
Identifying Oxidized Lipid Metabolism-Related LncRNAs as Prognostic Biomarkers of Head and Neck Squamous Cell Carcinoma
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of Information of Patients with HNSCC
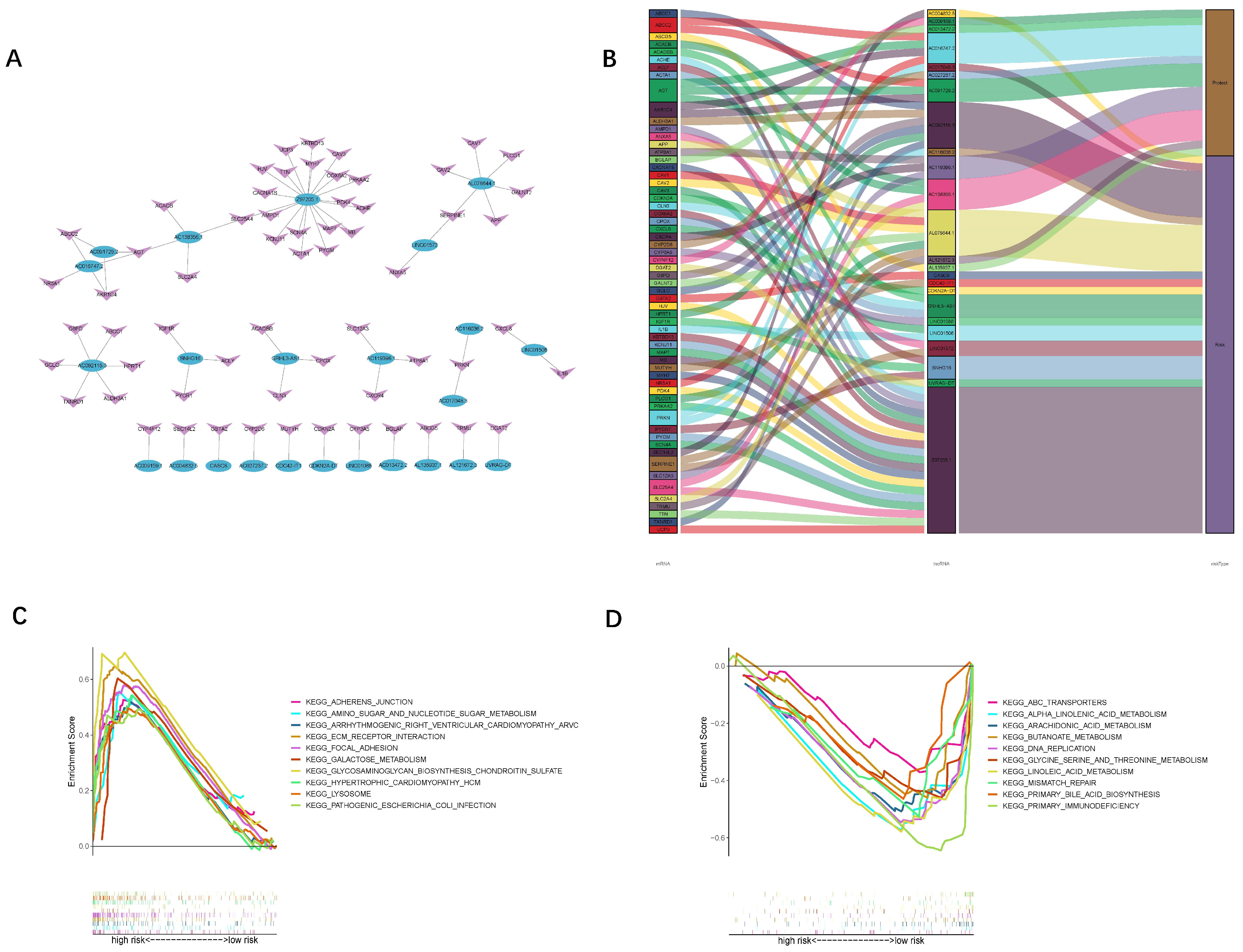
2.2. Selection of Oxidized Lipid Metabolism-Related LncRNAs
2.3. Establishment and Validation of the Risk Score
2.4. Validation of Prognostic Signature
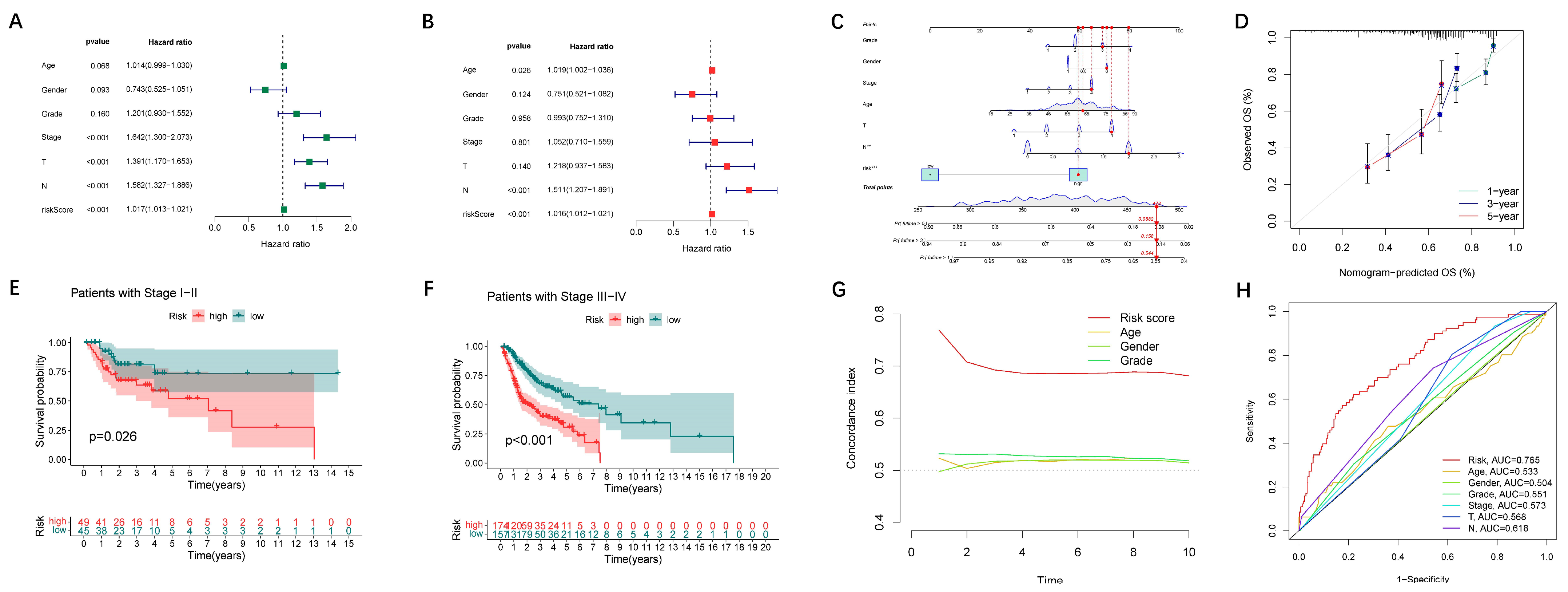
2.5. Construction and Application of Nomogram
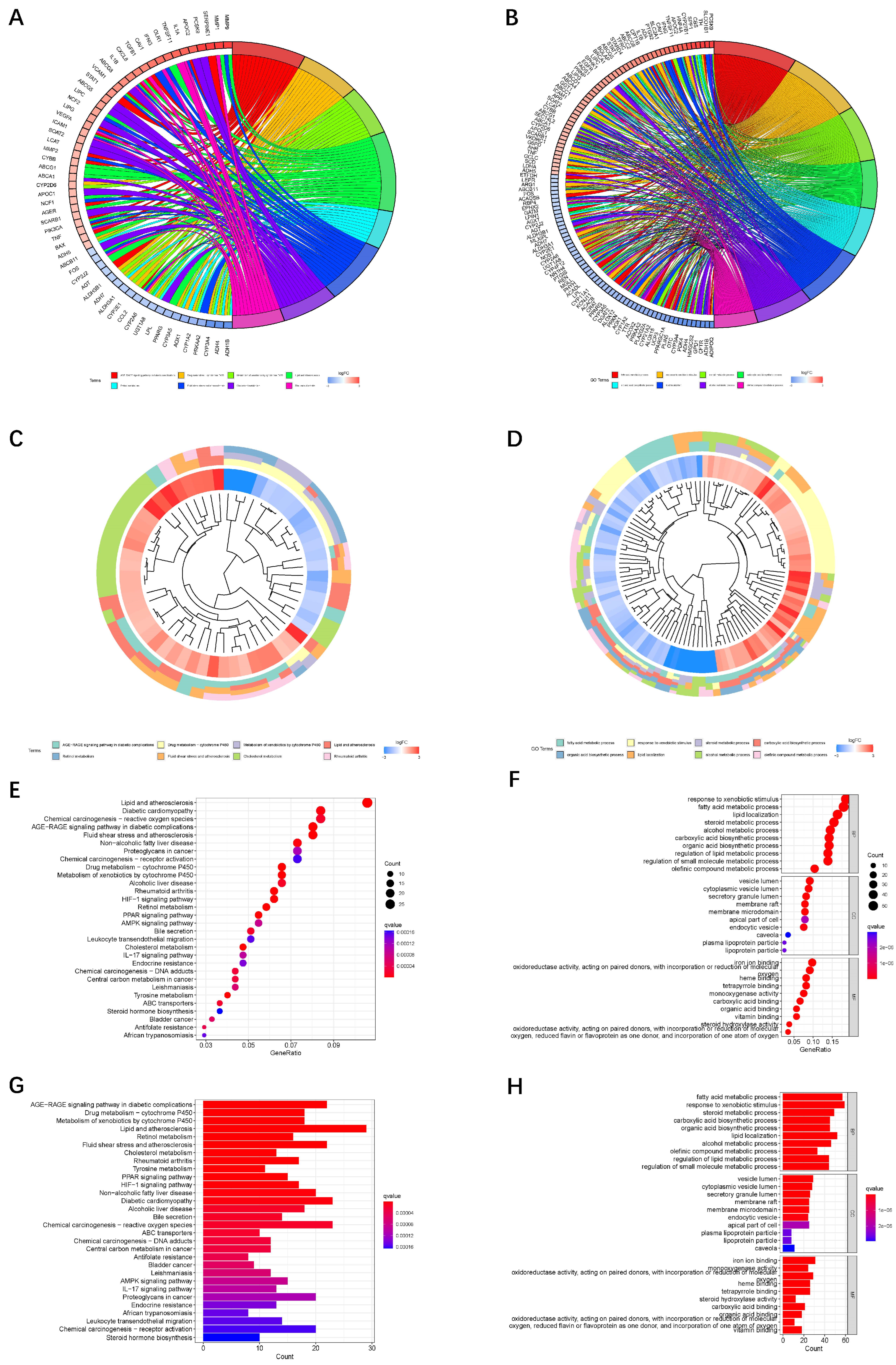
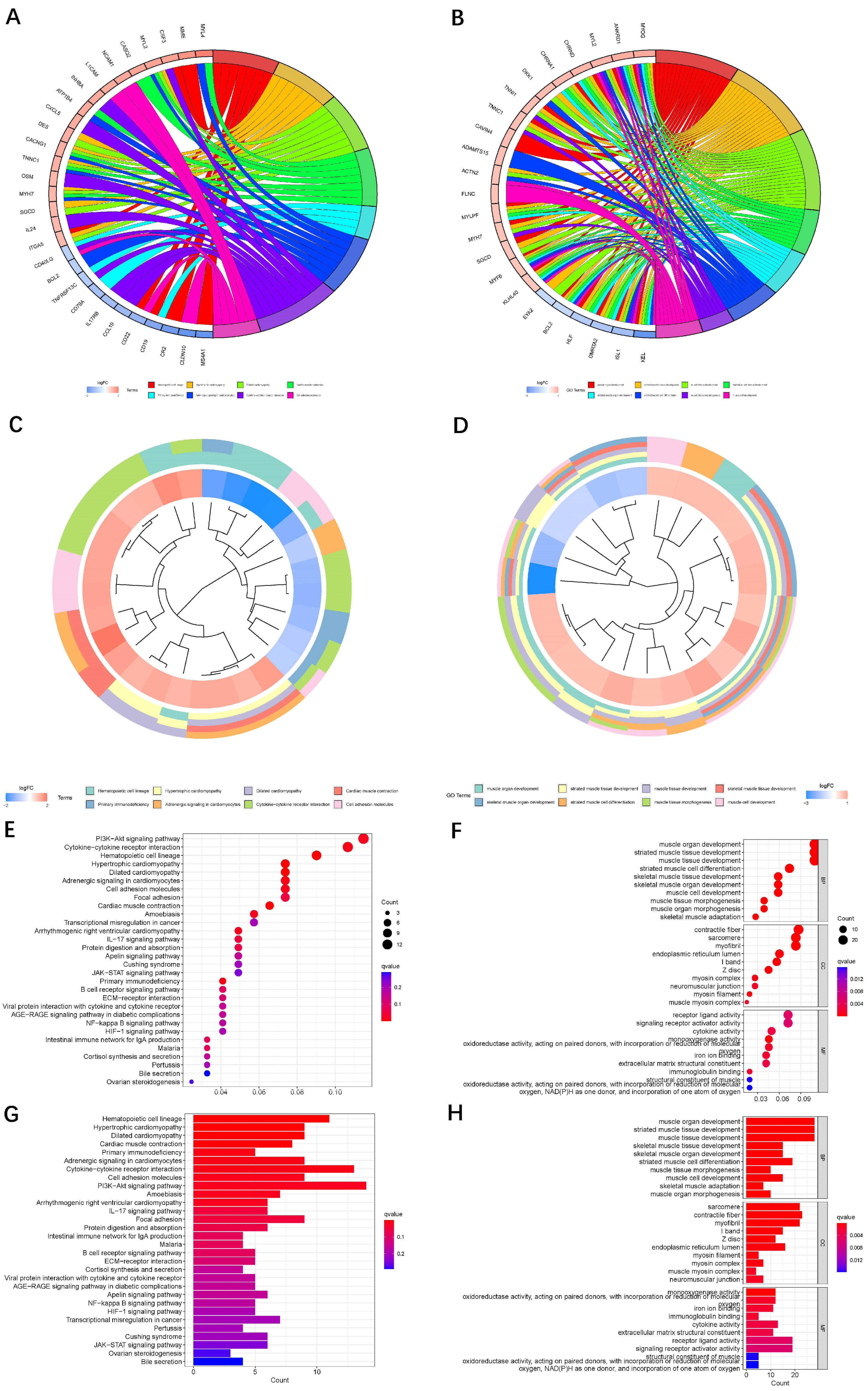
2.6. Functional Enrichment Analysis
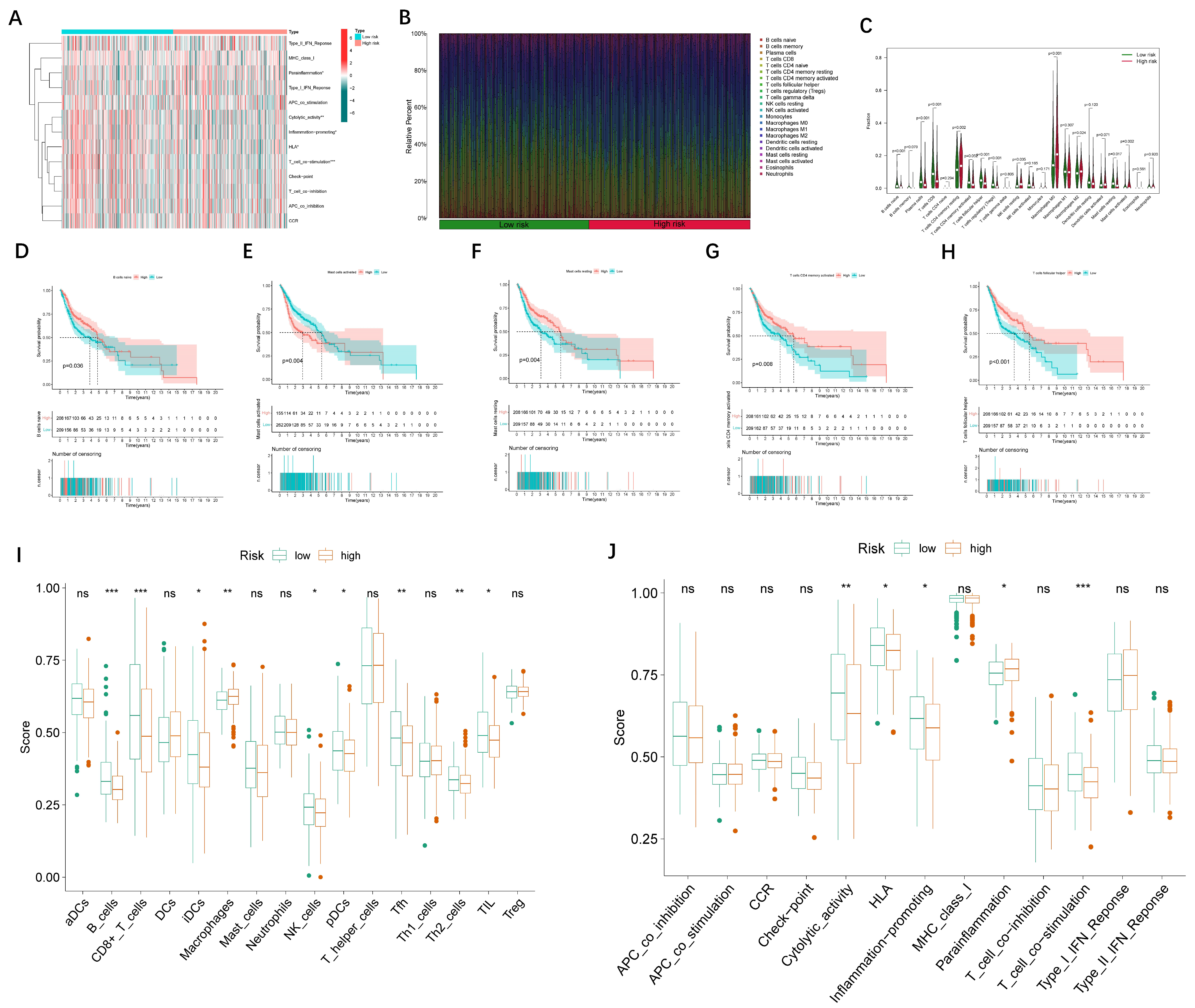
2.7. Analysis of Immune Cell Infiltration in High- and Low-Risk Patient Groups
2.8. Statistical Analysis
3. Results
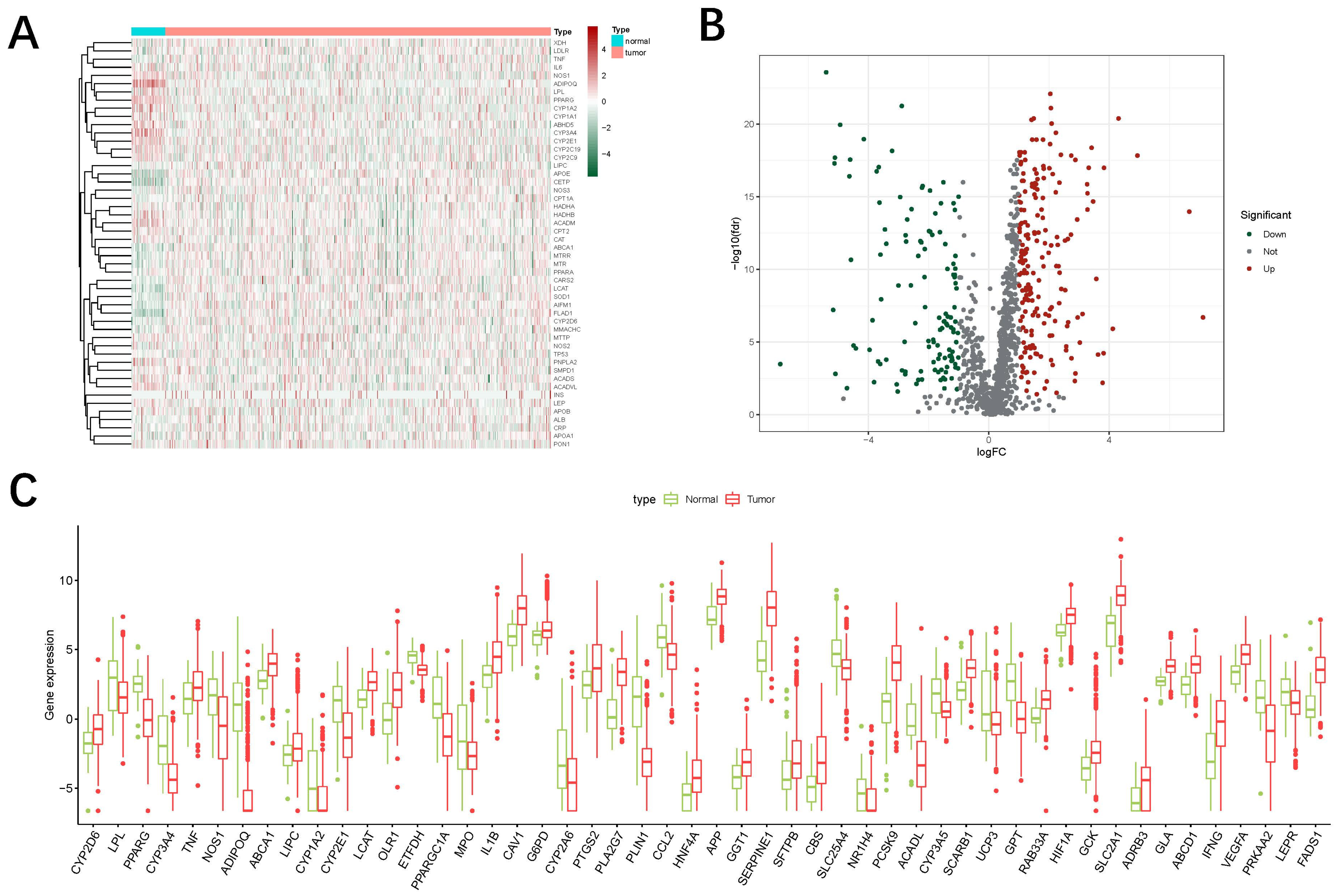
3.1. Identification of Oxidized Lipid Metabolism-Related LncRNAs
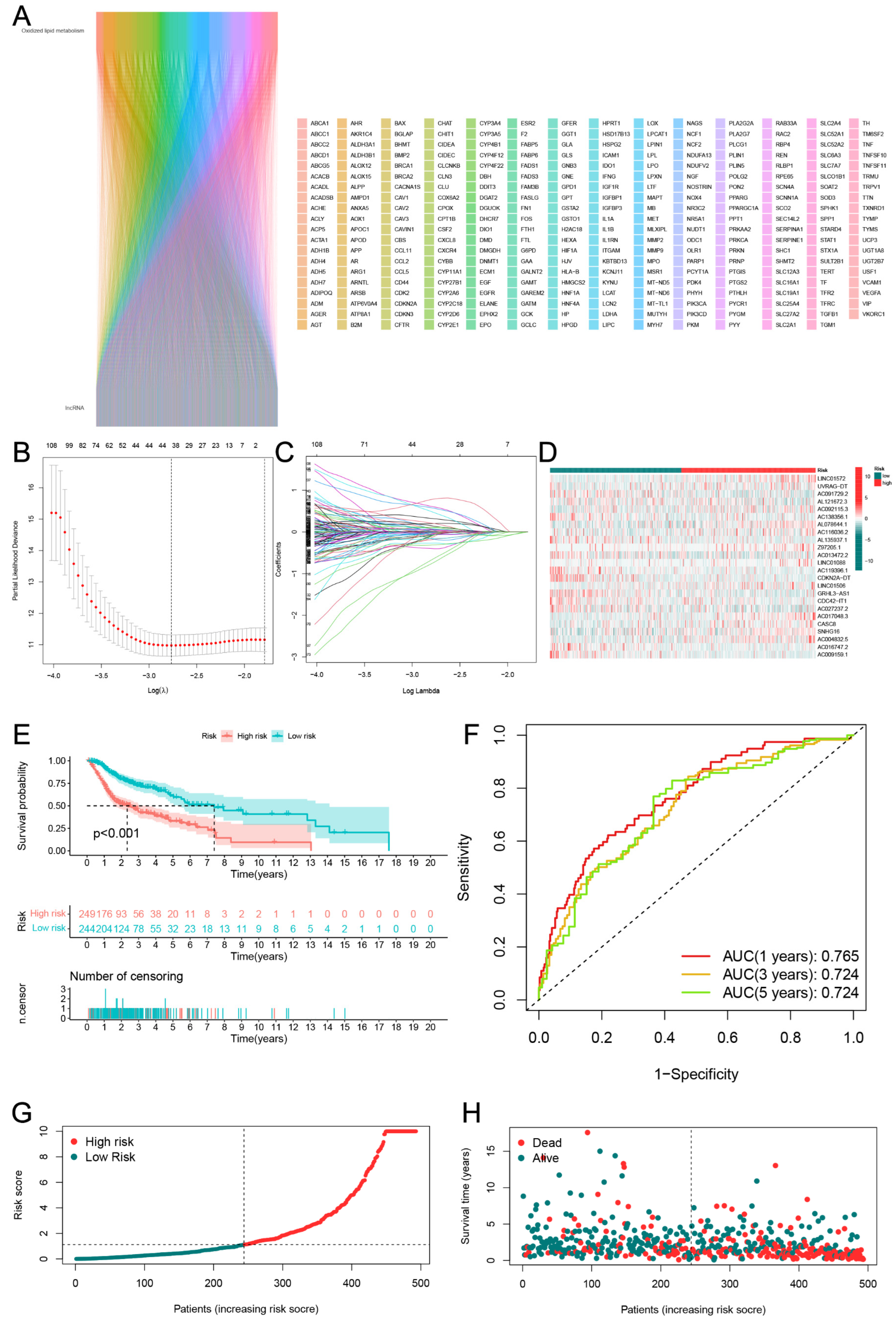
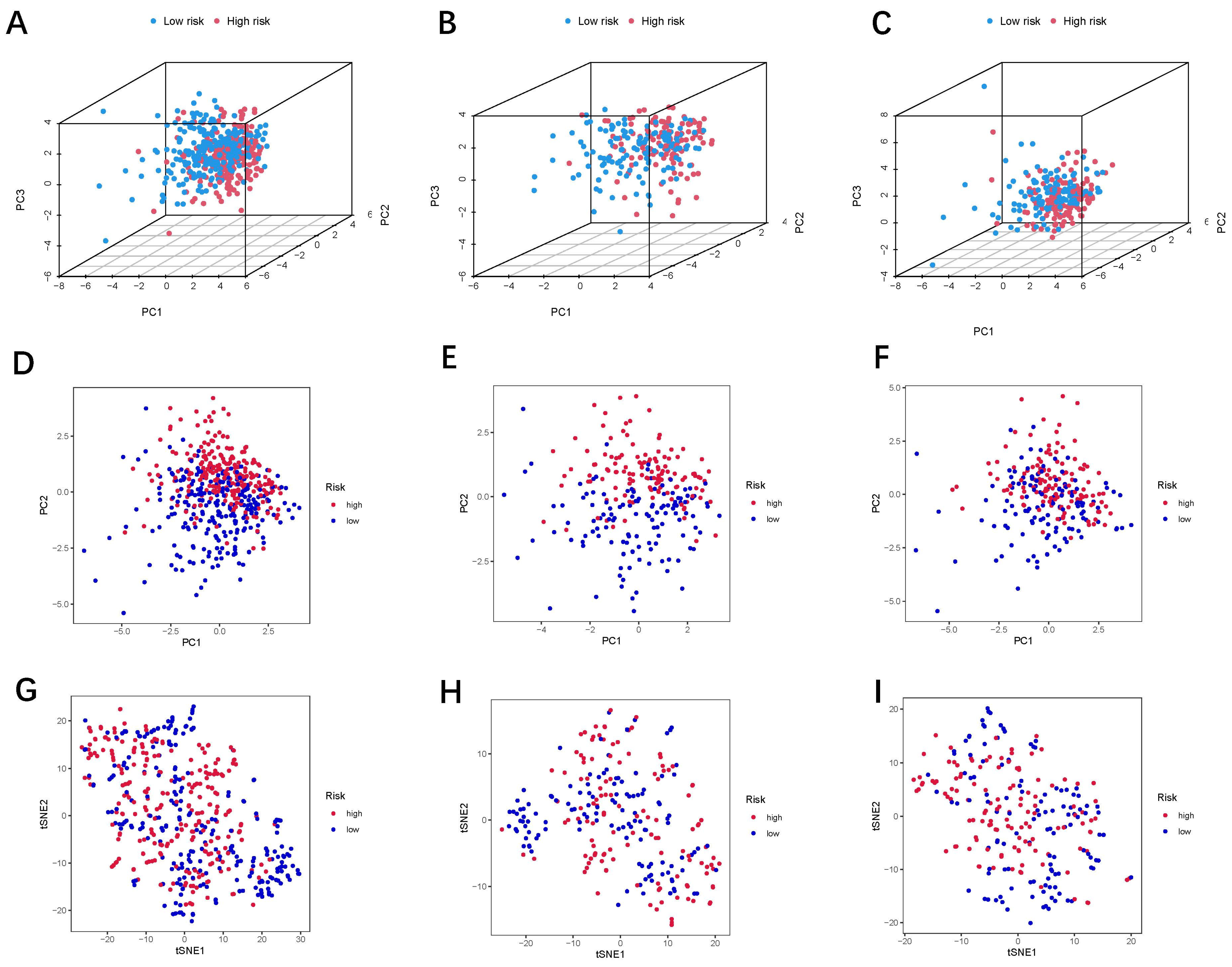
3.2. Establishment and Verification of the 24-LncRNAs Prognostic Risk Scores
3.3. Assessment of Nomogram
3.4. Immune Cell Infiltration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, H.; Zhou, L.; Shen, N.; Ning, X.; Wu, D.; Jiang, K.; Huang, X. M1 macrophage-derived exosomes and their key molecule lncRNA HOTTIP suppress head and neck squamous cell carcinoma progression by upregulating the TLR5/NF-kappaB pathway. Cell Death Dis. 2022, 13, 183. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, N.; Zhou, Q.; Li, Z.; Shen, L. Development and Validation of an Autophagy-Related LncRNA Prognostic Signature in Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 743611. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Song, H. Development of Gene Therapeutics for Head and Neck Cancer in China: From Bench to Bedside. Hum. Gene Ther. 2018, 29, 180–187. [Google Scholar] [CrossRef] [PubMed]
