
STAT3 and NTRK2 Genes Predicted by the Bioinformatics Approach May Play Important Roles in the Pathogenesis of Multiple Sclerosis and Obsessive–Compulsive Disorder

 , ,
, ,
Abstract
:1. Introduction
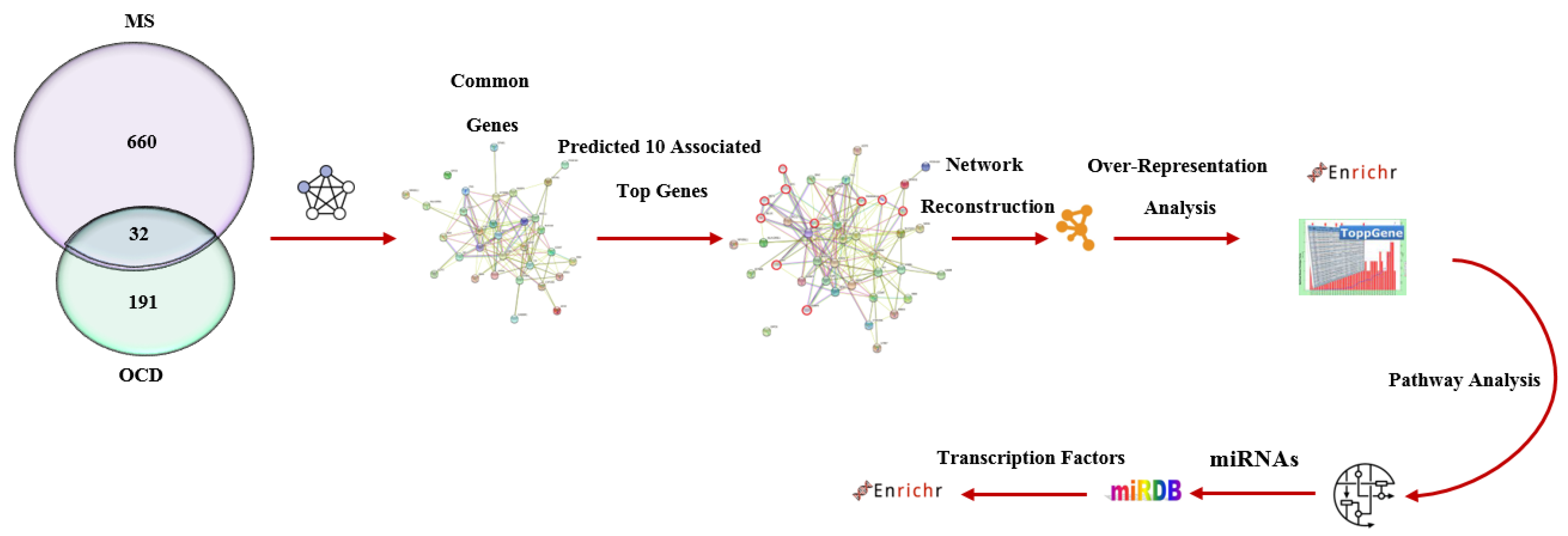
2. Materials and Methods
2.1. Study Design
2.2. Gene Set Selection
2.3. Genetic Network Reconstruction Using Cytoscape
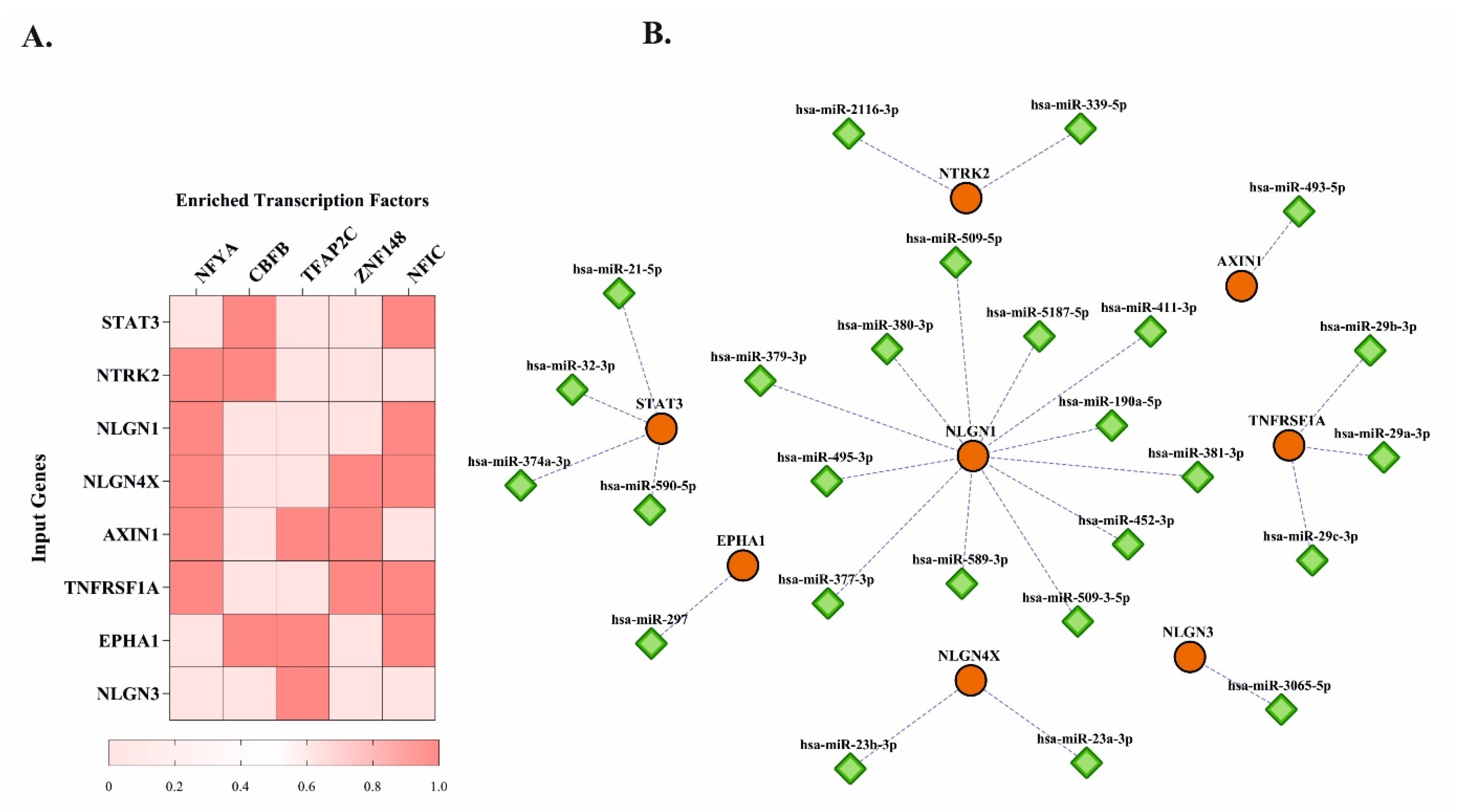
2.4. TRANSFAC Analysis and microRNA Target Prediction
2.5. Gene Ontology Enrichment Analysis
3. Results
3.1. Finding Genes According to the Literature Review and Harmonizome
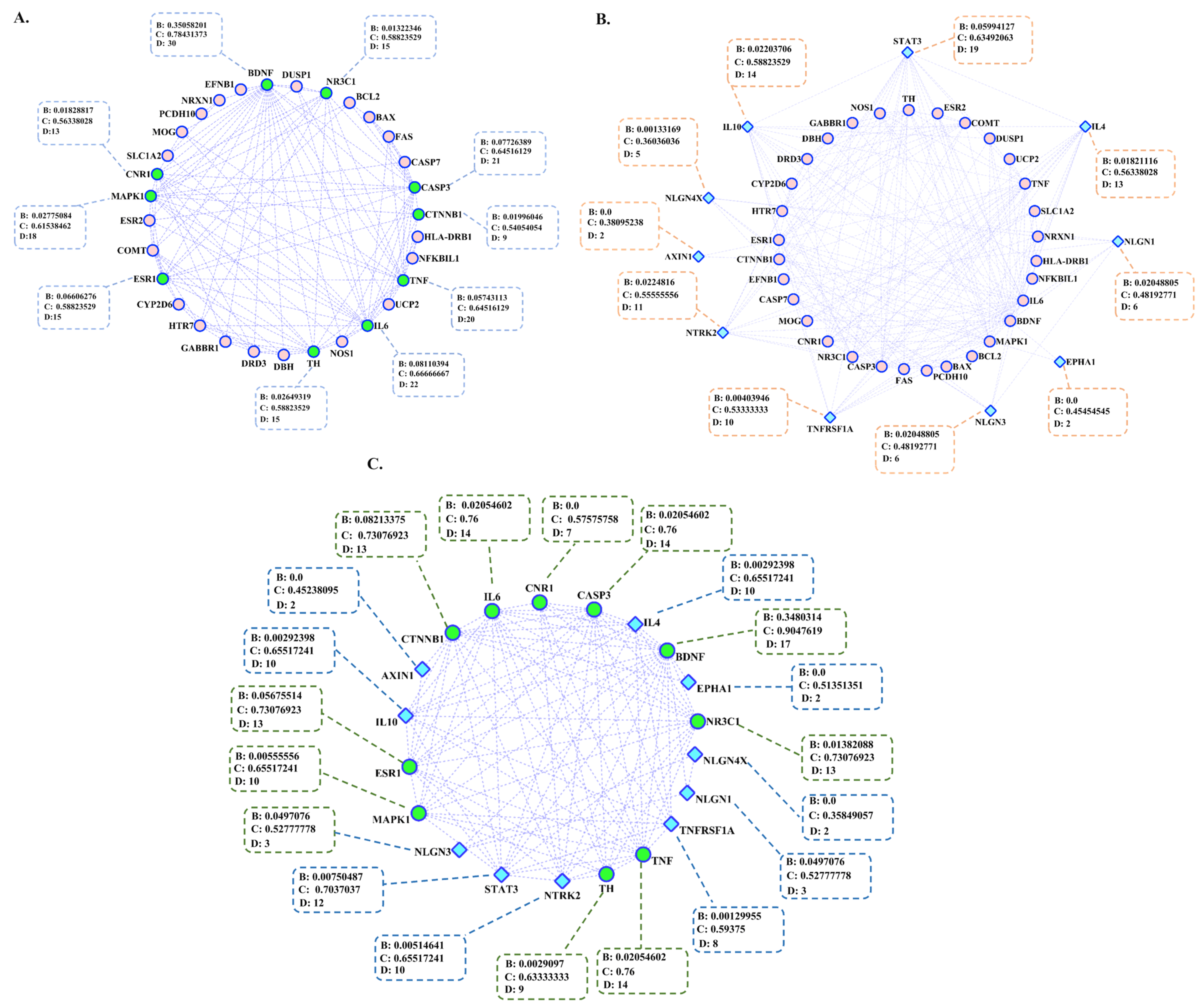
3.2. Genetic Network Reconstruction
3.3. Predicted Transcription Factors and miRNAs for Hub Genes
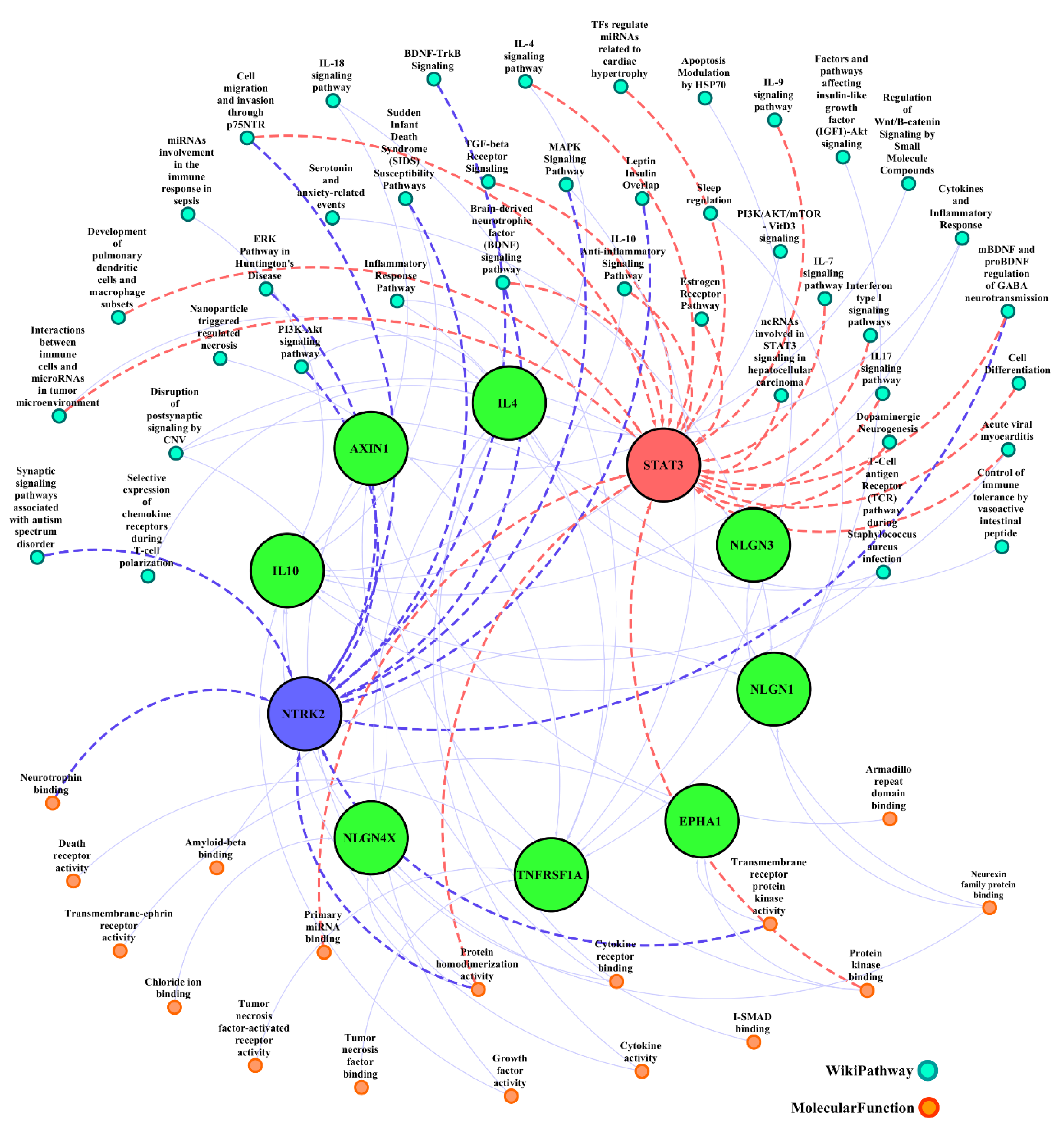
3.4. Gene Ontology Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Engin, B.; Uguz, F.; Yilmaz, E.; Özdemir, M.; Mevlitoglu, I. The levels of depression, anxiety and quality of life in patients with chronic idiopathic urticaria. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 36–40. [Google Scholar] [CrossRef]
- Filser, M.; Baetge, S.J.; Balloff, C.; Buchner, A.; Fink, G.R.; Heibel, M.; Meier, U.; Rau, D.; Renner, A.; Schreiber, H.; et al. Mental symptoms in MS (MeSyMS): Development and validation of a new assessment. Mult. Scler. Relat. Disord. 2021, 49, 102744. [Google Scholar] [CrossRef] [PubMed]
- Foroughipour, M.; Behdani, F.; Hebrani, P.; Marvast, M.N.; Esmatinia, F.; Akhavanrezayat, A. Frequency of obsessive-compulsive disorder in patients with multiple sclerosis: A cross-sectional study. J. Res. Med. Sci. 2012, 17, 248–253. [Google Scholar] [PubMed]
- Mrabet, S.; Boudriga, S.; Souissi, A.; Gharbi, A.; Nasri, A.; Ben Djebara, M.; Gargouri, A.; Kacem, I.; Gouider, R. Depression, Anxiety and Alexithymia in Tunisian Patients with Multiple Sclerosis. Mult. Scler. Relat. Disord. 2022, 59, 103596. [Google Scholar] [CrossRef]
