
The Role of BDNF as a Biomarker in Cognitive and Sensory Neurodegeneration

 , , , , , and
, , , , , and 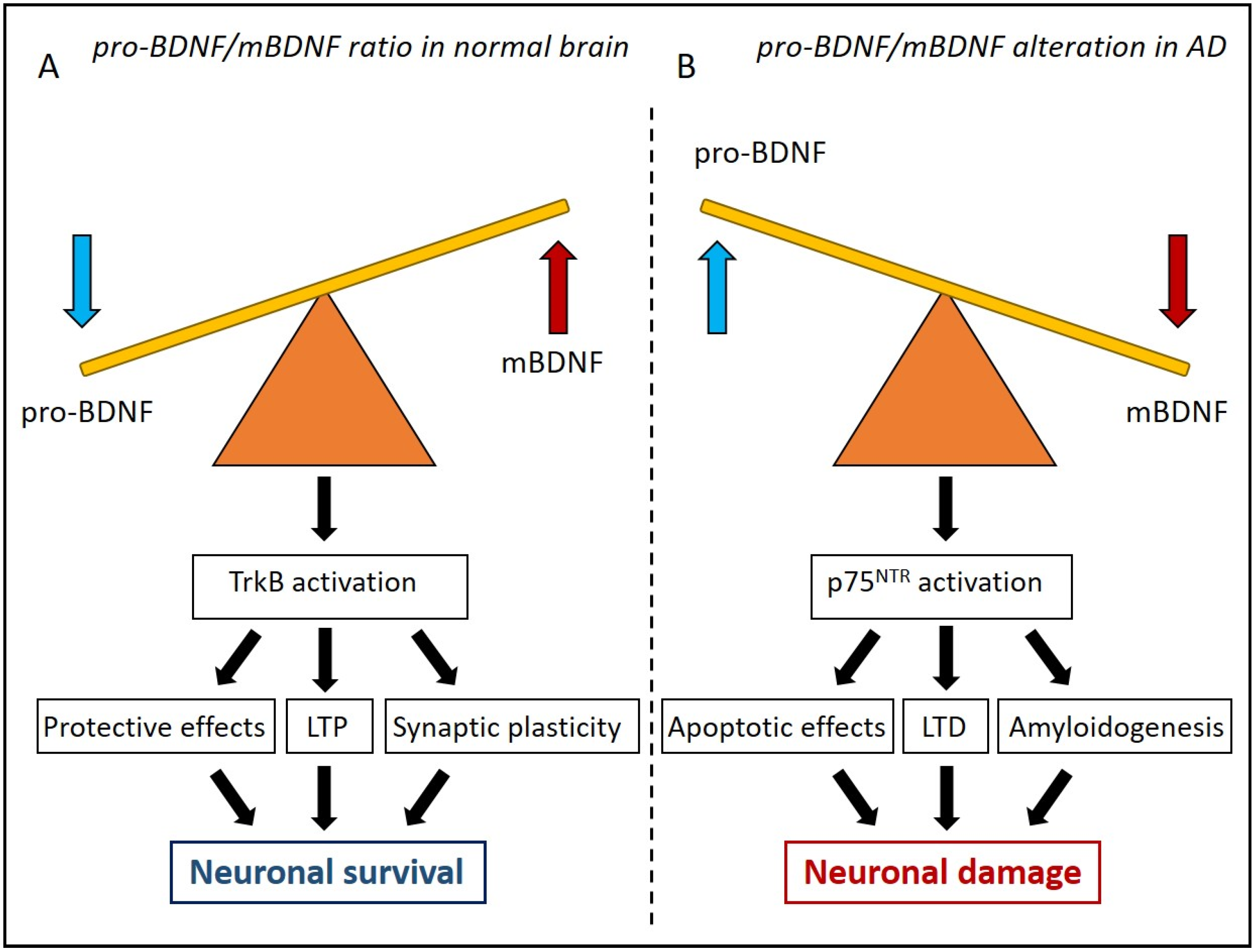{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Antagonist Role of Pro-BDNF and mBDNF
3. BDNF in the Brain: Its Role in Learning and Memory Processes
Role of BDNF as an Early Biomarker in Neurodegenerative Disorders
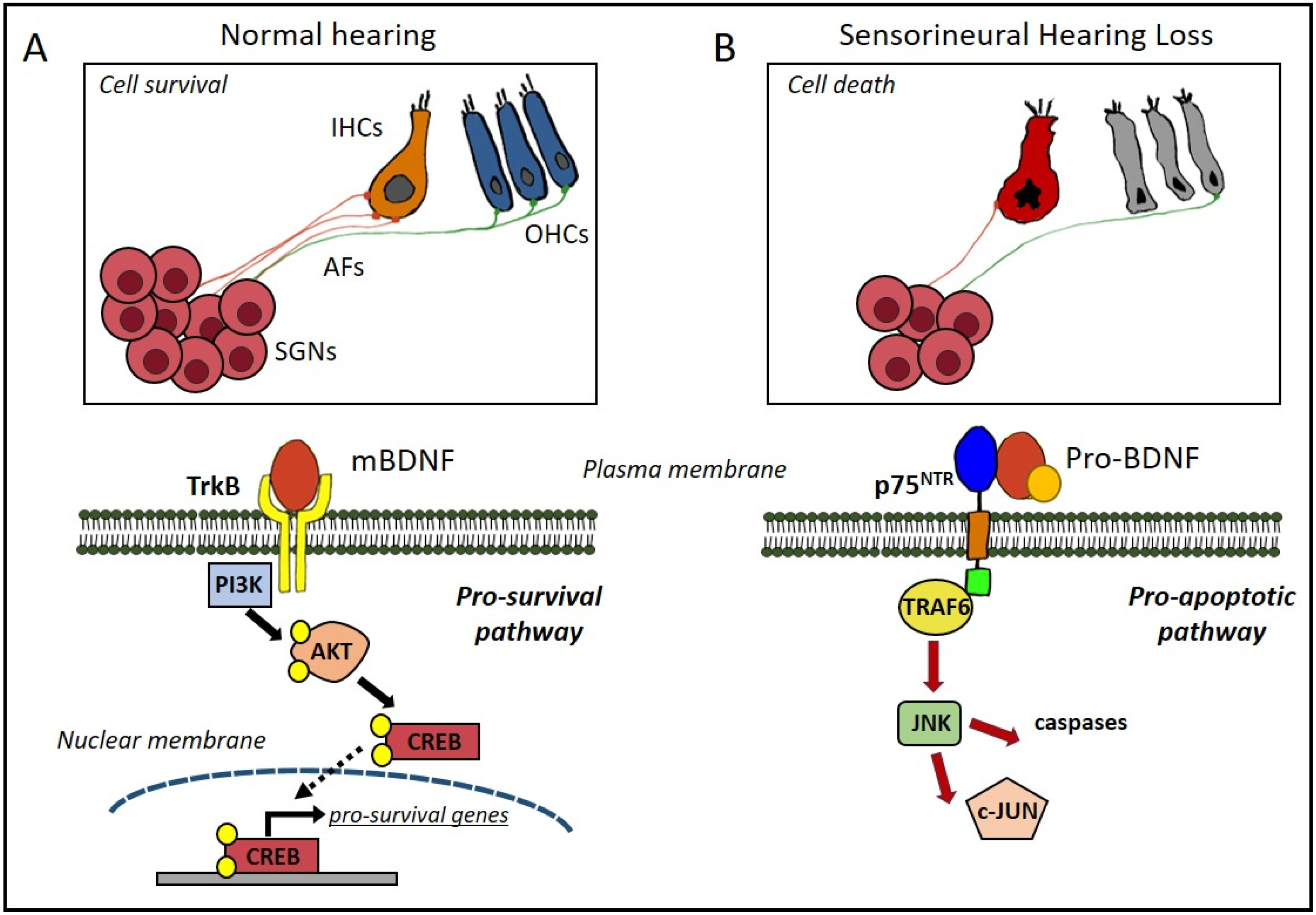
4. BDNF in the Auditory System
4.1. Cochlear Damage: Sensorineural Hearing Loss
4.2. BDNF Protective Effects against Hearing Loss
5. The Role of BDNF in Olfaction
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, G.R.; Barde, Y.A. Physiology of the Neurotrophins. Annu. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Maisonpierre, P.C.; Belluscio, L.; Friedman, B.; Alderson, R.F.; Wiegand, S.J.; Furth, M.E.; Lindsay, R.M.; Yancopoulos, G.D. NT-3, BDNF, and NGF in the Developing Rat Nervous System: Parallel as Well as Reciprocal Patterns of Expression. Neuron 1990, 5, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, J.C.; Wu, S.H. Neurotrophin Signaling: Many Exciting Surprises! Cell. Mol. Life Sci. CMLS 2006, 63, 1523–1537. [Google Scholar] [CrossRef]
- Biffo, S.; Offenhäuser, N.; Carter, B.D.; Barde, Y.A. Selective Binding and Internalisation by Truncated Receptors Restrict the Availability of BDNF during Development. Dev. Camb. Engl. 1995, 121, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Martinowich, K.; Lu, B. Interaction between BDNF and Serotonin: Role in Mood Disorders. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2008, 33, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Tsuji, M.; Oguchi, T.; Kasuga, K.; Kimura, A.; Futamura, A.; Sugimoto, A.; Kasai, H.; Kuroda, T.; Yano, S.; et al. Serum BDNF as a Potential Biomarker of Alzheimer’s Disease: Verification Through Assessment of Serum, Cerebrospinal Fluid, and Medial Temporal Lobe Atrophy. Front. Neurol. 2021, 12, 653267. [Google Scholar] [CrossRef] [PubMed]
- Malekan, M.; Nezamabadi, S.S.; Samami, E.; Mohebalizadeh, M.; Saghazadeh, A.; Rezaei, N. BDNF and Its Signaling in Cancer. J. Cancer Res. Clin. Oncol. 2022. [Google Scholar] [CrossRef]
- Leal, G.; Afonso, P.M.; Salazar, I.L.; Duarte, C.B. Regulation of Hippocampal Synaptic Plasticity by BDNF. Brain Res. 2015, 1621, 82–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yao, W.; Hashimoto, K. Brain-Derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-Related Depression and Potential Therapeutic Targets. Curr. Neuropharmacol. 2016, 14, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Johnson Chacko, L.; Blumer, M.J.F.; Pechriggl, E.; Rask-Andersen, H.; Dietl, W.; Haim, A.; Fritsch, H.; Glueckert, R.; Dudas, J.; Schrott-Fischer, A. Role of BDNF and Neurotrophic Receptors in Human Inner Ear Development. Cell Tissue Res. 2017, 370, 347–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappoli, N.; Tabolacci, E.; Aceto, P.; Dello Russo, C. The Emerging Role of the BDNF-TrkB Signaling Pathway in the Modulation of Pain Perception. J. Neuroimmunol. 2020, 349, 577406. [Google Scholar] [CrossRef]
- Dincheva, I.; Lynch, N.B.; Lee, F.S. The Role of BDNF in the Development of Fear Learning. Depress. Anxiety 2016, 33, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.Z.; Nusslock, R. Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front. Neurosci. 2018, 12, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, W.A.; Jiang, H.; Tartaglia, N.; Feng, L.; Figurov, A.; Lu, B. Signaling Mechanisms Mediating BDNF Modulation of Synaptic Plasticity in the Hippocampus. Learn. Mem. 1999, 6, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Constantine-Paton, M. Postsynaptic BDNF-TrkB Signaling in Synapse Maturation, Plasticity, and Disease. Dev. Neurobiol. 2010, 70, 304–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S.S.; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotox. Res. 2019, 35, 775–795. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Chen, C.-C.; Hung, T.-H.; Chuang, Y.-C.; Chao, M.; Shyue, S.-K.; Chen, S.-F. Activation of TrkB/Akt Signaling by a TrkB Receptor Agonist Improves Long-Term Histological and Functional Outcomes in Experimental Intracerebral Hemorrhage. J. Biomed. Sci. 2019, 26, 53. [Google Scholar] [CrossRef] [Green Version]
- Arancibia, S.; Silhol, M.; Moulière, F.; Meffre, J.; Höllinger, I.; Maurice, T.; Tapia-Arancibia, L. Protective Effect of BDNF against Beta-Amyloid Induced Neurotoxicity in Vitro and in Vivo in Rats. Neurobiol. Dis. 2008, 31, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Shepherd, R.K. Aminoglycoside-Induced Degeneration of Adult Spiral Ganglion Neurons Involves Differential Modulation of Tyrosine Kinase B and P75 Neurotrophin Receptor Signaling. Am. J. Pathol. 2006, 169, 528–543. [Google Scholar] [CrossRef] [Green Version]
- Mowla, S.J.; Farhadi, H.F.; Pareek, S.; Atwal, J.K.; Morris, S.J.; Seidah, N.G.; Murphy, R.A. Biosynthesis and Post-Translational Processing of the Precursor to Brain-Derived Neurotrophic Factor. J. Biol. Chem. 2001, 276, 12660–12666. [Google Scholar] [CrossRef] [Green Version]
- Hempstead, B.L. Brain-Derived Neurotrophic Factor: Three Ligands, Many Actions. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 9–19. [Google Scholar]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of Cell Survival by Secreted Proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Pang, P.T.; Woo, N.H. The Yin and Yang of Neurotrophin Action. Nat. Rev. Neurosci. 2005, 6, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffino, L.; Mottarlini, F.; Fumagalli, F. Born to Protect: Leveraging BDNF Against Cognitive Deficit in Alzheimer’s Disease. CNS Drugs 2020, 34, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Pang, P.T.; Teng, H.K.; Zaitsev, E.; Woo, N.T.; Sakata, K.; Zhen, S.; Teng, K.K.; Yung, W.-H.; Hempstead, B.L.; Lu, B. Cleavage of ProBDNF by TPA/Plasmin Is Essential for Long-Term Hippocampal Plasticity. Science 2004, 306, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Woo, N.H.; Teng, H.K.; Siao, C.-J.; Chiaruttini, C.; Pang, P.T.; Milner, T.A.; Hempstead, B.L.; Lu, B. Activation of P75NTR by ProBDNF Facilitates Hippocampal Long-Term Depression. Nat. Neurosci. 2005, 8, 1069–1077. [Google Scholar] [CrossRef]
- Gentry, J.J.; Barker, P.A.; Carter, B.D. The P75 Neurotrophin Receptor: Multiple Interactors and Numerous Functions. Prog. Brain Res. 2004, 146, 25–39. [Google Scholar] [CrossRef]
- Teng, H.K.; Teng, K.K.; Lee, R.; Wright, S.; Tevar, S.; Almeida, R.D.; Kermani, P.; Torkin, R.; Chen, Z.-Y.; Lee, F.S.; et al. ProBDNF Induces Neuronal Apoptosis via Activation of a Receptor Complex of P75NTR and Sortilin. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 5455–5463. [Google Scholar] [CrossRef]
- Volosin, M.; Song, W.; Almeida, R.D.; Kaplan, D.R.; Hempstead, B.L.; Friedman, W.J. Interaction of Survival and Death Signaling in Basal Forebrain Neurons: Roles of Neurotrophins and Proneurotrophins. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 7756–7766. [Google Scholar] [CrossRef] [Green Version]
- Friedman, W.J. Proneurotrophins, Seizures, and Neuronal Apoptosis. Neuroscientist 2010, 16, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Poo, M.M. Neurotrophins as Synaptic Modulators. Nat. Rev. Neurosci. 2001, 2, 24–32. [Google Scholar] [CrossRef]
- Boyd, J.G.; Gordon, T. The Neurotrophin Receptors, TrkB and P75, Differentially Regulate Motor Axonal Regeneration. J. Neurobiol. 2001, 49, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Avwenagha, O.; Campbell, G.; Bird, M.M. The Outgrowth Response of the Axons of Developing and Regenerating Rat Retinal Ganglion Cells In Vitro to Neurotrophin Treatment. J. Neurocytol. 2003, 32, 1055–1075. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Li, Q.; Hempstead, B.L.; Madri, J.A. Paracrine and Autocrine Functions of Brain-Derived Neurotrophic Factor (BDNF) and Nerve Growth Factor (NGF) in Brain-Derived Endothelial Cells. J. Biol. Chem. 2004, 279, 33538–33546. [Google Scholar] [CrossRef] [Green Version]