- St, L.G.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Maenner, S.; Muller, M.; Becker, P.B. Roles of long, non-coding RNA in chromosome-wide transcription regulation: Lessons from two dosage compensation systems. Biochimie 2012, 94, 1490–1498. [Google Scholar] [CrossRef]
- Wei, B.; Wei, W.; Zhao, B.; Guo, X.; Liu, S. Long non-coding RNA HOTAIR inhibits miR-17-5p to regulate osteogenic differentiation and proliferation in non-traumatic osteonecrosis of femoral head. PLoS ONE 2017, 12, e169097. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Guo, H.; Tong, T.; Xie, F.; Qin, X.; Wang, X.; Chen, W.; Zhang, J. lncRNA lnc-POP1-1 upregulated by VN1R5 promotes cisplatin resistance in head and neck squamous cell carcinoma through interaction with MCM5. Mol. Ther. 2022, 30, 448–467. [Google Scholar] [CrossRef]
- Jiang, Y.; Cao, W.; Wu, K.; Qin, X.; Wang, X.; Li, Y.; Yu, B.; Zhang, Z.; Wang, X.; Yan, M.; et al. LncRNA LINC00460 promotes EMT in head and neck squamous cell carcinoma by facilitating peroxiredoxin-1 into the nucleus. J. Exp. Clin. Cancer Res. 2019, 38, 365. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Xiao, H.; Yang, Q.; Hu, R.; Jiang, L.; Bi, R.; Jiang, X.; Wang, L.; Mei, J.; Ding, F.; et al. The USP21/YY1/SNHG16 axis contributes to tumor proliferation, migration, and invasion of non-small-cell lung cancer. Exp. Mol. Med. 2020, 52, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Xu, J.; Wang, W.; Zhao, H.; Liu, H.; Liu, X.; Liu, J.; Sun, Y.; Dunaif, A.; Du, Y.; et al. Long non-coding RNA LINC-01572:28 inhibits granulosa cell growth via a decrease in p27 (Kip1) degradation in patients with polycystic ovary syndrome. Ebiomedicine 2018, 36, 526–538. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Du, X.; Liu, M.; Wang, J.; Sun, L.; Li, Y. Increased expression of long non-coding RNA SNHG16 correlates with tumor progression and poor prognosis in non-small cell lung cancer. Int. J. Biol. Macromol. 