- Khatri, I.A.; Aljwair, S.; Alammar, H.; Altariq, A.; Masud, N.; Al Malik, Y.; Kojan, S. Social Anxiety and Obsessive-Compulsive Disorder Are Common Among Persons With Multiple Sclerosis at King Abdulaziz Medical City, Riyadh. Cureus 2021, 13, e13619. [Google Scholar] [CrossRef]
- Soureshgani, A.; Khodabakhshi-Koolaee, A. Analysis of Obsessive-Compulsive disorder experiences in multiple sclerosis patients: A phenomenological study. Iran. J. Psychiatr. Nurs. 2021, 9, 1–11. [Google Scholar]
- Ostler, T.; Ackerson, B. Chronic Parental Mental Illness. In Encyclopedia of Infant and Early Childhood Development, 2nd ed.; Benson, J.B., Ed.; Elsevier: Oxford, UK, 2020; pp. 306–316. [Google Scholar]
- Douzenis, A.; Michalopoulou, P.G.; Voumvourakis, C.; Typaldou, M.; Michopoulos, I.; Lykouras, L. Obsessive-compulsive disorder associated with parietal white matter multiple sclerosis plaques. World J. Biol. Psychiatry 2009, 10, 956–960. [Google Scholar] [CrossRef]
- Tinelli, E.; Francia, A.; Quartuccio, E.; Morreale, M.; Contessa, G.; Pascucci, S.; Sbardella, E.; Pozzilli, C.; Pantano, P. Structural brain MR imaging changes associated with obsessive-compulsive disorder in patients with multiple sclerosis. Am. J. Neuroradiol. 2013, 34, 305–309. [Google Scholar] [CrossRef] [Green Version]
- George, M.S.; Kellner, C.H.; Fossey, M.D. Obsessive-compulsive symptoms in a patient with multiple sclerosis. J. Nerv. Ment. Dis. 1989, 177, 304–305. [Google Scholar] [CrossRef]
- Korostil, M.; Feinstein, A. Anxiety disorders and their clinical correlates in multiple sclerosis patients. Mult. Scler. 2007, 13, 67–72. [Google Scholar] [CrossRef]