- Lessmann, V.; Gottmann, K.; Malcangio, M. Neurotrophin Secretion: Current Facts and Future Prospects. Prog. Neurobiol. 2003, 69, 341–374. [Google Scholar] [CrossRef]
- McAllister, A.K. Subplate Neurons: A Missing Link among Neurotrophins, Activity, and Ocular Dominance Plasticity? Proc. Natl. Acad. Sci. USA 1999, 96, 13600–13602. [Google Scholar] [CrossRef] [Green Version]
- Zagrebelsky, M.; Holz, A.; Dechant, G.; Barde, Y.-A.; Bonhoeffer, T.; Korte, M. The P75 Neurotrophin Receptor Negatively Modulates Dendrite Complexity and Spine Density in Hippocampal Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 9989–9999. [Google Scholar] [CrossRef] [Green Version]
- Bibel, M.; Hoppe, E.; Barde, Y.A. Biochemical and Functional Interactions between the Neurotrophin Receptors Trk and P75NTR. EMBO J. 1999, 18, 616–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, T.; Palm, K.; Metsis, M.; Reintam, T.; Paalme, V.; Saarma, M.; Persson, H. Multiple Promoters Direct Tissue-Specific Expression of the Rat BDNF Gene. Neuron 1993, 10, 475–489. [Google Scholar] [CrossRef]
- Chen, W.G.; Chang, Q.; Lin, Y.; Meissner, A.; West, A.E.; Griffith, E.C.; Jaenisch, R.; Greenberg, M.E. Derepression of BDNF Transcription Involves Calcium-Dependent Phosphorylation of MeCP2. Science 2003, 302, 885–889. [Google Scholar] [CrossRef]
- Metsis, M.; Timmusk, T.; Arenas, E.; Persson, H. Differential Usage of Multiple Brain-Derived Neurotrophic Factor Promoters in the Rat Brain Following Neuronal Activation. Proc. Natl. Acad. Sci. USA 1993, 90, 8802–8806. [Google Scholar] [CrossRef] [Green Version]
- Oliff, H.S.; Berchtold, N.C.; Isackson, P.; Cotman, C.W. Exercise-Induced Regulation of Brain-Derived Neurotrophic Factor (BDNF) Transcripts in the Rat Hippocampus. Brain Res. Mol. Brain Res. 1998, 61, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Lagopoulos, J. Stress and Trauma: BDNF Control of Dendritic-Spine Formation and Regression. Prog. Neurobiol. 2014, 112, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Rizavi, H.S.; Pandey, G.N. Antidepressants Reverse Corticosterone-Mediated Decrease in Brain-Derived Neurotrophic Factor Expression: Differential Regulation of Specific Exons by Antidepressants and Corticosterone. Neuroscience 2006, 139, 1017–1029. [Google Scholar] [CrossRef] [Green Version]
- Donnici, L.; Tiraboschi, E.; Tardito, D.; Musazzi, L.; Racagni, G.; Popoli, M. Time-Dependent Biphasic Modulation of Human BDNF by Antidepressants in Neuroblastoma Cells. BMC Neurosci. 2008, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Sohrabji, F.; Lewis, D.K. Estrogen-BDNF Interactions: Implications for Neurodegenerative Diseases. Front. Neuroendocrinol. 2006, 27, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.C.; Hill, R.A.; Gogos, A.; van den Buuse, M. Sex Differences and the Role of Estrogen in Animal Models of Schizophrenia: Interaction with BDNF. Neuroscience 2013, 239, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Boulle, F.; van den Hove, D.L.A.; Jakob, S.B.; Rutten, B.P.; Hamon, M.; van Os, J.; Lesch, K.-P.; Lanfumey, L.; Steinbusch, H.W.; Kenis, G. Epigenetic Regulation of the BDNF Gene: Implications for Psychiatric Disorders. Mol. Psychiatry 2012, 17, 584–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.M.; Brown, A.N.; Bhide, P.G. Regulation of BDNF Expression by Cocaine. Yale J. Biol. Med. 2012, 85, 437–446. [Google Scholar]
- Li, Y.; Luikart, B.W.; Birnbaum, S.; Chen, J.; Kwon, C.-H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB Regulates Hippocampal Neurogenesis and Governs Sensitivity to Antidepressive Treatment. Neuron 2008, 59, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Liao, J.; Qi, F.; Meng, Z.; Pan, S. Evidence for the Contribution of BDNF-TrkB Signal Strength in Neurogenesis: An Organotypic Study. Neurosci. Lett. 2015, 606, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.F.; Ribeiro, F.F.; Rodrigues, R.S.; Sebastião, A.M.; Xapelli, S. Brain-Derived Neurotrophic Factor (BDNF) Role in Cannabinoid-Mediated Neurogenesis. Front. Cell. Neurosci. 2018, 12, 441. [Google Scholar] [CrossRef] [Green Version]
- Vitaliano, G.D.; Kim, J.K.; Kaufman, M.J.; Adam, C.W.; Zeballos, G.; Shanmugavadivu, A.; Subburaju, S.; McLaughlin, J.P.; Lukas, S.E.; Vitaliano, F. Clathrin-Nanoparticles Deliver BDNF to Hippocampus and Enhance Neurogenesis, Synaptogenesis and Cognition in HIV/NeuroAIDS Mouse Model. Commun. Biol. 2022, 5, 236. [Google Scholar] [CrossRef] [PubMed]
- Alsina, B.; Vu, T.; Cohen-Cory, S. Visualizing Synapse Formation in Arborizing Optic Axons in Vivo: Dynamics and Modulation by BDNF. Nat. Neurosci. 2001, 4, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A Simple Role for BDNF in Learning and Memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Edelmann, E.; Lessmann, V.; Brigadski, T. Pre- and Postsynaptic Twists in BDNF Secretion and Action in Synaptic Plasticity. Neuropharmacology 2014, 76, 610–627. [Google Scholar] [CrossRef] [PubMed]
- Dieni, S.; Matsumoto, T.; Dekkers, M.; Rauskolb, S.; Ionescu, M.S.; Deogracias, R.; Gundelfinger, E.D.; Kojima, M.; Nestel, S.; Frotscher, M.; et al. BDNF and Its Pro-Peptide Are Stored in Presynaptic Dense Core Vesicles in Brain Neurons. J. Cell Biol. 2012, 196, 775–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Siao, C.-J.; Nagappan, G.; Marinic, T.; Jing, D.; McGrath, K.; Chen, Z.-Y.; Mark, W.; Tessarollo, L.; Lee, F.S.; et al. Neuronal Release of ProBDNF. Nat. Neurosci. 2009, 12, 113–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasi, M.; Vignoli, B.; Canossa, M.; Blum, R. Neurobiology of Local and Intercellular BDNF Signaling. Pflugers Arch. 2017, 469, 593–610. [Google Scholar] [CrossRef] [Green Version]
- Stenovec, M.; Lasič, E.; Božić, M.; Bobnar, S.T.; Stout, R.F.; Grubišić, V.; Parpura, V.; Zorec, R. Ketamine Inhibits ATP-Evoked Exocytotic Release of Brain-Derived Neurotrophic Factor from Vesicles in Cultured Rat Astrocytes. Mol. Neurobiol. 2016, 53, 6882–6896. [Google Scholar] [CrossRef] [PubMed]
- Leschik, J.; Eckenstaler, R.; Endres, T.; Munsch, T.; Edelmann, E.; Richter, K.; Kobler, O.; Fischer, K.-D.; Zuschratter, W.; Brigadski, T.; et al. Prominent Postsynaptic and Dendritic Exocytosis of Endogenous BDNF Vesicles in BDNF-GFP Knock-in Mice. Mol. Neurobiol. 2019, 56, 6833–6855. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.E.; Billington, C.J.; Kotz, C.M.; Wang, C. The Lighter Side of BDNF. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1053–R1069. [Google Scholar] [CrossRef] [PubMed]
- Hing, B.; Sathyaputri, L.; Potash, J.B. A Comprehensive Review of Genetic and Epigenetic Mechanisms That Regulate BDNF Expression and Function with Relevance to Major Depressive Disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatr. Genet. 2018, 177, 143–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, A.E.; Pruunsild, P.; Timmusk, T. Neurotrophins: Transcription and Translation. Handb. Exp. Pharmacol. 2014, 220, 67–100. [Google Scholar] [CrossRef] [PubMed]
- Hofer, M.; Pagliusi, S.R.; Hohn, A.; Leibrock, J.; Barde, Y.A. Regional Distribution of Brain-Derived Neurotrophic Factor MRNA in the Adult Mouse Brain. EMBO J. 1990, 9, 2459–2464. [Google Scholar] [CrossRef]
- Brigadski, T.; Leßmann, V. BDNF: A Regulator of Learning and Memory Processes with Clinical Potential. E-Neuroforum 2014, 20, 1–11. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. Handb. Exp. Pharmacol. 2014, 220, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Nabeshima, T. Brain-Derived Neurotrophic Factor/TrkB Signaling in Memory Processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Mizuno, M.; Nabeshima, T. Role for Brain-Derived Neurotrophic Factor in Learning and Memory. Life Sci. 2002, 70, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Kuang, S.; Li, H.; Ran, D.; Yang, J. CAMP/PKA-CREB-BDNF Signaling Pathway in Hippocampus Mediates Cyclooxygenase 2-Induced Learning/Memory Deficits of Rats Subjected to Chronic Unpredictable Mild Stress. Oncotarget 2017, 8, 35558–35572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loprinzi, P.D.; Frith, E. A Brief Primer on the Mediational Role of BDNF in the Exercise-Memory Link. Clin. Physiol. Funct. Imaging 2019, 39, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, T.; Xue, H.; Wang, L.; Deng, L.; Xie, Y.; Bai, X.; Xin, D.; Yuan, H.; Qiu, J.; et al. Inhibition of Necroptosis Rescues SAH-Induced Synaptic Impairments in Hippocampus via CREB-BDNF Pathway. Front. Neurosci. 2018, 12, 990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuman, E.M. Neurotrophin Regulation of Synaptic Transmission. Curr. Opin. Neurobiol. 1999, 9, 105–109. [Google Scholar] [CrossRef]