2019, 121, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J. Oxidative stress, antioxidant defenses, and damage removal, repair, and replacement systems. IUBMB Life 2000, 50, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, T.; Dong, X.; Wang, X.; Zhang, Z.; Zhao, C.; Kang, X.; Zheng, R.; Li, X. Untargeted Metabolomic Profiling of Cuprizone-Induced Demyelination in Mouse Corpus Callosum by UPLC-Orbitrap/MS Reveals Potential Metabolic Biomarkers of CNS Demyelination Disorders. Oxidative Med. Cell. Longev. 2021, 2021, 7093844. [Google Scholar] [CrossRef]
- Zhao, Z.; Zheng, R.; Wang, X.; Li, T.; Dong, X.; Zhao, C.; Li, X. Integrating Lipidomics and Transcriptomics Reveals the Crosstalk Between Oxidative Stress and Neuroinflammation in Central Nervous System Demyelination. Front. Aging Neurosci. 2022, 14, 870957. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Zhong, H.; Yin, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biol. 2015, 4, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Sutaria, S.R.; Gori, S.S.; Morris, J.D.; Xie, Z.; Fu, X.A.; Nantz, M.H. Lipid Peroxidation Produces a Diverse Mixture of Saturated and Unsaturated Aldehydes in Exhaled Breath That Can Serve as Biomarkers of Lung Cancer-A Review. Metabolites 2022, 12, 561. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab, W.M. Evidence in support of potential applications of lipid peroxidation products in cancer treatment. Oxidative Med. Cell. Longev. 2013, 2013, 931251. [Google Scholar] [CrossRef]
- Gupta, A.; Bhatt, M.L.; Misra, M.K. Lipid peroxidation and antioxidant status in head and neck squamous cell carcinoma patients. Oxidative Med. Cell. Longev. 2009, 2, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Si, Y.; Feng, Y.; Zhuo, S.; Cui, B.; Zhang, Z. Prognostic value of lipid metabolism-related genes in head and neck squamous cell carcinoma. Immun. Inflamm. Dis. 2021, 9, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Takashi, Y.; Ouchi, Y.; Kuwahara, Y.; Igarashi, K.; Nagasawa, T.; Nabika, H.; Kurimasa, A.; Fukumoto, M.; Nishitani, Y.; et