- Kinnear, C.J.; Niehaus, D.J.H.; Moolman-Smook, J.C.; du Toit, P.L.; van Kradenberg, J.; Weyers, J.B.; Potgieter, A.; Marais, V.; Emsley, R.A.; Knowles, J.A.; et al. Obsessive-compulsive disorder and the promoter region polymorphism (5-HTTLPR) in the serotonin transporter gene (SLC6A4): A negative association study in the Afrikaner population. Int. J. Neuropsychopharmacol. 2000, 3, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme, R.; Krebs, M.-O.; Chabane, N.; Roy, I.; Millet, B.; Mouren-Simeoni, M.C.; Maier, W.; Bourgeron, T.; Leboyer, M. Frequency and transmission of glutamate receptors GRIK2 and GRIK3 polymorphisms in patients with obsessive compulsive disorder. Neuroreport 2004, 15, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Camarena, B.; Rinetti, G.; Cruz, C.; Gómez, A.; de la Fuente, J.R.; Nicolini, H. Additional evidence that genetic variation of MAO-A gene supports a gender subtype in obsessive-compulsive disorder. Am. J. Med. Genet. 2001, 105, 279–282. [Google Scholar] [CrossRef]
- Cruz, C.; Camarena, B.; King, N.; Páez, F.; Sidenberg, D.; de la Fuente, J.R.; Nicolini, H. Increased prevalence of the seven-repeat variant of the dopamine D4 receptor gene in patients with obsessive-compulsive disorder with tics. Neurosci. Lett. 1997, 231, 1–4. [Google Scholar] [CrossRef]
- Karayiorgou, M.; Sobin, C.; Blundell, M.L.; Galke, B.L.; Malinova, L.; Goldberg, P.; Ott, J.; Gogos, J.A. Family-based association studies support a sexually dimorphic effect of COMT and MAOA on genetic susceptibility to obsessive-compulsive disorder. Biol. Psychiatry 1999, 45, 1178–1189. [Google Scholar] [CrossRef]