- Lipsky, R.H.; Marini, A.M. Brain-Derived Neurotrophic Factor in Neuronal Survival and Behavior-Related Plasticity. Ann. N. Y. Acad. Sci. 2007, 1122, 130–143. [Google Scholar] [CrossRef]
- Koehl, M.; Abrous, D.N. A New Chapter in the Field of Memory: Adult Hippocampal Neurogenesis. Eur. J. Neurosci. 2011, 33, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Poo, M. Neurotrophin Regulation of Neural Circuit Development and Function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Minichiello, L. TrkB Signalling Pathways in LTP and Learning. Nat. Rev. Neurosci. 2009, 10, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, I.; Medina, J.H. Memory Formation: The Sequence of Biochemical Events in the Hippocampus and Its Connection to Activity in Other Brain Structures. Neurobiol. Learn. Mem. 1997, 68, 285–316. [Google Scholar] [CrossRef] [Green Version]
- Lisman, J.; Cooper, K.; Sehgal, M.; Silva, A.J. Memory Formation Depends on Both Synapse-Specific Modifications of Synaptic Strength and Cell-Specific Increases in Excitability. Nat. Neurosci. 2018, 21, 309–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliss, T.; Collingridge, G.; Morris, R. Synaptic Plasticity in the Hippocampus. In The Hippocampus Book; Oxford University Press: New York, NY, USA, 2007; pp. 343–474. ISBN 978-0-19-510027-3. [Google Scholar]
- Neves, G.; Cooke, S.F.; Bliss, T.V.P. Synaptic Plasticity, Memory and the Hippocampus: A Neural Network Approach to Causality. Nat. Rev. Neurosci. 2008, 9, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Voglis, G.; Tavernarakis, N. The Role of Synaptic Ion Channels in Synaptic Plasticity. EMBO Rep. 2006, 7, 1104–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruel-Jungerman, E.; Davis, S.; Laroche, S. Brain Plasticity Mechanisms and Memory: A Party of Four. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2007, 13, 492–505. [Google Scholar] [CrossRef]
- Kesslak, J.P.; So, V.; Choi, J.; Cotman, C.W.; Gomez-Pinilla, F. Learning Upregulates Brain-Derived Neurotrophic Factor Messenger Ribonucleic Acid: A Mechanism to Facilitate Encoding and Circuit Maintenance? Behav. Neurosci. 1998, 112, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Yamada, K.; Olariu, A.; Nawa, H.; Nabeshima, T. Involvement of Brain-Derived Neurotrophic Factor in Spatial Memory Formation and Maintenance in a Radial Arm Maze Test in Rats. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 7116–7121. [Google Scholar] [CrossRef] [Green Version]
- Cirulli, F.; Berry, A.; Chiarotti, F.; Alleva, E. Intrahippocampal Administration of BDNF in Adult Rats Affects Short-Term Behavioral Plasticity in the Morris Water Maze and Performance in the Elevated plus-Maze. Hippocampus 2004, 14, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Bekinschtein, P.; Cammarota, M.; Izquierdo, I.; Medina, J.H. BDNF and Memory Formation and Storage. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2008, 14, 147–156. [Google Scholar] [CrossRef]
- Heldt, S.A.; Stanek, L.; Chhatwal, J.P.; Ressler, K.J. Hippocampus-Specific Deletion of BDNF in Adult Mice Impairs Spatial Memory and Extinction of Aversive Memories. Mol. Psychiatry 2007, 12, 656–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, M.E.; Bucci, D.J. BDNF Expression in Perirhinal Cortex Is Associated with Exercise-Induced Improvement in Object Recognition Memory. Neurobiol. Learn. Mem. 2010, 94, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, C.K.; Kelly, Á.M. Differential BDNF Signaling in Dentate Gyrus and Perirhinal Cortex during Consolidation of Recognition Memory in the Rat. Hippocampus 2012, 22, 2127–2135. [Google Scholar] [CrossRef]
- Miranda, M.; Kent, B.A.; Morici, J.F.; Gallo, F.; Saksida, L.M.; Bussey, T.J.; Weisstaub, N.; Bekinschtein, P. NMDA Receptors and BDNF Are Necessary for Discrimination of Overlapping Spatial and Non-Spatial Memories in Perirhinal Cortex and Hippocampus. Neurobiol. Learn. Mem. 2018, 155, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Zuccato, C.; Cattaneo, E. Brain-Derived Neurotrophic Factor in Neurodegenerative Diseases. Nat. Rev. Neurol. 2009, 5, 311–322. [Google Scholar] [CrossRef]
- Hadoush, H.; Banihani, S.A.; Khalil, H.; Al-Qaisi, Y.; Al-Sharman, A.; Al-Jarrah, M. Dopamine, BDNF and Motor Function Postbilateral Anodal Transcranial Direct Current Stimulation in Parkinson’s Disease. Neurodegener. Dis. Manag. 2018, 8, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R. Recent Advances on the Role of Brain-Derived Neurotrophic Factor (BDNF) in Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 6827. [Google Scholar] [CrossRef]
- Keifer, J. Regulation of AMPAR Trafficking in Synaptic Plasticity by BDNF and the Impact of Neurodegenerative Disease. J. Neurosci. Res. 2022, 100, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Zarneshan, S.N.; Fakhri, S.; Khan, H. Targeting Akt/CREB/BDNF Signaling Pathway by Ginsenosides in Neurodegenerative Diseases: A Mechanistic Approach. Pharmacol. Res. 2022, 177, 106099. [Google Scholar] [CrossRef] [PubMed]
- Laske, C.; Stransky, E.; Leyhe, T.; Eschweiler, G.W.; Wittorf, A.; Richartz, E.; Bartels, M.; Buchkremer, G.; Schott, K. Stage-Dependent BDNF Serum Concentrations in Alzheimer’s Disease. J. Neural Transm. 2006, 113, 1217–1224. [Google Scholar] [CrossRef]
- Laske, C.; Stransky, E.; Leyhe, T.; Eschweiler, G.W.; Maetzler, W.; Wittorf, A.; Soekadar, S.; Richartz, E.; Koehler, N.; Bartels, M.; et al. BDNF Serum and CSF Concentrations in Alzheimer’s Disease, Normal Pressure Hydrocephalus and Healthy Controls. J. Psychiatr. Res. 2007, 41, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Arancibia, L.; Aliaga, E.; Silhol, M.; Arancibia, S. New Insights into Brain BDNF Function in Normal Aging and Alzheimer Disease. Brain Res. Rev. 2008, 59, 201–220. [Google Scholar] [CrossRef]
- Scalzo, P.; Kümmer, A.; Bretas, T.L.; Cardoso, F.; Teixeira, A.L. Serum Levels of Brain-Derived Neurotrophic Factor Correlate with Motor Impairment in Parkinson’s Disease. J. Neurol. 2010, 257, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yun, W.; Zhang, M.; Luo, W.; Zhou, X. Serum Concentration and Clinical Significance of Brain-Derived Neurotrophic Factor in Patients with Parkinson’s Disease or Essential Tremor. J. Int. Med. Res. 2018, 46, 1477–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahara, A.H.; Tuszynski, M.H. Potential Therapeutic Uses of BDNF in Neurological and Psychiatric Disorders. Nat. Rev. Drug Discov. 2011, 10, 209–219. [Google Scholar] [CrossRef]
- Giralt, A.; Carretón, O.; Lao-Peregrin, C.; Martín, E.D.; Alberch, J. Conditional BDNF Release under Pathological Conditions Improves Huntington’s Disease Pathology by Delaying Neuronal Dysfunction. Mol. Neurodegener. 2011, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Weissmiller, A.M.; Wu, C. Current Advances in Using Neurotrophic Factors to Treat Neurodegenerative Disorders. Transl. Neurodegener. 2012, 1, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.; Manach, Y.L.; Riou, B.; Houle, T.T.; Warner, D.S. Statistical Evaluation of a Biomarker. Anesthesiology 2010, 112, 1023–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballman, K.V. Biomarker: Predictive or Prognostic. J. Clin. Oncol. 2015, 33, 3968–3971. [Google Scholar] [CrossRef]