al. Lipid peroxidation increases hydrogen peroxide permeability leading to cell death in cancer cell lines that lack mtDNA. Cancer Sci. 2019, 110, 2856–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaganjac, M.; Borovic, S.S.; Zarkovic, N. Utilizing Iron for Targeted Lipid Peroxidation as Anticancer Option of Integrative Biomedicine: A Short Review of Nanosystems Containing Iron. Antioxidants 2020, 9, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Yang, Y.; Wang, F.; Yang, W.G.; Zhang, J.J.; Zou, Z.Q. Docosahexaenoic acid monoglyceride induces apoptosis and autophagy in breast cancer cells via lipid peroxidation-mediated endoplasmic reticulum stress. J. Food Sci. 2021, 86, 4704–4716. [Google Scholar] [CrossRef]
- Ulker, O.C.; Panieri, E.; Suzen, S.; Jaganjac, M.; Zarkovic, N.; Saso, L. Short overview on the relevance of microRNA-reactive oxygen species (ROS) interactions and lipid peroxidation for modulation of oxidative stress-mediated signalling pathways in cancer treatment. J. Pharm. Pharmacol. 2022, 74, 503–515. [Google Scholar] [CrossRef]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Lorch, J.H.; Goloubeva, O.; Haddad, R.I.; Cullen, K.; Sarlis, N.; Tishler, R.; Tan, M.; Fasciano, J.; Sammartino, D.E.; Posner, M.R. Induction chemotherapy with cisplatin and fluorouracil alone or in combination with docetaxel in locally advanced squamous-cell cancer of the head and neck: Long-term results of the TAX 324 randomised phase 3 trial. Lancet Oncol. 2011, 12, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Marur, S.; Forastiere, A.A. Head and Neck Squamous Cell Carcinoma: Update on Epidemiology, Diagnosis, and Treatment. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2016; Volume 91, pp. 386–396. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Li, Y.; Yang, X.; Wu, X.; Liu, Z.; Mou, Y.; Song, X. Identification and Validation of 17-lncRNA Related to Regulatory T Cell Heterogeneity as a Prognostic Signature for Head and Neck Squamous Cell Carcinoma. Front. Immunol. 2021, 12, 782216. [Google Scholar] [CrossRef]
