostigmatic Mites Associated with Coleoptera and Biodiversity Calculation of These Mites Phoretic on Dung Beetles in Golestan Province (North of Iran). Middle-East J. Sci. Res. 2011, 9, 345–366. [Google Scholar]
- Goff, M.L. Gamasid mites as potential indicators of postmortem interval. In Progress in Acarology Volume; Channabasavanna, G., Viraktamatch, C., Eds.; Oxford and IBH Publishing: New Delhi, India, 1989; Volume 1, pp. 443–450. [Google Scholar]
- Niogret, J.; Lumaret, J.; Bertrand, M. Review of the phoretic association between coprophilous insects and Macrochelid mites (Acari: Mesostigmata) in France. Elytron 2008, 20, 99–121. [Google Scholar]
- Bloszyk, J.; Bajerlein, D.; Blaszak, C. The use of pedicels of phoretic deutonymph of Uropoda orbicularis (Acari: Uropodidae) connected with coprophagous beetles (Insecta: Coleoptera) by Macrocheles. Pol. J. Entomol. 2002, 71, 241–246. [Google Scholar]
- Evans, G.O.; Browning, E. British mites of the subfamily Macrochelinae Trägardh (Gamasina-Macrochelidae). Bull. Br. Mus. (Nat. Hist. Zool.) 1956, 4, 1–56. [Google Scholar]
- Costa, M. The mesostigmatic mites associated with Copris hispanus (L.) (Coleoptera, Scarabaeidae) in Israel. J. Linn. Soc. Lond. Zool. 1963, 45, 25–45. [Google Scholar] [CrossRef]
- Axtell, R.C. Phoretic Relationship of Some Common Manure-Inhabiting Macrochelidae (Acarina: Mesostigmata) to the House Fly. Ann. Entomol. Soc. Am. 1964, 57, 584–587. [Google Scholar] [CrossRef]
- Rodrigueiro, T.S.C.; do Prado, A.P. Macrocheles muscaedomesticae (Acari, Macrochelidae) and a species of Uroseius (Acari, Polyaspididae) phoretic on Musca domestica (Diptera, Muscidae): Effects on dispersal and colonization of poultry manure. Iheringia Série Zool. 2004, 94, 181–185. [Google Scholar] [CrossRef]
- Perotti, A.; Brasesco, M. Especificidad forética de Macrocheles muscaedomesticae (Acari: Macrochelidae). Ecol. Austral 1996, 6, 3–8. [Google Scholar]
- Marquardt, T.; Kaczmarek, S.; Halliday, B. Ovoviviparity in Macrocheles glaber (Müller) (Acari: Macrochelidae), with notes on parental care and egg cannibalism. Int. J. Acarol. 2015, 41, 71–76. [Google Scholar] [CrossRef]
- Filipponi, A.; Francaviglia, G. Larviparitá facoltativa in alcuni Macrochelidi (Acari: Mesostigmata) associati a muscidi di interesse sanitario. Parasitologia 1964, 6, 99–113. [Google Scholar]
- Kamaruzaman, N.A.C.; Mašán, P.; Velásquez, Y.; González-Medina, A.; Lindström, A.; Braig, H.R.; Perotti, M.A. Macrocheles species (Acari: Macrochelidae) associated with human corpses in Europe. Exp. Appl. Acarol. 2018, 1–19. [Google Scholar] [CrossRef]
- Samsinak, K. Mites on flies of the family Sphaeroceridae. II. Acarologia 1989, 30, 85–105. [Google Scholar]
- Lundqvist, L. Phoretic Gamasina (Acari) from Southern Sweden: Taxonomy, host preferences and seasonality. Acarologia 1998, 39, 111–114. [Google Scholar]
- Springett, B.P. Aspects of the relationship between burying beetles, Necrophorus spp. and the mites, Poecilochirus necrophori Vitz. J. Anim. Ecol. 1968, 37, 417–424. [Google Scholar] [CrossRef]
- Schwarz, H.H.; Müller, J.K. The dispersal behaviour of the phoretic mite Poecilochirus carabi (Mesostigmata, Parasitidae): Adaptation to the breeding biology of its carrier Necrophorus vespilloides (Coleoptera, Silphidae). Oecologia 1992, 89, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Wilson, D.S. Local specialization of phoretic mites on sympatric carrion beetle hosts. Ecology 1992, 73, 463–478. [Google Scholar] [CrossRef]