- Califf, R.M. Biomarker Definitions and Their Applications. Exp. Biol. Med. Maywood N. J. 2018, 243, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Lima Giacobbo, B.; Doorduin, J.; Klein, H.C.; Dierckx, R.A.J.O.; Bromberg, E.; de Vries, E.F.J. Brain-Derived Neurotrophic Factor in Brain Disorders: Focus on Neuroinflammation. Mol. Neurobiol. 2019, 56, 3295–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, E.; Emiliano, A.; Maudsley, S.; Windham, B.G.; Carlson, O.D.; Egan, J.M.; Driscoll, I.; Ferrucci, L.; Martin, B.; Mattson, M.P. Circulating Brain-Derived Neurotrophic Factor and Indices of Metabolic and Cardiovascular Health: Data from the Baltimore Longitudinal Study of Aging. PLoS ONE 2010, 5, e10099. [Google Scholar] [CrossRef] [PubMed]
- Karantali, E.; Kazis, D.; Papavasileiou, V.; Prevezianou, A.; Chatzikonstantinou, S.; Petridis, F.; McKenna, J.; Luca, A.-C.; Trus, C.; Ciobica, A.; et al. Serum BDNF Levels in Acute Stroke: A Systematic Review and Meta-Analysis. Med. Kaunas Lith. 2021, 57, 297. [Google Scholar] [CrossRef]
- Caldieraro, M.A.; McKee, M.; Leistner-Segal, S.; Vares, E.A.; Kubaski, F.; Spanemberg, L.; Brusius-Facchin, A.C.; Fleck, M.P.; Mischoulon, D. Val66Met Polymorphism Association with Serum BDNF and Inflammatory Biomarkers in Major Depression. World J. Biol. Psychiatry 2018, 19, 402–409. [Google Scholar] [CrossRef]
- Bath, K.G.; Lee, F.S. Variant BDNF (Val66Met) Impact on Brain Structure and Function. Cogn. Affect. Behav. Neurosci. 2006, 6, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Pivac, N.; Kozaric-Kovacic, D.; Grubisic-Ilic, M.; Nedic, G.; Rakos, I.; Nikolac, M.; Blazev, M.; Muck-Seler, D. The Association between Brain-Derived Neurotrophic Factor Val66Met Variants and Psychotic Symptoms in Posttraumatic Stress Disorder. World J. Biol. Psychiatry 2012, 13, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Notaras, M.; Hill, R.; van den Buuse, M. The BDNF Gene Val66Met Polymorphism as a Modifier of Psychiatric Disorder Susceptibility: Progress and Controversy. Mol. Psychiatry 2015, 20, 916–930. [Google Scholar] [CrossRef] [PubMed]
- Notaras, M.; van den Buuse, M. Brain-Derived Neurotrophic Factor (BDNF): Novel Insights into Regulation and Genetic Variation. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2019, 25, 434–454. [Google Scholar] [CrossRef] [PubMed]
- Notaras, M.; van den Buuse, M. Neurobiology of BDNF in Fear Memory, Sensitivity to Stress, and Stress-Related Disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.; Budson, A. Current Understanding of Alzheimer’s Disease Diagnosis and Treatment. F1000Research 2018, 7. F1000 Faculty Rev-1161. [Google Scholar] [CrossRef] [Green Version]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Mol. Basel Switz. 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s Disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Boyle, P.A.; Wilson, R.S.; Aggarwal, N.T.; Tang, Y.; Bennett, D.A. Mild Cognitive Impairment: Risk of Alzheimer Disease and Rate of Cognitive Decline. Neurology 2006, 67, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Segawa, E.; Boyle, P.A.; Anagnos, S.E.; Hizel, L.P.; Bennett, D.A. The Natural History of Cognitive Decline in Alzheimer’s Disease. Psychol. Aging 2012, 27, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.; Xu, X.; You, Z.; Huang, Q.; Huang, Y.; Guo, Q.; Guan, Y.; Zhao, J.; Liu, J.; et al. In Vivo Synaptic Density Loss Correlates with Impaired Functional and Related Structural Connectivity in Alzheimer’s Disease. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2023, 271678X231153730. [Google Scholar] [CrossRef] [PubMed]
- De Paula, V.J.R.; Guimarães, F.M.; Diniz, B.S.; Forlenza, O.V. Neurobiological Pathways to Alzheimer’s Disease: Amyloid-Beta, TAU Protein or Both? Dement. Neuropsychol. 2009, 3, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.L.; Lee, V.M.-Y. Seeding of Normal Tau by Pathological Tau Conformers Drives Pathogenesis of Alzheimer-like Tangles. J. Biol. Chem. 2011, 286, 15317–15331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasagna-Reeves, C.A.; Castillo-Carranza, D.L.; Sengupta, U.; Guerrero-Munoz, M.J.; Kiritoshi, T.; Neugebauer, V.; Jackson, G.R.; Kayed, R. Alzheimer Brain-Derived Tau Oligomers Propagate Pathology from Endogenous Tau. Sci. Rep. 2012, 2, 700. [Google Scholar] [CrossRef] [Green Version]
- Kocahan, S.; Doğan, Z. Mechanisms of Alzheimer’s Disease Pathogenesis and Prevention: The Brain, Neural Pathology, N-Methyl-D-Aspartate Receptors, Tau Protein and Other Risk Factors. Clin. Psychopharmacol. Neurosci. Off. Sci. J. Korean Coll. Neuropsychopharmacol. 2017, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Butterfield, D.A. Role of Oxidative Stress in the Progression of Alzheimer’s Disease. J. Alzheimers Dis. JAD 2010, 19, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Bisht, K.; Sharma, K.; Tremblay, M.-È. Chronic Stress as a Risk Factor for Alzheimer’s Disease: Roles of Microglia-Mediated Synaptic Remodeling, Inflammation, and Oxidative Stress. Neurobiol. Stress 2018, 9, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Paciello, F.; Ripoli, C.; Fetoni, A.R.; Grassi, C. Redox Imbalance as a Common Pathogenic Factor Linking Hearing Loss and Cognitive Decline. Antioxidants 2023, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; Orellana, J.A.; von Bernhardi, R. Understanding Risk Factors for Alzheimer’s Disease: Interplay of Neuroinflammation, Connexin-Based Communication and Oxidative Stress. Arch. Med. Res. 2012, 43, 632–644. [Google Scholar] [CrossRef]
- Spires-Jones, T.L.; Hyman, B.T. The Intersection of Amyloid Beta and Tau at Synapses in Alzheimer’s Disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corey-Bloom, J. The ABC of Alzheimer’s Disease: Cognitive Changes and Their Management in Alzheimer’s Disease and Related Dementias. Int. Psychogeriatr. 2002, 14 (Suppl. S1), 51–75. [Google Scholar] [CrossRef]
- Jarrard, L.E. On the Role of the Hippocampus in Learning and Memory in the Rat. Behav. Neural Biol. 1993, 60, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Wittenberg, G.M.; Tsien, J.Z. An Emerging Molecular and Cellular Framework for Memory Processing by the Hippocampus. Trends Neurosci. 2002, 25, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Connor, B.; Young, D.; Yan, Q.; Faull, R.L.; Synek, B.; Dragunow, M. Brain-Derived Neurotrophic Factor Is Reduced in Alzheimer’s Disease. Brain Res. Mol. Brain Res. 1997, 49, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Phillips, H.S.; Hains, J.M.; Armanini, M.; Laramee, G.R.; Johnson, S.A.; Winslow, J.W. BDNF MRNA Is Decreased in the Hippocampus of Individuals with Alzheimer’s Disease. Neuron 1991, 7, 695–702. [Google Scholar] [CrossRef]
- Peng, S.; Wuu, J.; Mufson, E.J.; Fahnestock, M. Precursor Form of Brain-Derived Neurotrophic Factor and Mature Brain-Derived Neurotrophic Factor Are Decreased in the Pre-Clinical Stages of Alzheimer’s Disease. J. Neurochem. 2005, 93, 1412–1421. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, C.; Fabiani, C.; Bonadonna, C.; Origlia, N.; Domenici, L. BDNF Prevents Amyloid-Dependent Impairment of LTP in the Entorhinal Cortex by Attenuating P38 MAPK Phosphorylation. Neurobiol. Aging 2015, 36, 1303–1309. [Google Scholar] [CrossRef]
- Gibon, J.; Barker, P.A.; Séguéla, P. Opposing Presynaptic Roles of BDNF and ProBDNF in the Regulation of Persistent Activity in the Entorhinal Cortex. Mol. Brain 2016, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, I.; Marín, C.; Rey, M.J.; Ribalta, T.; Goutan, E.; Blanco, R.; Tolosa, E.; Martí, E. BDNF and Full-Length and Truncated TrkB Expression in Alzheimer Disease. Implications in Therapeutic Strategies. J. Neuropathol. Exp. Neurol. 1999, 58, 729–739. [Google Scholar] [CrossRef]
- Hock, C.; Heese, K.; Hulette, C.; Rosenberg, C.; Otten, U. Region-Specific Neurotrophin Imbalances in Alzheimer Disease: Decreased Levels of Brain-Derived Neurotrophic Factor and Increased Levels of Nerve Growth Factor in Hippocampus and Cortical Areas. Arch. Neurol. 2000, 57, 846–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahnestock, M.; Garzon, D.; Holsinger, R.M.D.; Michalski, B. Neurotrophic Factors and Alzheimer’s Disease: Are We Focusing on the Wrong Molecule? J. Neural Transm. Suppl. 2002, 241–252. [Google Scholar] [CrossRef]