- Hall, D.; Dhilla, A.; Charalambous, A.; Gogos, J.A.; Karayiorgou, M. Sequence variants of the brain-derived neurotrophic factor (BDNF) gene are strongly associated with obsessive-compulsive disorder. Am. J. Hum. Genet. 2003, 73, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Jensen, L.J. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Von Mering, C. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2007, 2, 2366. [Google Scholar] [CrossRef] [Green Version]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2014, 43, D146–D152. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gene Ontology Consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [CrossRef] [PubMed]
- Nayak, R.C.; Chang, K.H.; Singh, A.K.; Kotliar, M.; Desai, M.; Wellendorf, A.M.; Wunderlich, M.; Bartram, J.; Mizukawa, B.; Cuadrado, M.; et al. Nuclear Vav3 is required for polycomb repression complex-1 activity in B-cell lymphoblastic leukemogenesis. Nat. Commun. 2022, 13, 1–16. [Google Scholar] [CrossRef]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef]
- Endres, D.; Pollak, T.A.; Bechter, K.; Denzel, D.; Pitsch, K.; Nickel, K.; Runge, K.; Pankratz, B.; Klatzmann, D.; Tamouza, R.; et al. Immunological causes of obsessive-compulsive disorder: Is it time for the concept of an “autoimmune OCD” subtype? Transl. Psychiatry 2022, 12, 5. [Google Scholar] [CrossRef]
- Pattwell, S.S.; Konnick, E.Q.; Liu, Y.J.; Yoda, R.A.; Sekhar, L.N.; Cimino, P.J. Neurotrophic Receptor Tyrosine Kinase 2 (NTRK2) Alterations in Low-Grade Gliomas: Report of a Novel Gene Fusion Partner in a Pilocytic Astrocytoma and Review of the Literature. Case Rep. Pathol. 2020, 2020, 5903863. [Google Scholar] [CrossRef] [Green Version]
- Alonso, P.; Gratacòs, M.; Menchón, J.M.; Saiz-Ruiz, J.; Segalàs, C.; Baca-García, E.; Labad, J.; Fernández-Piqueras, J.; Real, E.; Vaquero, C.; et al. Extensive genotyping of the BDNF and NTRK2 genes define protective haplotypes against obsessive-compulsive disorder. Biol. Psychiatry 2008, 63, 619–628. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, G.; Di, Z.; Zhao, Q. miR-339-5p inhibits alcohol-induced brain inflammation through regulating NF-κB pathway. Biochem. Biophys. Res. Commun. 2014, 452, 450–456. [Google Scholar] [CrossRef]