- Wang, X.; Cao, K.; Guo, E.; Mao, X.; Guo, L.; Zhang, C.; Guo, J.; Wang, G.; Yang, X.; Sun, J.; et al. Identification of Immune-Related LncRNA Pairs for Predicting Prognosis and Immunotherapeutic Response in Head and Neck Squamous Cell Carcinoma. Front. Immunol. 2021, 12, 658631. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Quan, Z.; Hao, Y.; Liu, J.; Wang, Z.; Dai, L.; Dai, H.; He, S.; Tang, B. Long Non-Coding RNA LINC01572 Promotes Hepatocellular Carcinoma Progression via Sponging miR-195-5p to Enhance PFKFB4-Mediated Glycolysis and PI3K/AKT Activation. Front. Cell Dev. Biol. 2021, 9, 783088. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Jia, N.; Li, W.; Zhang, X.Y. LINC01572 Regulates Cisplatin Resistance in Gastric Cancer Cells by Mediating miR-497-5p. Onco Targets Ther. 2020, 13, 10877–10887. [Google Scholar] [CrossRef]
- Liu, J.Q.; Feng, Y.H.; Zeng, S.; Zhong, M.Z. linc01088 promotes cell proliferation by scaffolding EZH2 and repressing p21 in human non-small cell lung cancer. Life Sci. 2020, 241, 117134. [Google Scholar] [CrossRef]
- Li, C.; Pan, B.; Wang, X.; Liu, X.; Qin, J.; Gao, T.; Sun, H.; Pan, Y.; Wang, S. Upregulated LINC01088 facilitates malignant phenotypes and immune escape of colorectal cancer by regulating microRNAs/G3BP1/PD-L1 axis. J. Cancer Res. Clin. Oncol. 2022, 148, 1965–1982. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Chen, D.L.; Chen, S.L. LINC01088 promotes the growth and invasion of glioma cells through regulating small nuclear ribonucleoprotein polypeptide A transcription. Bioengineered 2022, 13, 9172–9183. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Gu, H.; Chen, Y.; Shi, Y.; Liu, C.; Lv, L.; Sun, Y.; Zhang, Y. Long non-coding RNA CASC8 polymorphisms are associated with the risk of esophageal cancer in a Chinese population. Thorac. Cancer 2020, 11, 2852–2857. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, W.; Liu, K.; Liu, Y. LncRNA SNHG16 promotes tumor growth of pancreatic cancer by targeting miR-218-5p. Biomed. Pharmacother. 2019, 114, 108862. [Google Scholar] [CrossRef]