- Garzon, D.; Yu, G.; Fahnestock, M. A New Brain-Derived Neurotrophic Factor Transcript and Decrease in Brain-Derived Neurotrophic Factor Transcripts 1, 2 and 3 in Alzheimer’s Disease Parietal Cortex. J. Neurochem. 2002, 82, 1058–1064. [Google Scholar] [CrossRef] [Green Version]
- Michalski, B.; Fahnestock, M. Pro-Brain-Derived Neurotrophic Factor Is Decreased in Parietal Cortex in Alzheimer’s Disease. Brain Res. Mol. Brain Res. 2003, 111, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Gezen-Ak, D.; Dursun, E.; Hanağası, H.; Bilgiç, B.; Lohman, E.; Araz, Ö.S.; Atasoy, İ.L.; Alaylıoğlu, M.; Önal, B.; Gürvit, H.; et al. BDNF, TNFα, HSP90, CFH, and IL-10 Serum Levels in Patients with Early or Late Onset Alzheimer’s Disease or Mild Cognitive Impairment. J. Alzheimers Dis. 2013, 37, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Pláteník, J.; Fišar, Z.; Buchal, R.; Jirák, R.; Kitzlerová, E.; Zvěřová, M.; Raboch, J. GSK3β, CREB, and BDNF in Peripheral Blood of Patients with Alzheimer’s Disease and Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 50, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wu, H.-T.; Qin, X.-Y.; Cao, C.; Liu, Y.; Cao, Z.-Z.; Cheng, Y. Postmortem Brain, Cerebrospinal Fluid, and Blood Neurotrophic Factor Levels in Alzheimer’s Disease: A Systematic Review and Meta-Analysis. J. Mol. Neurosci. MN 2018, 65, 289–300. [Google Scholar] [CrossRef]
- Korte, M.; Carroll, P.; Wolf, E.; Brem, G.; Thoenen, H.; Bonhoeffer, T. Hippocampal Long-Term Potentiation Is Impaired in Mice Lacking Brain-Derived Neurotrophic Factor. Proc. Natl. Acad. Sci. USA 1995, 92, 8856–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoletti, A.; Cancedda, L.; Reid, S.W.; Tessarollo, L.; Porciatti, V.; Pizzorusso, T.; Maffei, L. Heterozygous Knock-out Mice for Brain-Derived Neurotrophic Factor Show a Pathway-Specific Impairment of Long-Term Potentiation but Normal Critical Period for Monocular Deprivation. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 10072–10077. [Google Scholar] [CrossRef]
- Faria, M.C.; Gonçalves, G.S.; Rocha, N.P.; Moraes, E.N.; Bicalho, M.A.; Gualberto Cintra, M.T.; Jardim de Paula, J.; José Ravic de Miranda, L.F.; Clayton de Souza Ferreira, A.; Teixeira, A.L.; et al. Increased Plasma Levels of BDNF and Inflammatory Markers in Alzheimer’s Disease. J. Psychiatr. Res. 2014, 53, 166–172. [Google Scholar] [CrossRef]
- Shin, H.W.; Kim, H.; Lee, K.J. Differences in BDNF Serum Levels in Patients with Alzheimer’s Disease and Mild Cognitive Impairment. Afr. J Psychiatry 2015, 18, 245. [Google Scholar]
- Bharani, K.L.; Ledreux, A.; Gilmore, A.; Carroll, S.L.; Granholm, A.-C. Serum Pro-BDNF Levels Correlate with Phospho-Tau Staining in Alzheimer’s Disease. Neurobiol. Aging 2020, 87, 49–59. [Google Scholar] [CrossRef]
- Holsinger, R.M.; Schnarr, J.; Henry, P.; Castelo, V.T.; Fahnestock, M. Quantitation of BDNF MRNA in Human Parietal Cortex by Competitive Reverse Transcription-Polymerase Chain Reaction: Decreased Levels in Alzheimer’s Disease. Brain Res. Mol. Brain Res. 2000, 76, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Rabizadeh, S.; Bredesen, D.E. Ten Years on: Mediation of Cell Death by the Common Neurotrophin Receptor P75(NTR). Cytokine Growth Factor Rev. 2003, 14, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Nykjaer, A.; Lee, R.; Teng, K.K.; Jansen, P.; Madsen, P.; Nielsen, M.S.; Jacobsen, C.; Kliemannel, M.; Schwarz, E.; Willnow, T.E.; et al. Sortilin Is Essential for ProNGF-Induced Neuronal Cell Death. Nature 2004, 427, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Costantini, C.; Rossi, F.; Formaggio, E.; Bernardoni, R.; Cecconi, D.; Della-Bianca, V. Characterization of the Signaling Pathway Downstream P75 Neurotrophin Receptor Involved in Beta-Amyloid Peptide-Dependent Cell Death. J. Mol. Neurosci. 2005, 25, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Gerenu, G.; Martisova, E.; Ferrero, H.; Carracedo, M.; Rantamäki, T.; Ramirez, M.J.; Gil-Bea, F.J. Modulation of BDNF Cleavage by Plasminogen-Activator Inhibitor-1 Contributes to Alzheimer’s Neuropathology and Cognitive Deficits. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, T.; Jiao, S.; Zhou, X.; Zhong, J.; Wang, Y.; Liu, J.; Deng, J.; Wang, S.; Xu, Z. ProBDNF Accelerates Brain Amyloid-β Deposition and Learning and Memory Impairment in APPswePS1dE9 Transgenic Mice. J. Alzheimers Dis. 2017, 59, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Fleitas, C.; Piñol-Ripoll, G.; Marfull, P.; Rocandio, D.; Ferrer, I.; Rampon, C.; Egea, J.; Espinet, C. ProBDNF Is Modified by Advanced Glycation End Products in Alzheimer’s Disease and Causes Neuronal Apoptosis by Inducing P75 Neurotrophin Receptor Processing. Mol. Brain 2018, 11, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, M.T.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, TrkB-TK+ and GAD67 MRNA Expression in the Hippocampus of Individuals with Schizophrenia and Mood Disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Carlo, P.; Punzi, G.; Ursini, G. BDNF and Schizophrenia. Psychiatr. Genet. 2019, 29, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Brenè, S.; Mathé, A.A. BDNF in Schizophrenia, Depression and Corresponding Animal Models. Mol. Psychiatry 2005, 10, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T. Effects of Antipsychotics on the BDNF in Schizophrenia. Curr. Med. Chem. 2013, 20, 345–350. [Google Scholar] [PubMed]
- Zhang, X.Y.; Liang, J.; Chen, D.C.; Xiu, M.H.; De Yang, F.; Kosten, T.A.; Kosten, T.R. Low BDNF Is Associated with Cognitive Impairment in Chronic Patients with Schizophrenia. Psychopharmacology 2012, 222, 277–284. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Chen, D.C.; Xiu, M.H.; Haile, C.N.; Luo, X.; Xu, K.; Zhang, H.P.; Zuo, L.; Zhang, Z.; Zhang, X.; et al. Cognitive and Serum BDNF Correlates of BDNF Val66Met Gene Polymorphism in Patients with Schizophrenia and Normal Controls. Hum. Genet. 2012, 131, 1187–1195. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The Role of BDNF on Neural Plasticity in Depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Nestler, E.J. The Molecular Neurobiology of Depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, J.D. Traumatic Stress: Effects on the Brain. Dialogues Clin. Neurosci. 2006, 8, 445–461. [Google Scholar] [CrossRef]
- Levy, M.J.F.; Boulle, F.; Steinbusch, H.W.; van den Hove, D.L.A.; Kenis, G.; Lanfumey, L. Neurotrophic Factors and Neuroplasticity Pathways in the Pathophysiology and Treatment of Depression. Psychopharmacology 2018, 235, 2195–2220. [Google Scholar] [CrossRef] [Green Version]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the Role of BDNF as a Biomarker for Treatment of Depression. J. Mol. Neurosci. 2021, 71, 2008–2021. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Carnahan, J.; Greenberg, M.E. Requirement for BDNF in Activity-Dependent Survival of Cortical Neurons. Science 1994, 263, 1618–1623. [Google Scholar] [CrossRef]
- Ernfors, P.; Duan, M.L.; ElShamy, W.M.; Canlon, B. Protection of Auditory Neurons from Aminoglycoside Toxicity by Neurotrophin-3. Nat. Med. 1996, 2, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Agerman, K.; Hjerling-Leffler, J.; Blanchard, M.P.; Scarfone, E.; Canlon, B.; Nosrat, C.; Ernfors, P. BDNF Gene Replacement Reveals Multiple Mechanisms for Establishing Neurotrophin Specificity during Sensory Nervous System Development. Dev. Camb. Engl. 2003, 130, 1479–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, L.N.; Clark, G.M.; Marzella, P.L. Delayed Neurotrophin Treatment Supports Auditory Neuron Survival in Deaf Guinea Pigs. Neuroreport 2004, 15, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, L.M.; Conover, J.C.; Fritzsch, B.; DeChiara, T.; Lindsay, R.M.; Yancopoulos, G.D. Degeneration of Vestibular Neurons in Late Embryogenesis of Both Heterozygous and Homozygous BDNF Null Mutant Mice. Dev. Camb. Engl. 1996, 122, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Ernfors, P.; Lee, K.F.; Jaenisch, R. Mice Lacking Brain-Derived Neurotrophic Factor Develop with Sensory Deficits. Nature 1994, 368, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Fariñas, I.; Jones, K.R.; Tessarollo, L.; Vigers, A.J.; Huang, E.; Kirstein, M.; de Caprona, D.C.; Coppola, V.; Backus, C.; Reichardt, L.F.; et al. Spatial Shaping of Cochlear Innervation by Temporally Regulated Neurotrophin Expression. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 6170–6180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirvola, U.; Ylikoski, J. Neurotrophic Factors during Inner Ear Development. Curr. Top. Dev. Biol. 2003, 57, 207–223. [Google Scholar] [CrossRef]