- Lu, H.C.; Kim, S.; Steelman, A.J.; Tracy, K.; Zhou, B.; Michaud, D.; Hillhouse, A.E.; Konganti, K.; Li, J. STAT3 signaling in myeloid cells promotes pathogenic myelin-specific T cell differentiation and autoimmune demyelination. Proc. Natl. Acad. Sci. USA 2020, 117, 5430–5441. [Google Scholar] [CrossRef] [Green Version]
- Reisinger, S. Transcriptional and Behavioural Modulation by Serotonergic STAT3 Relevant to Mood and Psychotic Disorders; Department of Neurophysiology and Neuropharmacology Center for Physiology: Tokyo, Japan, 2020. [Google Scholar]
- de Oliveira, K.C.; Camilo, C.; Gastaldi, V.D.; Sant’Anna Feltrin, A.; Lisboa BC, G.; de Jesus Rodrigues de Paula, V.; Brentani, H. Brain areas involved with obsessive-compulsive disorder present different DNA methylation modulation. BMC Genomic Data 2021, 22, 45. [Google Scholar] [CrossRef] [PubMed]
- Cosco, T.D.; Pillinger, T.; Emam, H.; Solmi, M.; Budhdeo, S.; Prina, A.M.; Maes, M.; Stein, D.J.; Stubbs, B.; Carvalho, A.F. Immune aberrations in obsessive-compulsive disorder: A systematic review and meta-analysis. Mol. Neurobiol. 2019, 56, 4751–4759. [Google Scholar] [CrossRef] [PubMed]
- Al-Naseri, M.A.; Salman, E.D.; Ad’hiah, A.H. Association between interleukin-4 and interleukin-10 single nucleotide polymorphisms and multiple sclerosis among Iraqi patients. Neurol. Sci. 2019, 40, 2383–2389. [Google Scholar] [CrossRef] [PubMed]