- Christensen, L.L.; True, K.; Hamilton, M.P.; Nielsen, M.M.; Damas, N.D.; Damgaard, C.K.; Ongen, H.; Dermitzakis, E.; Bramsen, J.B.; Pedersen, J.S.; et al. SNHG16 is regulated by the Wnt pathway in colorectal cancer and affects genes involved in lipid metabolism. Mol. Oncol. 2016, 10, 1266–1282. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Z.; Li, R.; Wang, S.; Wang, W.; Tang, M.; Zhang, W. The role of ANXA5 in DBP-induced oxidative stress through ERK/Nrf2 pathway. Environ. Toxicol. Pharmacol. 2019, 72, 103236. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, S.; Zhang, D.; Tao, Q.; Zhang, T.; Liu, G.; Liu, X.; Zhao, T. Identification of SERPINE1, PLAU and ACTA1 as biomarkers of head and neck squamous cell carcinoma based on integrated bioinformatics analysis. Int. J. Clin. Oncol. 2019, 24, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Chen, L.; Yang, Y.C.; Wang, X.M.; Wang, R.Y.; Li, L.; Wen, W.; Chang, Y.X.; Chen, C.Y.; Tang, J. CYP3A5 Functions as a Tumor Suppressor in Hepatocellular Carcinoma by Regulating mTORC2/Akt Signaling. Cancer Res. 2015, 75, 1470–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Shults, K.; Flye, L.; Jiang, F.; Head, D.R.; Briggs, R.C. Overexpression of GSTA2 protects against cell cycle arrest and apoptosis induced by the DNA inter-strand crosslinking nitrogen mustard, mechlorethamine. J. Cell. Biochem. 2005, 95, 339–351. [Google Scholar] [CrossRef]
- Stalnecker, C.A.; Grover, K.R.; Edwards, A.C.; Coleman, M.F.; Yang, R.; DeLiberty, J.M.; Papke, B.; Goodwin, C.M.; Pierobon, M.; Petricoin, E.F.; et al. Concurrent Inhibition of IGF1R and ERK Increases Pancreatic Cancer Sensitivity to Autophagy Inhibitors. Cancer Res. 2022, 82, 586–598. [Google Scholar] [CrossRef]