- Schimmang, T.; Tan, J.; Müller, M.; Zimmermann, U.; Rohbock, K.; Kôpschall, I.; Limberger, A.; Minichiello, L.; Knipper, M. Lack of Bdnf and TrkB Signalling in the Postnatal Cochlea Leads to a Spatial Reshaping of Innervation along the Tonotopic Axis and Hearing Loss. Dev. Camb. Engl. 2003, 130, 4741–4750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ylikoski, J.; Pirvola, U.; Moshnyakov, M.; Palgi, J.; Arumäe, U.; Saarma, M. Expression Patterns of Neurotrophin and Their Receptor MRNAs in the Rat Inner Ear. Hear. Res. 1993, 65, 69–78. [Google Scholar] [CrossRef]
- Gestwa, G.; Wiechers, B.; Zimmermann, U.; Praetorius, M.; Rohbock, K.; Köpschall, I.; Zenner, H.P.; Knipper, M. Differential Expression of TrkB.T1 and TrkB.T2, Truncated TrkC, and P75(NGFR) in the Cochlea Prior to Hearing Function. J. Comp. Neurol. 1999, 414, 33–49. [Google Scholar] [CrossRef]
- Schimmang, T.; Minichiello, L.; Vazquez, E.; San Jose, I.; Giraldez, F.; Klein, R.; Represa, J. Developing Inner Ear Sensory Neurons Require TrkB and TrkC Receptors for Innervation of Their Peripheral Targets. Dev. Camb. Engl. 1995, 121, 3381–3391. [Google Scholar] [CrossRef]
- Green, S.H.; Bailey, E.; Wang, Q.; Davis, R.L. The Trk A, B, C’s of Neurotrophins in the Cochlea. Anat. Rec. Hoboken NJ 2007 2012, 295, 1877–1895. [Google Scholar] [CrossRef]
- Wan, G.; Gómez-Casati, M.E.; Gigliello, A.R.; Liberman, M.C.; Corfas, G. Neurotrophin-3 Regulates Ribbon Synapse Density in the Cochlea and Induces Synapse Regeneration after Acoustic Trauma. eLife 2014, 3, e03564. [Google Scholar] [CrossRef]
- Barbacid, M. The Trk Family of Neurotrophin Receptors. J. Neurobiol. 1994, 25, 1386–1403. [Google Scholar] [CrossRef] [PubMed]
- Snider, W.D. Functions of the Neurotrophins during Nervous System Development: What the Knockouts Are Teaching Us. Cell 1994, 77, 627–638. [Google Scholar] [CrossRef]
- Fritzsch, B.; Silos-Santiago, I.; Bianchi, L.M.; Fariñas, I. The Role of Neurotrophic Factors in Regulating the Development of Inner Ear Innervation. Trends Neurosci. 1997, 20, 159–164. [Google Scholar] [CrossRef]
- Ernfors, P.; Van De Water, T.; Loring, J.; Jaenisch, R. Complementary Roles of BDNF and NT-3 in Vestibular and Auditory Development. Neuron 1995, 14, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Minichiello, L.; Piehl, F.; Vazquez, E.; Schimmang, T.; Hökfelt, T.; Represa, J.; Klein, R. Differential Effects of Combined Trk Receptor Mutations on Dorsal Root Ganglion and Inner Ear Sensory Neurons. Dev. Camb. Engl. 1995, 121, 4067–4075. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.L. Gradients of Neurotrophins, Ion Channels, and Tuning in the Cochlea. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2003, 9, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramekers, D.; Versnel, H.; Grolman, W.; Klis, S.F.L. Neurotrophins and Their Role in the Cochlea. Hear. Res. 2012, 288, 19–33. [Google Scholar] [CrossRef]
- Adamson, C.L.; Reid, M.A.; Davis, R.L. Opposite Actions of Brain-Derived Neurotrophic Factor and Neurotrophin-3 on Firing Features and Ion Channel Composition of Murine Spiral Ganglion Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 1385–1396. [Google Scholar] [CrossRef] [Green Version]
- Hafidi, A. Distribution of BDNF, NT-3 and NT-4 in the Developing Auditory Brainstem. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1999, 17, 285–294. [Google Scholar] [CrossRef]
- Sobkowicz, H.M.; August, B.K.; Slapnick, S.M. Influence of Neurotrophins on the Synaptogenesis of Inner Hair Cells in the Deaf Bronx Waltzer (Bv) Mouse Organ of Corti in Culture. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2002, 20, 537–554. [Google Scholar] [CrossRef]
- Wissel, K.; Wefstaedt, P.; Miller, J.M.; Lenarz, T.; Stöver, T. Differential Brain-Derived Neurotrophic Factor and Transforming Growth Factor-Beta Expression in the Rat Cochlea Following Deafness. Neuroreport 2006, 17, 1297–1301. [Google Scholar] [CrossRef]
- Liu, W.; Kinnefors, A.; Boström, M.; Rask-Andersen, H. Expression of TrkB and BDNF in Human Cochlea-an Immunohistochemical Study. Cell Tissue Res. 2011, 345, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Wiechers, B.; Gestwa, G.; Mack, A.; Carroll, P.; Zenner, H.P.; Knipper, M. A Changing Pattern of Brain-Derived Neurotrophic Factor Expression Correlates with the Rearrangement of Fibers during Cochlear Development of Rats and Mice. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 3033–3042. [Google Scholar] [CrossRef] [Green Version]
- Bailey, E.M.; Green, S.H. Postnatal Expression of Neurotrophic Factors Accessible to Spiral Ganglion Neurons in the Auditory System of Adult Hearing and Deafened Rats. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 13110–13126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Otero, J.; Davis, R.L. Synaptic Proteins Are Tonotopically Graded in Postnatal and Adult Type I and Type II Spiral Ganglion Neurons. J. Comp. Neurol. 2011, 519, 1455–1475. [Google Scholar] [CrossRef] [Green Version]
- Meltser, I.; Tahera, Y.; Canlon, B. Differential Activation of Mitogen-Activated Protein Kinases and Brain-Derived Neurotrophic Factor after Temporary or Permanent Damage to a Sensory System. Neuroscience 2010, 165, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Rüttiger, L.; Panford-Walsh, R.; Schimmang, T.; Tan, J.; Zimmermann, U.; Rohbock, K.; Köpschall, I.; Limberger, A.; Müller, M.; Fraenzer, J.-T.; et al. BDNF MRNA Expression and Protein Localization Are Changed in Age-Related Hearing Loss. Neurobiol. Aging 2007, 28, 586–601. [Google Scholar] [CrossRef] [PubMed]
- Marchetta, P.; Savitska, D.; Kübler, A.; Asola, G.; Manthey, M.; Möhrle, D.; Schimmang, T.; Rüttiger, L.; Knipper, M.; Singer, W. Age-Dependent Auditory Processing Deficits after Cochlear Synaptopathy Depend on Auditory Nerve Latency and the Ability of the Brain to Recruit LTP/BDNF. Brain Sci. 2020, 10, 710. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Picciotti, P.M.; Paludetti, G.; Troiani, D. Pathogenesis of Presbycusis in Animal Models: A Review. Exp. Gerontol. 2011, 46, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Puel, J.-L. Presbycusis: An Update on Cochlear Mechanisms and Therapies. J. Clin. Med. 2020, 9, E218. [Google Scholar] [CrossRef] [Green Version]
- Paciello, F.; Pisani, A.; Rinaudo, M.; Cocco, S.; Paludetti, G.; Fetoni, A.R.; Grassi, C. Noise-Induced Auditory Damage Affects Hippocampus Causing Memory Deficits in a Model of Early Age-Related Hearing Loss. Neurobiol. Dis. 2023, 178, 106024. [Google Scholar] [CrossRef]
- Rask-Andersen, H.; Boström, M.; Gerdin, B.; Kinnefors, A.; Nyberg, G.; Engstrand, T.; Miller, J.M.; Lindholm, D. Regeneration of Human Auditory Nerve. In Vitro/in Video Demonstration of Neural Progenitor Cells in Adult Human and Guinea Pig Spiral Ganglion. Hear. Res. 2005, 203, 180–191. [Google Scholar] [CrossRef]
- Wilson, B.S.; Tucci, D.L.; Merson, M.H.; O’Donoghue, G.M. Global Hearing Health Care: New Findings and Perspectives. Lancet Lond. Engl. 2017, 390, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Deafness and Hearing Loss. Available online: https://www.who.int/news-room/fact-sheets/detail/deafness-and-hearing-loss (accessed on 8 February 2023).