- Zhang , J.; Luo, Y. Degree centrality, betweenness centrality, and closeness centrality in social network. In Proceedings of the 2017 2nd International Conference on Modelling, Simulation and Applied Mathematics (MSAM2017), Bangkok, Thailand, 26–27 March 2017; Atlantis Press: Amsterdam, The Netherlands. [Google Scholar]
- Golbeck, J. (Ed.) Chapter 3—Network Structure and Measures. In Analyzing the Social Web; Morgan Kaufmann: Boston, MA, USA, 2013; pp. 25–44. [Google Scholar]
- Zito, A.; Lualdi, M.; Granata, P.; Cocciadiferro, D.; Novelli, A.; Alberio, T.; Casalone, R.; Fasano, M. Gene Set Enrichment Analysis of Interaction Networks Weighted by Node Centrality. Front. Genet. 2012, 12, 577623. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Genes Associated with Diseases | ||
|---|---|---|
| Gene Source | Multiple Sclerosis | Obsessive–Compulsive Disorder |
| Literature review | 368 | 133 |
| Harmonizome | 292 | 58 |
| Total genes | 660 | 191 |
| Shared genes | 32 | |
| Hub genes (Acquired by STRING) | 10 | |
| Description | Target Genes | −log (p-Value) |
|---|---|---|
| Presynaptic membrane assembly | NLGN3, NLGN1, NLGN4X | 7.871923987 |
| Postsynaptic membrane assembly | NLGN3, NLGN1, NLGN4X | 7.747146969 |
| Presynaptic membrane organization | NLGN3, NLGN1, NLGN4X | 7.528708289 |
| Regulation of chronic inflammatory response | IL10, TNFRSF1A, IL4 | 7.178551981 |
| Neuron cell–cell adhesion | NLGN3, NLGN1, NLGN4X | 7.178551981 |
| Positive regulation of developmental process | NLGN3, NLGN1, NTRK2, STAT3, IL4, IL10, EPHA1, TNFRSF1A | 7.108016769 |
| Regulation of multicellular organismal development | NLGN3, NLGN1, NTRK2, STAT3, IL4, IL10, EPHA1, TNFRSF1A | 6.813326133 |
| Positive regulation of signal transduction | NLGN3, NLGN1, NTRK2, STAT3, IL4, IL10, AXIN1, TNFRSF1A | 6.771086594 |
| Positive regulation of multicellular organismal process | NLGN3, NLGN1, NTRK2, STAT3, IL4, IL10, EPHA1, TNFRSF1A | 6.753009301 |