- Blackman, S.W.; Evans, G.O. Observations on a mite (Poecilochirus davydovae) predatory on the eggs of burying beetles (Nicrophorus vespilloides) with a review of its taxonomic status. J. Zool. 1994, 234, 217–227. [Google Scholar] [CrossRef]
- Schwarz, H.H.; Walzl, M.G. Pairing, oviposition and development in two sibling species of phoretic mites (Acari: Mesostigmata: Parasitidae: Poecilochirus spp.) associated with burying beetles (Coleoptera: Silphidae: Nicrophorus spp.). J. Nat. Hist. 1996, 30, 1337–1348. [Google Scholar] [CrossRef]
- Baker, A.S.; Schwarz, H.H. Morphological differences between sympatric populations of the Poecilochirus carabi complex (Acari: Mesostigmata: Parasitidae) associated with burying beetles (Silphidae: Nicrophorus). Syst. Parasitol. 1997, 37, 179–185. [Google Scholar] [CrossRef]
- Salmane, I.; Telnov, D. Mesostigmata Mites (Acari: Parasitiformes) Associated with Beetles (Insecta: Coleoptera) in Latvia. Latvijas Entomol. 2009, 47, 58–70. [Google Scholar]
- González Medina, A.; González Herrera, L.; Perotti, M.A.; Jiménez Ríos, G. Occurrence of Poecilochirus austroasiaticus (Acari: Parasitidae) in forensic autopsies and its application on postmortem interval estimation. Exp. Appl. Acarol. 2013, 59, 297–305. [Google Scholar] [CrossRef]
- Barton, P.S.; Weaver, H.J.; Manning, A.D. Contrasting diversity dynamics of phoretic mites and beetles associated with vertebrate carrion. Exp. Appl. Acarol. 2014, 63, 1–13. [Google Scholar] [CrossRef] [PubMed]
- García-Guerrero, D.A.; Monjarás-Barrera, J.I.; Hernandez-Juarez, A.; Aguirre-Uribe, L.A. Asociación del ácaro Poecilochirus sp. (Acari: Parasitidae) y Nicrophorus marginatus (Coleoptera: Silphidae) en Saltillo, Coahuila, México. Entomol. Mex. 2014, 1, 69–73. [Google Scholar]
- Houck, M.A.; OConnor, B.M. Ecological and Evolutionary Significance of Phoresy in the Astigmata. Annu. Rev. Entomol. 1991, 36, 611–636. [Google Scholar] [CrossRef]
- Moraza, M.L.; Hernández, M.A. Mesostigmata mite community in two Japanese larch (Larix kaempferi) plantations and an oak (Quercus pyrenaica) forest in Navarra (northern Spain). Rev. Ibérica Aracnol. 2016, 28, 35–44. [Google Scholar]
- Christie, J.E. A new species of Alliphis (Mesostigmata: Eviphididae) from Britain. Acarologia 1983, 24, 231–242. [Google Scholar]
- Makarova, G.L. Gamasid Mites of the Genus Crassicheles (Mesostigmata, Eviphididae) in Russia and Ontogenesis of Crassicheles greeni. Entomol. Rev. 1994, 73, 107–117. [Google Scholar]
- Mašán, P.; Perotti, M.A.; Saloña-Bordas, M.I.; Braig, H.R. Proctolaelaps euserratus, an ecologically unusual melicharid mite (Acari, Mesostigmata) associated with animal and human decomposition. Exp. Appl. Acarol. 2013, 61, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Moraza, M.L. Efecto de la degradación de un encinar de Quercus rotundifolia en la comunidad de ácaros cryptostigmados y mesostigmados (Acari: Cryptostigmata, Mesostigmata). Rev. Ibérica Aracnol. 2007, 13, 171–182. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sp. No. | Family | Species | No. | Stage | Trap |