- Fetoni, A.R.; Pisani, A.; Rolesi, R.; Paciello, F.; Viziano, A.; Moleti, A.; Sisto, R.; Troiani, D.; Paludetti, G.; Grassi, C. Early Noise-Induced Hearing Loss Accelerates Presbycusis Altering Aging Processes in the Cochlea. Front. Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Gomaa, N.A.; Jimoh, Z.; Campbell, S.; Zenke, J.K.; Szczepek, A.J. Biomarkers for Inner Ear Disorders: Scoping Review on the Role of Biomarkers in Hearing and Balance Disorders. Diagn. Basel Switz. 2020, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Paludetti, G.; Troiani, D. Targeting Dysregulation of Redox Homeostasis in Noise-Induced Hearing Loss: Oxidative Stress and ROS Signaling. Free Radic. Biol. Med. 2019, 135, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The Role of Oxidative Stress in Noise-Induced Hearing Loss. Ear Hear. 2006, 27, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauschecker, J.P. Auditory Cortical Plasticity: A Comparison with Other Sensory Systems. Trends Neurosci. 1999, 22, 74–80. [Google Scholar] [CrossRef]
- Syka, J. Plastic Changes in the Central Auditory System after Hearing Loss, Restoration of Function, and during Learning. Physiol. Rev. 2002, 82, 601–636. [Google Scholar] [CrossRef]
- Caspary, D.M.; Ling, L.; Turner, J.G.; Hughes, L.F. Inhibitory Neurotransmission, Plasticity and Aging in the Mammalian Central Auditory System. J. Exp. Biol. 2008, 211, 1781–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Weiner, B.D.; Zhang, L.S.; Cho, S.-J.; Bao, S. Homeostatic Plasticity Drives Tinnitus Perception in an Animal Model. Proc. Natl. Acad. Sci. USA 2011, 108, 14974–14979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberman, M.C.; Kiang, N.Y. Acoustic Trauma in Cats. Cochlear Pathology and Auditory-Nerve Activity. Acta Oto-Laryngol. Suppl. 1978, 358, 1–63. [Google Scholar]
- Milbrandt, J.C.; Holder, T.M.; Wilson, M.C.; Salvi, R.J.; Caspary, D.M. GAD Levels and Muscimol Binding in Rat Inferior Colliculus Following Acoustic Trauma. Hear. Res. 2000, 147, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Salvi, R.J.; Wang, J.; Ding, D. Auditory Plasticity and Hyperactivity Following Cochlear Damage. Hear. Res. 2000, 147, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.D.; Brozoski, T.J.; Ling, L.L.; Caspary, D.M. Targeting Inhibitory Neurotransmission in Tinnitus. Brain Res. 2012, 1485, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engineer, N.D.; Møller, A.R.; Kilgard, M.P. Directing Neural Plasticity to Understand and Treat Tinnitus. Hear. Res. 2013, 295, 58–66. [Google Scholar] [CrossRef]
- Fetoni, A.R.; De Bartolo, P.; Eramo, S.L.M.; Rolesi, R.; Paciello, F.; Bergamini, C.; Fato, R.; Paludetti, G.; Petrosini, L.; Troiani, D. Noise-Induced Hearing Loss (NIHL) as a Target of Oxidative Stress-Mediated Damage: Cochlear and Cortical Responses after an Increase in Antioxidant Defense. J. Neurosci. 2013, 33, 4011–4023. [Google Scholar] [CrossRef] [Green Version]
- Fetoni, A.R.; Troiani, D.; Petrosini, L.; Paludetti, G. Cochlear Injury and Adaptive Plasticity of the Auditory Cortex. Front. Aging Neurosci. 2015, 7, 8. [Google Scholar] [CrossRef]
- Paciello, F.; Podda, M.V.; Rolesi, R.; Cocco, S.; Petrosini, L.; Troiani, D.; Fetoni, A.R.; Paludetti, G.; Grassi, C. Anodal Transcranial Direct Current Stimulation Affects Auditory Cortex Plasticity in Normal-Hearing and Noise-Exposed Rats. Brain Stimulat. 2018, 11, 1008–1023. [Google Scholar] [CrossRef]
- Chengzhi, C.; Yan, T.; Xuejun, J.; Xiang, L.; Youbin, Q.; Baijie, T. Recovery of Chronic Noise Exposure Induced Spatial Learning and Memory Deficits in Young Male Sprague-Dawley Rats. J. Occup. Health 2011, 53, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.; Li, K. Chronic Noise Exposure and Alzheimer Disease: Is There an Etiological Association? Med. Hypotheses 2013, 81, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, D.; Feldmann, M.; Shchyglo, O.; Manahan-Vaughan, D. Hippocampal Synaptic Plasticity, Spatial Memory, and Neurotransmitter Receptor Expression Are Profoundly Altered by Gradual Loss of Hearing Ability. Cereb. Cortex N. Y. N 1991 2020, 30, 4581–4596. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Liu, L.; Shi, L.; Li, X.; Shen, P.; Xun, Q.; Guo, X.; Yu, Z.; Wang, J. Spatial Learning and Memory Deficits in Young Adult Mice Exposed to a Brief Intense Noise at Postnatal Age. J. Otol. 2015, 10, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurioka, T.; Mogi, S.; Yamashita, T. Decreasing Auditory Input Induces Neurogenesis Impairment in the Hippocampus. Sci. Rep. 2021, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Juhn, S.K.; Rybak, L.P. Labyrinthine Barriers and Cochlear Homeostasis. Acta Otolaryngol. (Stockh.) 1981, 91, 529–534. [Google Scholar] [CrossRef]
- Sun, W.; Wang, W. Advances in Research on Labyrinth Membranous Barriers. J. Otol. 2015, 10, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetoni, A.R.; Garzaro, M.; Ralli, M.; Landolfo, V.; Sensini, M.; Pecorari, G.; Mordente, A.; Paludetti, G.; Giordano, C. The Monitoring Role of Otoacoustic Emissions and Oxidative Stress Markers in the Protective Effects of Antioxidant Administration in Noise-Exposed Subjects: A Pilot Study. Med. Sci. Monit. 2009, 15, PR1–PR8. [Google Scholar]
- Zine, A.; Messat, Y.; Fritzsch, B. A Human Induced Pluripotent Stem Cell-Based Modular Platform to Challenge Sensorineural Hearing Loss. Stem Cells Dayt. Ohio 2021, 39, 697–706. [Google Scholar] [CrossRef]
- Liberman, M.C.; Kujawa, S.G. Cochlear Synaptopathy in Acquired Sensorineural Hearing Loss: Manifestations and Mechanisms. Hear. Res. 2017, 349, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Staecker, H.; Gabaizadeh, R.; Federoff, H.; Van De Water, T.R. Brain-Derived Neurotrophic Factor Gene Therapy Prevents Spiral Ganglion Degeneration after Hair Cell Loss. Otolaryngol.--Head Neck Surg. Off. J. Am. Acad. Otolaryngol.-Head Neck Surg. 1998, 119, 7–13. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, S.L.; Shepherd, R.K. Exogenous BDNF Rescues Rat Spiral Ganglion Neurons in Vivo. Otol. Neurotol. Off. Publ. Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2005, 26, 1064–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noushi, F.; Richardson, R.T.; Hardman, J.; Clark, G.; O’Leary, S. Delivery of Neurotrophin-3 to the Cochlea Using Alginate Beads. Otol. Neurotol. Off. Publ. Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2005, 26, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.T.; O’Leary, S.; Wise, A.; Hardman, J.; Clark, G. A Single Dose of Neurotrophin-3 to the Cochlea Surrounds Spiral Ganglion Neurons and Provides Trophic Support. Hear. Res. 2005, 204, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Le Prell, C.G.; Prieskorn, D.M.; Wys, N.L.; Altschuler, R.A. Delayed Neurotrophin Treatment Following Deafness Rescues Spiral Ganglion Cells from Death and Promotes Regrowth of Auditory Nerve Peripheral Processes: Effects of Brain-Derived Neurotrophic Factor and Fibroblast Growth Factor. J. Neurosci. Res. 2007, 85, 1959–1969. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, R.K.; Coco, A.; Epp, S.B.; Crook, J.M. Chronic Depolarization Enhances the Trophic Effects of Brain-Derived Neurotrophic Factor in Rescuing Auditory Neurons Following a Sensorineural Hearing Loss. J. Comp. Neurol. 2005, 486, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, R.K.; Coco, A.; Epp, S.B. Neurotrophins and Electrical Stimulation for Protection and Repair of Spiral Ganglion Neurons Following Sensorineural Hearing Loss. Hear. Res. 2008, 242, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, A.K.; Richardson, R.; Hardman, J.; Clark, G.; O’leary, S. Resprouting and Survival of Guinea Pig Cochlear Neurons in Response to the Administration of the Neurotrophins Brain-Derived Neurotrophic Factor and Neurotrophin-3. J. Comp. Neurol. 2005, 487, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Rejali, D.; Lee, V.A.; Abrashkin, K.A.; Humayun, N.; Swiderski, D.L.; Raphael, Y. Cochlear Implants and Ex Vivo BDNF Gene Therapy Protect Spiral Ganglion Neurons. Hear. Res. 2007, 228, 180–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agterberg, M.J.H.; Versnel, H.; de Groot, J.C.M.J.; Smoorenburg, G.F.; Albers, F.W.J.; Klis, S.F.L. Morphological Changes in Spiral Ganglion Cells after Intracochlear Application of Brain-Derived Neurotrophic Factor in Deafened Guinea Pigs. Hear. Res. 2008, 244, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Agterberg, M.J.H.; Versnel, H.; van Dijk, L.M.; de Groot, J.C.M.J.; Klis, S.F.L. Enhanced Survival of Spiral Ganglion Cells after Cessation of Treatment with Brain-Derived Neurotrophic Factor in Deafened Guinea Pigs. J. Assoc. Res. Otolaryngol. JARO 2009, 10, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Glueckert, R.; Bitsche, M.; Miller, J.M.; Zhu, Y.; Prieskorn, D.M.; Altschuler, R.A.; Schrott-Fischer, A. Deafferentation-Associated Changes in Afferent and Efferent Processes in the Guinea Pig Cochlea and Afferent Regeneration with Chronic Intrascalar Brain-Derived Neurotrophic Factor and Acidic Fibroblast Growth Factor. J. Comp. Neurol. 2008, 507, 1602–1621. [Google Scholar] [CrossRef] [Green Version]
- Wise, A.K.; Hume, C.R.; Flynn, B.O.; Jeelall, Y.S.; Suhr, C.L.; Sgro, B.E.; O’Leary, S.J.; Shepherd, R.K.; Richardson, R.T. Effects of Localized Neurotrophin Gene Expression on Spiral Ganglion Neuron Resprouting in the Deafened Cochlea. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Havenith, S.; Versnel, H.; Agterberg, M.J.H.; de Groot, J.C.M.J.; Sedee, R.-J.; Grolman, W.; Klis, S.F.L. Spiral Ganglion Cell Survival after Round Window Membrane Application of Brain-Derived Neurotrophic Factor Using Gelfoam as Carrier. Hear. Res. 2011, 272, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Leake, P.A.; Hradek, G.T.; Hetherington, A.M.; Stakhovskaya, O. Brain-Derived Neurotrophic Factor Promotes Cochlear Spiral Ganglion Cell Survival and Function in Deafened, Developing Cats. J. Comp. Neurol. 2011, 519, 1526–1545. [Google Scholar] [CrossRef] [Green Version]
- Pettingill, L.N.; Wise, A.K.; Geaney, M.S.; Shepherd, R.K. Enhanced Auditory Neuron Survival Following Cell-Based BDNF Treatment in the Deaf Guinea Pig. PLoS ONE 2011, 6, e18733. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Lyu, A.-R.; Park, S.-J.; Xu, J.; Cui, J.; Sohn, K.-C.; Hur, G.M.; Jin, Y.; Park, Y.-H. Early Postnatal NT-3 Gene Delivery Enhances Hearing Acquisition in the Developmental Period. Laryngoscope 2016, 126, E379–E385. [Google Scholar] [CrossRef] [PubMed]