| Regulation of anatomical structure morphogenesis | NLGN3, NLGN1, NTRK2, STAT3, IL10, EPHA1, TNFRSF1A | 6.378512135 |
| Positive regulation of synaptic transmission, glutamatergic | NLGN3, NLGN1, NTRK2 | 6.16627931 |
| Postsynapse assembly | NLGN3, NLGN1, NLGN4X | 6.16627931 |
| Chronic inflammatory response | IL10, TNFRSF1A, IL4 | 6.1316495 |
| Peptidyl-tyrosine phosphorylation | NTRK2, STAT3, IL4, EPHA1, TNFRSF1A | 6.070274622 |
| Peptidyl-tyrosine modification | NTRK2, STAT3, IL4, EPHA1, TNFRSF1A | 6.05527064 |
| Cell junction organization | NLGN3, NLGN1, NLGN4X, IL10, EPHA1, NTRK2 | 5.919734373 |
| Behavior | NLGN3, NLGN1, NTRK2, STAT3, AXIN1, NLGN4X | 5.806041022 |
| Postsynaptic membrane organization | NLGN3, NLGN1, NLGN4X | 5.775208044 |
| Receptor signaling pathway via JAK-STAT | IL10, TNFRSF1A, IL4, STAT3 | 5.767766479 |
| Regulation of nervous system process | NLGN3, NLGN1, NLGN4X, IL10 | 5.72514968 |
| Receptor signaling pathway via STAT | IL10, TNFRSF1A, IL4, STAT3 | 5.699839463 |
| Cell junction assembly | NLGN3, NLGN1, NLGN4X, NTRK2, EPHA1 | 5.661145254 |
| Regulation of tumor necrosis factor production | IL10, TNFRSF1A, IL4, STAT3 | 5.634699251 |
| Negative regulation of reactive oxygen species biosynthetic process | IL10, IL4, STAT3 | 5.628193541 |
| Blood vessel morphogenesis | IL10, TNFRSF1A, EPHA1, STAT3, NTRK2, NLGN1 | 5.626168855 |
| Regulation of tumor necrosis factor superfamily cytokine production | IL10, TNFRSF1A, IL4, STAT3 | 5.595166283 |
| Tumor necrosis factor production | IL10, TNFRSF1A, IL4, STAT3 | 5.587371479 |
| Modulation of excitatory postsynaptic potential | NLGN3, NLGN1, NLGN4X | 5.56050941 |
| Presynapse assembly | NLGN3, NLGN1, NLGN4X | 5.56050941 |
| Positive regulation of angiogenesis | IL10, TNFRSF1A, EPHA1, STAT3 | 5.548981548 |