|---|---|---|---|---|---|
| 1 | Parasitidae | Cornigamasus lunaris (Berlese, 1882) | 2 | DN | C, L |
| 2 | Gamasodes spiniger (Trägårdh, 1910) | 34 | DN | C, D, L | |
| 3 | Parasitus (Coleogamasus) americanus (Berlese, 1906) | 1 | ♀ e | C | |
| 4 | Parasitus coleoptratorum (Linneaus, 1758) | 38 | ♂, DN | C, L | |
| 5 | Parasitus consanguineus Oudemans & Voigts, 1904 | 2 | DN | L | |
| 6 | Parasitus fimetorum (Berlese, 1903) | 20 | DN | C, L | |
| 7 | Pergamasus sp. | 2 | ♀, ♂ | C | |
| 8 | Poecilochirus austroasiaticus Vitzthum, 1930 | 111 | DN | C, L | |
| 9 | Poecilochirus carabi G. & R. Canestrini, 1882 | 91 | DN | C, L | |
| 10 | Poecilochirus subterraneus (Müller, 1859) | 268 | DN | C, L | |
| 11 | Vulgarogamasus remberti (Oudemans, 1912) | 1 | DN | C | |
| 12 | Veigaiidae | Veigaia planicola (Berlese, 1892) | 1 | ♀ | L |
| 13 | Macrochelidae | Glyptholaspis confusa (Foá, 1900) | 1 | ♀ | L |
| 14 | Macrocheles glaber (Müller, 1860) | 45 | ♀, ♀ e, ♀ Lv | L | |
| 15 | Macrocheles merdarius (Berlese, 1889) | 116 | ♀, ♀ e, ♀ Lv | C, L | |
| 16 | Macrocheles muscaedomesticae (Scopoli, 1772) | 27 | ♀ e, ♀ Lv | D, C | |
| 17 | Eviphididae | Alliphis kargi Arutunian, 1991 | 1 | ♀ | C |
| 18 | Alliphis necrophilus Christie, 1983 | 1 | ♀ | L | |
| 19 | Crassicheles holsaticus Willm, 1937 | 21 | DN | C, L | |
| 20 | Scarabaspis inexpectatus (Oudemans, 1903) | 3 | ♀, DN | L | |
| 21 | Ascidae | Proctolaelaps sp. | 7 | ♀ | L |
| 22 | Halolaelapidae | Halolaelaps sp. | 1 | DN | C |
| 23 | Halolaelaps octoclavatus (Vitzthum, 1920) | 34 | DN | C, L | |
| 24 | Laelapidae | Cosmolaelaps vacua (Michael, 1891) | 2 | ♀ | C |
| 25 | Cosmolaelaps lutegiensis (Shcherbak, 1971) | 12 | ♀, ♀ e | C | |
| 26 | Hypoaspis (Gaeolaelaps) sp. | 2 | ♀ | C | |
| Total collected: | 844 |
| Sp. No. | 11 April | 24 April | 29 April | 7 May | 13 May | 21 May | 27 May | 4 June | 10 June | 17 June | 24 June |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | • | ||||||||||
| 2 | • | • | • | • | • | • | • | • | |||
| 3 | • | ||||||||||
| 4 | • | • | • | • | • | ||||||
| 5 | • | • | |||||||||
| 6 | • | • | • | • | • | ||||||
| 7 | |||||||||||
| 8 | |||||||||||
| 9 | |||||||||||
| 10 | |||||||||||
| 11 | |||||||||||
| 12 | |||||||||||
| 13 | |||||||||||
| 14 | |||||||||||
| 15 | |||||||||||
| 16 | |||||||||||
| 17 | |||||||||||
| 18 | |||||||||||
| 19 | |||||||||||
| 20 | |||||||||||
| 21 | |||||||||||
| 22 | |||||||||||
| 23 | |||||||||||
| 24 | |||||||||||
| 25 | |||||||||||
| 26 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Martínez, S.; Moraza, M.L.; Saloña-Bordas, M.I. Gamasina Mites (Acari: Mesostigmata) Associated with Animal Remains in the Mediterranean Region of Navarra (Northern Spain). Insects 2019, 10, 5. https://doi.org/10.3390/insects10010005
Pérez-Martínez S, Moraza ML, Saloña-Bordas MI. Gamasina Mites (Acari: Mesostigmata) Associated with Animal Remains in the Mediterranean Region of Navarra (Northern Spain). Insects. 2019; 10(1):5. https://doi.org/10.3390/insects10010005
Chicago/Turabian StylePérez-Martínez, Sandra, María Lourdes Moraza, and Marta Inés Saloña-Bordas. 2019. "Gamasina Mites (Acari: Mesostigmata) Associated with Animal Remains in the Mediterranean Region of Navarra (Northern Spain)" Insects 10, no. 1: 5. https://doi.org/10.3390/insects10010005