- Williams, N.C.; O’Neill, L.A. ACLY-matizing Macrophages to Histone Modification during Immunometabolic Reprogramming. Trends Immunol. 2020, 41, 93–94. [Google Scholar] [CrossRef]
- Kuo, C.; Chou, H.; Chiu, Y.; Cheng, A.; Fan, C.; Chang, Y.; Chen, C.; Jiang, S.; Chen, N.; Lee, A.Y. Mitochondrial oxidative stress by Lon-PYCR1 maintains an immunosuppressive tumor microenvironment that promotes cancer progression and metastasis. Cancer Lett. 2020, 474, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Zhang, L.; Zhu, Z.; Li, R.; Wang, S.; Wang, W.; Qin, Z.; Zhang, W. Overexpression of miR-506-3p Aggravates DBP-Induced Testicular Oxidative Stress in Rats by Downregulating ANXA5 via Nrf2/HO-1 Signaling Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 4640605. [Google Scholar] [CrossRef]
- Peng, B.; Liu, S.; Guo, C.; Sun, X.; Sun, M.Z. ANXA5 level is linked to in vitro and in vivo tumor malignancy and lymphatic metastasis of murine hepatocarcinoma cell. Future Oncol. 2016, 12, 31–42. [Google Scholar] [CrossRef]
- Sun, X.; Wei, B.; Liu, S.; Guo, C.; Wu, N.; Liu, Q.; Sun, M.Z. Anxa5 mediates the in vitro malignant behaviours of murine hepatocarcinoma Hca-F cells with high lymph node metastasis potential preferentially via ERK2/p-ERK2/c-Jun/p-c-Jun(Ser73) and E-cadherin. Biomed. Pharmacother. 2016, 84, 645–654. [Google Scholar] [CrossRef]
- Arroyo-Solera, I.; Pavon, M.A.; Leon, X.; Lopez, M.; Gallardo, A.; Cespedes, M.V.; Casanova, I.; Pallares, V.; Lopez-Pousa, A.; Mangues, M.A.; et al. Effect of serpinE1 overexpression on the primary tumor and lymph node, and lung metastases in head and neck squamous cell carcinoma. Head Neck 2019, 41, 429–439. [Google Scholar] [CrossRef]