| Positive regulation of vasculature development | IL10, TNFRSF1A, EPHA1, STAT3 | 5.548981548 |
| Tumor necrosis factor superfamily cytokine production | IL10, TNFRSF1A, IL4, STAT3 | 5.541362151 |
| Negative regulation of chronic inflammatory response | IL10, IL4 | 5.500725418 |
| Positive regulation of peptidyl-tyrosine phosphorylation | IL4, TNFRSF1A, STAT3, NTRK2 | 5.496481687 |
| Regulation of cell junction assembly | NLGN3, NLGN1, NTRK2, EPHA1 | 5.481881053 |
| Synapse assembly | NLGN3, NLGN1, NLGN4X, NTRK2 | 5.481881053 |
| Synapse organization | NLGN3, NLGN1, NLGN4X, NTRK2, IL10 | 5.430977414 |
| Positive regulation of phosphorylation | IL4, NTRK2, STAT3, AXIN1, EPHA1, TNFRSF1A | 5.377785977 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepehrinezhad, A.; Shahbazi, A.; Bozorgmehr, A.; Kateb, B.; Yamamoto, V.; Negah, S.S. STAT3 and NTRK2 Genes Predicted by the Bioinformatics Approach May Play Important Roles in the Pathogenesis of Multiple Sclerosis and Obsessive–Compulsive Disorder. J. Pers. Med. 2022, 12, 1043. https://doi.org/10.3390/jpm12071043
Sepehrinezhad A, Shahbazi A, Bozorgmehr A, Kateb B, Yamamoto V, Negah SS. STAT3 and NTRK2 Genes Predicted by the Bioinformatics Approach May Play Important Roles in the Pathogenesis of Multiple Sclerosis and Obsessive–Compulsive Disorder. Journal of Personalized Medicine. 2022; 12(7):1043. https://doi.org/10.3390/jpm12071043
Chicago/Turabian StyleSepehrinezhad, Ali, Ali Shahbazi, Ali Bozorgmehr, Babak Kateb, Vicky Yamamoto, and Sajad Sahab Negah. 2022. "STAT3 and NTRK2 Genes Predicted by the Bioinformatics Approach May Play Important Roles in the Pathogenesis of Multiple Sclerosis and Obsessive–Compulsive Disorder" Journal of Personalized Medicine 12, no. 7: 1043. https://doi.org/10.3390/jpm12071043