- Dabdoub, A.; Nishimura, K. Cochlear Implants Meet Regenerative Biology: State of the Science and Future Research Directions. Otol. Neurotol. Off. Publ. Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2017, 38, e232–e236. [Google Scholar] [CrossRef]
- Konerding, W.S.; Janssen, H.; Hubka, P.; Tornøe, J.; Mistrik, P.; Wahlberg, L.; Lenarz, T.; Kral, A.; Scheper, V. Encapsulated Cell Device Approach for Combined Electrical Stimulation and Neurotrophic Treatment of the Deaf Cochlea. Hear. Res. 2017, 350, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Song, B.-N.; Li, Y.-X.; Han, D.-M. Delayed Electrical Stimulation and BDNF Application Following Induced Deafness in Rats. Acta Otolaryngol. (Stockh.) 2009, 129, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, A.; Wissel, K.; Hoffmann, A.; Hofmann, N.; Berkingali, N.; Gro, G.; Lenarz, T.; Stöver, T. The Biological Effects of Cell-Delivered Brain-Derived Neurotrophic Factor on Cultured Spiral Ganglion Cells. Neuroreport 2007, 18, 1683–1686. [Google Scholar] [CrossRef] [PubMed]
- Coco, A.; Epp, S.B.; Fallon, J.B.; Xu, J.; Millard, R.E.; Shepherd, R.K. Does Cochlear Implantation and Electrical Stimulation Affect Residual Hair Cells and Spiral Ganglion Neurons? Hear. Res. 2007, 225, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Scheper, V.; Paasche, G.; Miller, J.M.; Warnecke, A.; Berkingali, N.; Lenarz, T.; Stöver, T. Effects of Delayed Treatment with Combined GDNF and Continuous Electrical Stimulation on Spiral Ganglion Cell Survival in Deafened Guinea Pigs. J. Neurosci. Res. 2009, 87, 1389–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wefstaedt, P.; Scheper, V.; Lenarz, T.; Stöver, T. Brain-Derived Neurotrophic Factor/Glial Cell Line-Derived Neurotrophic Factor Survival Effects on Auditory Neurons Are Not Limited by Dexamethasone. Neuroreport 2005, 16, 2011–2014. [Google Scholar] [CrossRef]
- Hegarty, J.L.; Kay, A.R.; Green, S.H. Trophic Support of Cultured Spiral Ganglion Neurons by Depolarization Exceeds and Is Additive with That by Neurotrophins or CAMP and Requires Elevation of [Ca2+]i within a Set Range. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 1959–1970. [Google Scholar] [CrossRef] [Green Version]
- Schwieger, J.; Warnecke, A.; Lenarz, T.; Esser, K.-H.; Scheper, V. Neuronal Survival, Morphology and Outgrowth of Spiral Ganglion Neurons Using a Defined Growth Factor Combination. PLoS ONE 2015, 10, e0133680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.L.; Stewart, R.R.; Gao, W.Q. Neurotrophin-4/5 Enhances Survival of Cultured Spiral Ganglion Neurons and Protects Them from Cisplatin Neurotoxicity. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 5079–5087. [Google Scholar] [CrossRef]
- Zheng, J.L.; Gao, W.Q. Differential Damage to Auditory Neurons and Hair Cells by Ototoxins and Neuroprotection by Specific Neurotrophins in Rat Cochlear Organotypic Cultures. Eur. J. Neurosci. 1996, 8, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.L.; Ulfendahl, M.; Laurell, G.; Counter, S.A.; Pyykkö, I.; Borg, E.; Rosenhall, U. Protection and Treatment of Sensorineural Hearing Disorders Caused by Exogenous Factors: Experimental Findings and Potential Clinical Application. Hear. Res. 2002, 169, 169–178. [Google Scholar] [CrossRef]
- Zhai, S.-Q.; Guo, W.; Hu, Y.-Y.; Yu, N.; Chen, Q.; Wang, J.-Z.; Fan, M.; Yang, W.-Y. Protective Effects of Brain-Derived Neurotrophic Factor on the Noise-Damaged Cochlear Spiral Ganglion. J. Laryngol. Otol. 2011, 125, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Bowe, S.N.; Jacob, A. Round Window Perfusion Dynamics: Implications for Intracochlear Therapy. Curr. Opin. Otolaryngol. Head Neck Surg. 2010, 18, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Staecker, H.; Garnham, C. Neurotrophin Therapy and Cochlear Implantation: Translating Animal Models to Human Therapy. Exp. Neurol. 2010, 226, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kuang, R.; Hever, G.; Zajic, G.; Yan, Q.; Collins, F.; Louis, J.C.; Keithley, E.; Magal, E. Glial Cell Line-Derived Neurotrophic Factor. Potential for Otoprotection. Ann. N. Y. Acad. Sci. 1999, 884, 270–291. [Google Scholar] [CrossRef]
- Endo, T.; Nakagawa, T.; Kita, T.; Iguchi, F.; Kim, T.-S.; Tamura, T.; Iwai, K.; Tabata, Y.; Ito, J. Novel Strategy for Treatment of Inner Ears Using a Biodegradable Gel. Laryngoscope 2005, 115, 2016–2020. [Google Scholar] [CrossRef] [PubMed]
- Rathnam, C.; Chueng, S.-T.D.; Ying, Y.-L.M.; Lee, K.-B.; Kwan, K. Developments in Bio-Inspired Nanomaterials for Therapeutic Delivery to Treat Hearing Loss. Front. Cell. Neurosci. 2019, 13, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Zheng, J.; He, Y.; Lin, K.; Li, S.; Zhang, Y.; Song, P.; Zhou, Y.; Chen, X. Nanocarriers for Inner Ear Disease Therapy. Front. Cell. Neurosci. 2021, 15, 791573. [Google Scholar] [CrossRef]
- Foster, A.C.; Szobota, S.; Piu, F.; Jacques, B.E.; Moore, D.R.; Sanchez, V.A.; Anderson, J.J. A Neurotrophic Approach to Treating Hearing Loss: Translation from Animal Models to Clinical Proof-of-Concept. J. Acoust. Soc. Am. 2022, 151, 3937. [Google Scholar] [CrossRef] [PubMed]
- Piu, F.; Tsivkovskaia, N.; Fernandez, R.; Wang, X.; Harrop-Jones, A.; Altman, T.; Hou, J.; Pastuszka, M.; Vasquez, C.; DeGuzman, R.; et al. A Sustained-Exposure Formulation of the Neurotrophic Factor BDNF Protects against Noise-Induced Cochlear Synaptopathy in Young Adult and Aged Rats. In Proceedings of the 41st Association for Research in Otolaryngology Mid-Winter Meeting, San Diego, CA, USA, 10–14 February 2018. [Google Scholar]
- Warnecke, A.; Sasse, S.; Wenzel, G.I.; Hoffmann, A.; Gross, G.; Paasche, G.; Scheper, V.; Reich, U.; Esser, K.-H.; Lenarz, T.; et al. Stable Release of BDNF from the Fibroblast Cell Line NIH3T3 Grown on Silicone Elastomers Enhances Survival of Spiral Ganglion Cells in Vitro and in Vivo. Hear. Res. 2012, 289, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, S.; Kawamoto, K.; Kanzaki, S.; Raphael, Y. Gene Transfer into Supporting Cells of the Organ of Corti. Hear. Res. 2002, 173, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, S.; Kawamoto, K.; Stöver, T.; Kanzaki, S.; Yamasoba, T.; Raphael, Y. A Glucocorticoid Reduces Adverse Effects of Adenovirus Vectors in the Cochlea. Audiol. Neurootol. 2003, 8, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Hariri, A.R.; Goldberg, T.E.; Mattay, V.S.; Kolachana, B.S.; Callicott, J.H.; Egan, M.F.; Weinberger, D.R. Brain-Derived Neurotrophic Factor Val66met Polymorphism Affects Human Memory-Related Hippocampal Activity and Predicts Memory Performance. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 6690–6694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibu, K.; Kondo, K.; Ohta, Y.; Ishibashi, T.; Rothstein, J.L.; Kaga, K. Expression of NeuroD and TrkB in Developing and Aged Mouse Olfactory Epithelium. NeuroReport 2001, 12, 1615. [Google Scholar] [CrossRef]
- Feron, F.; Bianco, J.; Ferguson, I.; Mackay-Sim, A. Neurotrophin Expression in the Adult Olfactory Epithelium. Brain Res. 2008, 1196, 13–21. [Google Scholar] [CrossRef]
- Kim, B.-Y.; Park, J.Y.; Cho, K.J.; Bae, J.H. The Potential Cytotoxic Effects of Urban Particle Matter on Olfaction. Rhinology 2021, 59, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kuvarzin, S.R.; Mor, M.S.; Katolikova, N.V.; Shemiakova, T.S.; Razenkova, V.; Ptukha, M.; Kozlova, A.A.; Murtazina, R.Z.; Smirnova, D.; et al. Trace Amine-Associated Receptor 2 Is Expressed in the Limbic Brain Areas and Is Involved in Dopamine Regulation and Adult Neurogenesis. Front. Behav. Neurosci. 2022, 16, 847410. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-T.; Hong, T.-Y.; Yang, C.-J.; Hsieh, J.-C. Variant Brain-Derived Neurotrophic Factor Val66met Polymorphism Engages Memory-Associated Systems to Augment Olfaction. Sci. Rep. 2022, 12, 20007. [Google Scholar] [CrossRef] [PubMed]
- Uranagase, A.; Katsunuma, S.; Doi, K.; Nibu, K.-I. BDNF Expression in Olfactory Bulb and Epithelium during Regeneration of Olfactory Epithelium. Neurosci. Lett. 2012, 516, 45–49. [Google Scholar] [CrossRef]
- Sjölund, S.; Larsson, M.; Olofsson, J.K.; Seubert, J.; Laukka, E.J. Phantom Smells: Prevalence and Correlates in a Population-Based Sample of Older Adults. Chem. Senses 2017, 42, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hedner, M.; Nilsson, L.-G.; Olofsson, J.K.; Bergman, O.; Eriksson, E.; Nyberg, L.; Larsson, M. Age-Related Olfactory Decline Is Associated with the BDNF Val66met Polymorphism: Evidence from a Population-Based Study. Front. Aging Neurosci. 2010, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Biju, K.C.; Mast, T.G.; Fadool, D.A. Olfactory Sensory Deprivation Increases the Number of ProBDNF-Immunoreactive Mitral Cells in the Olfactory Bulb of Mice. Neurosci. Lett. 2008, 447, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Xie, G.; Zhou, T.; Tu, J.; Zhang, J.; Lin, Z.; Zhang, H.; Gao, L. Intranasal Administration of White Tea Alleviates the Olfactory Function Deficit Induced by Chronic Unpredictable Mild Stress. Pharm. Biol. 2020, 58, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, J.; Kawaguchi, Y.; Kawakami, Y.; Takei, K. Brain-Derived Neurotrophic Factor (BDNF) Induces Antagonistic Action to Nogo Signaling by the Upregulation of Lateral Olfactory Tract Usher Substance (LOTUS) Expression. J. Neurochem. 2023, 164, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Chen, X.; Deng, J.; Wei, J.; Qian, H.; Huang, Y.; Wu, S.; Gao, F.; Gong, C.; Liao, Y.; et al. Dexmedetomidine Alleviates Olfactory Cognitive Dysfunction by Promoting Neurogenesis in the Subventricular Zone of Hypoxic-Ischemic Neonatal Rats. Front. Pharmacol. 2022, 13, 983920. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pisani, A.; Paciello, F.; Del Vecchio, V.; Malesci, R.; De Corso, E.; Cantone, E.; Fetoni, A.R. The Role of BDNF as a Biomarker in Cognitive and Sensory Neurodegeneration. J. Pers. Med. 2023, 13, 652. https://doi.org/10.3390/jpm13040652
Pisani A, Paciello F, Del Vecchio V, Malesci R, De Corso E, Cantone E, Fetoni AR. The Role of BDNF as a Biomarker in Cognitive and Sensory Neurodegeneration. Journal of Personalized Medicine. 2023; 13(4):652. https://doi.org/10.3390/jpm13040652
Chicago/Turabian StylePisani, Anna, Fabiola Paciello, Valeria Del Vecchio, Rita Malesci, Eugenio De Corso, Elena Cantone, and Anna Rita Fetoni. 2023. "The Role of BDNF as a Biomarker in Cognitive and Sensory Neurodegeneration" Journal of Personalized Medicine 13, no. 4: 652. https://doi.org/10.3390/jpm13040652