- Minemura, C.; Asai, S.; Koma, A.; Kase-Kato, I.; Tanaka, N.; Kikkawa, N.; Kasamatsu, A.; Yokoe, H.; Hanazawa, T.; Uzawa, K.; et al. Identification of Tumor-Suppressive miR-30e-3p Targets: Involvement of SERPINE1 in the Molecular Pathogenesis of Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 3808. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Goodman, O.J. CYP3A5 regulates prostate cancer cell growth by facilitating nuclear translocation of AR. Prostate 2015, 75, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Jameson, M.J. Downregulation of IGF1R Expression Inhibits Growth and Enhances Cisplatin Sensitivity of Head and Neck Squamous Cell Carcinoma Cells In Vitro. Horm. Cancer 2019, 10, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Granchi, C. ATP citrate lyase (ACLY) inhibitors: An anti-cancer strategy at the crossroads of glucose and lipid metabolism. Eur. J. Med. Chem. 2018, 157, 1276–1291. [Google Scholar] [CrossRef] [PubMed]
- Gottgens, E.L.; van den Heuvel, C.N.; de Jong, M.C.; Kaanders, J.H.; Leenders, W.P.; Ansems, M.; Bussink, J.; Span, P.N. ACLY (ATP Citrate Lyase) Mediates Radioresistance in Head and Neck Squamous Cell Carcinomas and is a Novel Predictive Radiotherapy Biomarker. Cancers 2019, 11, 1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Kim, B.S.; Han, W.; Chen, X.; Yan, Y.; Lin, L.; Chai, G. Identifying Oxidized Lipid Metabolism-Related LncRNAs as Prognostic Biomarkers of Head and Neck Squamous Cell Carcinoma. J. Pers. Med. 2023, 13, 488. https://doi.org/10.3390/jpm13030488
Zhang Z, Kim BS, Han W, Chen X, Yan Y, Lin L, Chai G. Identifying Oxidized Lipid Metabolism-Related LncRNAs as Prognostic Biomarkers of Head and Neck Squamous Cell Carcinoma. Journal of Personalized Medicine. 2023; 13(3):488. https://doi.org/10.3390/jpm13030488
Chicago/Turabian StyleZhang, Ziwei, Byeong Seop Kim, Wenqing Han, Xiaojun Chen, Yingjie Yan, Li Lin, and Gang Chai. 2023. "Identifying Oxidized Lipid Metabolism-Related LncRNAs as Prognostic Biomarkers of Head and Neck Squamous Cell Carcinoma" Journal of Personalized Medicine 13, no. 3: 488. https://doi.org/10.3390/jpm13030488